
Ahmil Sholeh
Graduate School of Biotechnology, University of Jember, Jl. Kalimantan 37 Kampus Tegalboto, Jember, East Java 68121, Indonesia
Bambang Sugiharto
Graduate School of Biotechnology, University of Jember, Jl. Kalimantan 37 Kampus Tegalboto, Jember, East Java 68121, Indonesia
LiveDNA: 62.27669
Hardian Susilo Addy
Graduate School of Biotechnology, University of Jember, Jl. Kalimantan 37 Kampus Tegalboto, Jember, East Java 68121, Indonesia
LiveDNA: 62.26922
Journal of Applied Sciences
Year: 2019 | Volume: 19 | Issue: 7 | Page No.: 647-653







ABSTRACT
Background and Objective: Sugarcane mosaic virus (SCMV) is a member of the genus Potyvirus (family of Potyviridae) causing mosaic worldwide distributed. Recently, local varieties including a variety of NXI-4T (a genetically modified sugarcane) were cultivating in East Java, which has never been monitored for SCMV occurrence. This research aimed to monitor the SCMV distribution among local varieties recently cultivated in East Java. Materials and Methods: The research was performed in the 4 Sugarcane Plantation under the authority of the company of Nusantara XI in East Java between October-November, 2017. The SCMV monitoring was done by estimating the disease incidence on 10% of plants of each location. Confirmation of the virus was done by RT-PCR and western blot. Chlorophyll a, chlorophyll b and total chlorophyll were estimated by colorimetric assay at dual wavelengths of 645 and 663 nm. Results: The data showed that the incidence of mosaic disease on all varieties observed was below 20% and categorized as resistant to mosaic disease while NXI-4T (genetically modified sugarcane) had a higher incidence of a mosaic than other varieties. The additional analysis confirmed and proved that mosaic disease on all local varieties was caused by SCMV according to RT-PCR and western blot assay. The decrease of chlorophyll was the consequence of SCMV infection. Conclusion: The SCMV was more frequently occurred on genetically modified sugarcane NXI 4T than others of non-genetically modified sugarcane.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ahmil Sholeh, Bambang Sugiharto and Hardian Susilo Addy, 2019. Monitoring Sugarcane mosaic virus (SCMV) on Recent Sugarcane Varieties in East Java, Indonesia. Journal of Applied Sciences, 19: 647-653.
DOI: 10.3923/jas.2019.647.653
URL: https://scialert.net/abstract/?doi=jas.2019.647.653
DOI: 10.3923/jas.2019.647.653
URL: https://scialert.net/abstract/?doi=jas.2019.647.653
INTRODUCTION
Sugarcane mosaic virus (SCMV) is most widely distributed and the most common sugarcane pathogen in the world, causing mosaic disease1. The SCMV has been reported to infect sugarcane in Indonesia and distributed in 59 plantations from 5 sugar factories in Central and East Java2. Recent data showed that SCMV was detected to infect several sugarcane varieties in East Java, such as; NXI-1T, VMC 7616, COKRO, PS 881 and PS 864 with the incidence of disease and severity of 26-78%, respectively3. However, there is no data available on the varieties that are recently planted in East Java, such as; local varieties of Bululawang, HW Merah, NXI 1-3 and one of the approved genetically modified (GM) sugarcane4, NXI-4T.
More early infection of the virus causes severe effects as compare to the late infection5. The initial infection usually results in chlorotic mosaic, yellows line on the green tissues of infected plants as well as leaf reddening and necrosis. The chlorosis occurs due to a significant decrease of total chlorophyll content3, as a consequence of the damage of the chloroplast which affected the photosynthesis and leading to the decrease in sugarcane productivity6.
Mosaic on sugarcane could present as a consequence of nutritional deficiencies7 or plant virus infection3. In case of viral disease, mosaic symptom on sugarcane probably caused by Sugarcane streak mosaic virus (SCSMV)8, Sorghum mosaic virus (SrMV)9, Sugarcane bacilliform virus10 or Sugarcane mosaic virus and morphologically difficult to distinguish. Therefore, the accurate detection of the causative agent of sugarcane mosaic disease is necessary to perform by using molecular approach3,11.
The SCMV is a member of the genus Potyvirus (family of Potyviridae)5 has long rod shape (flexuous rod) particle with the length of about 750 nm and diameter12 of 11-13 nm. It has about 900 bp sequence encodes capsid protein representing about 34 kDa protein11.
In some cases, capsid of the SCMV is used as necessary information to perform diagnosis and detection of the pathogen such as; RT-PCR that employ a set primer PCR using capsid protein-based nucleotide sequence3 or western blotting11 and enzyme-linked immunosorbent assay (ELISA) that use coat protein-based antibody13,14. According to the condition of unavailability data of SCMV distribution in comparison, on GM and non-GM sugarcane in Indonesia, it is necessary to observe and evaluate the variety against the distribution of SCMV. In addition, an approved genetically modified (GM) sugarcane NXI-4T that cultivated in the field beside others local varieties has not been evaluated for their response against important sugarcane pathogen, the SCMV. Therefore, this research aimed to monitor the SCMV distribution among local varieties recently cultivated in East Java.
MATERIALS AND METHODS
Determination of sampling locations, observation technique and disease incidence: Observation on sugarcane crops was carried out in October-November, 2017 in the area under the authority of Sugarcane Plantation company of Nusantara XI in East Java at four sugar factories (PG) plantation such as; PG Djatiroto in Lumajang, PG Semboro in Jember, PG Prajekan in Bondowoso and PG Asembagus in Situbondo. The climatological information around the plantation areas was provided according to the Indonesian Agency for Meteorology, Climatology and Geophysics (BMKG) information (Table 1). About four sugarcane varieties (Asembagus, Prajekan, Semboro, Jatiroto) were selected as samples and about 10% of the field at each plantation was observed and assessed for disease incidence3. Observations were made on local varieties that were widely grown, namely Bululawang, HW Merah, NXI 1-3 and a GM sugarcane of NXI-4T.
SCMV detection using reverse transcription polymerase chain reaction (RT-PCR): The presence of SCMV was detected from sugarcane leaves (healthy and symptomatic leaf) through RT-PCR protocol. Briefly, the complementary DNA (synthesized by using BioRad Reverse Transcriptase Polymerase Chains Reaction Kit) was subjected to a PCR reaction (Promega corporate, USA) using a specific pair-primer of capsid protein SCMV of cp-SCMV-F of 5’-CCC CAT ATG ACA GTC GAT GCA GGT GCT-3’ and cp-SCMV-R of 5’-ATG GAT CCT AGT GGT GCT GCT GCA CTC CC-3’ (Macrogene, Geumchun-gu, Seoul Korea)3.
| Table 1: | Climatological information |
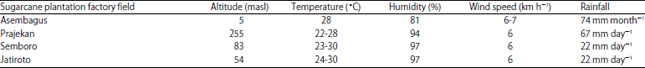 | |
| Source: Data were obtained from http://bmkg.go.id, masl: Meters above sea level | |
The thermal cycling condition of PCR was initial denaturation at 95°C for 3 min, denaturation at 95°C for 30 sec, annealing at 63°C for 20 sec and extension at 7°C for 60 sec, with a final extension at 72°C for 5 min. Detection of actin gene by PCR was done by using primer β-actin forward primer of 5’-GAA TTG CCT GAT GGA CGG T-3’ and reverse primer of 5’-GCT TTG GGA TCC ACA TCT A-3’ for checking the quality of cDNA15. PCR amplified product was visualized by using gel electrophoresis on 1% gel agarose.
Western blot analysis: The expression levels of the capsid protein of SCMV (cpSCMV) were detected by western blot. Briefly, the insoluble protein was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (12.5% acrylamides) and transferred to polyvinylidene fluoride (PDVF) membrane at 0.1 A, 4 W, 25 V for 2 h. The membrane was washed with Tris Buffered Saline (TBS) three times and blocked with 1% of skim milk in TBS for 30 min. The membrane was incubated for overnight in appropriate dilutions of CP SCMV antiserum at 28°C and washed with TBS three times. The membrane was incubated with secondary antibody in 1% skim milk for 1 h and followed by washing the membrane with TBS three times. The membrane was incubated in chromogenic dye (consist of 16.5 μL of 5-bromo-4-chloro-3-indolyl-phosphate (BCIP) and 33 μL of nitro blue tetrazolium (NBT) for every 5 mL of alkaline phosphate buffer for about 1.5 h at room temperature or until chromogenic bands appeared on the membrane11.
Estimation of total chlorophyll: Chlorophyll was extracted from about 2 g of finely cut leaves16. The leaves were taken and grounded with 2 mL of 80% acetone and centrifuged at 10,000 rpm for 10 min. The supernatant was transferred and the procedure was repeated until the residue becomes colorless. The absorbance was read at dual wavelengths3 of 645 and 663 nm against the solvent (acetone) blank. The estimation of total chlorophyll was calculated by following equation:
Chlorophyll a (mg g–1 FW) = [(12.7×A663)-(2.69×A645)]×0.5
Chlorophyll b (mg g–1 FW) = [(22.9×A645)-(4.69×A663)]×0.5
While17:
Total chlorophyll (mg g–1 FW) = [(20.2×A645)-(8.02×A663)]×0.5
Statistical analysis: The quantitative data were subjected to one way analysis of variance (ANOVA) with a completely randomized design (CRD) at a 95% confidence level. If the ANOVA results show significant differences, the analysis continued using Duncan's new multiple range test (DMRT) test at a 95% confidence levels.
RESULTS
Observation and sampling: The visual observations showed straight parallel yellow lines dispersed throughout the green leaf area, especially in the youngest leaves at the base of the leaf. However, there were some levels of symptom from the less until the most severe symptoms in all the observed plantations (Fig. 1a). In addition, there were variations of mosaics observed in each plantation, presumably mixed with other symptoms of the disease (Fig. 1b).
Disease incidence: Estimating data of the incidence of mosaic disease showed that only NXI-4T varieties, a GM sugarcane, had a higher incidence of a mosaic than other varieties. Mostly, all non-GM sugarcane had mosaic below 20% of incidence while the GM sugarcane had about 18.53% of mosaic incidence (Fig. 2).
Total chlorophyll contents: The results of chlorophyll contents analysis showed that there was a decrease in chlorophyll contents in symptomatic leaves (from fields) compared to non-symptomatic leaves (controls). The value of chlorophyll a, b and the total chlorophyll content in controls leaves were ranging from 17-57 mg g–1. Whereas in symptomatic (infected) leaves the chlorophyll a, b and total chlorophyll values were ranging from 13-47 mg g–1 (Fig. 3).
Detection SCMV using RT-PCR and western blot: The RT-PCR was performed to diagnose the possible causative SCMV using specific cp SCMV-gene PCR primer. All symptomatic mosaic plants was confirmed by RT-PCR which showed a single band with a size of 900 bp for coat protein of SCMV and 580 bp for actin (Fig. 4a). Indeed, the serological test using western blot result showed that all proteins from all symptomatic leaves produced a corresponding signal ranging ~37 kDa detected using polyclonal antibody of cpSCMV reaction (Fig. 4b) after staining with BCIP and NBT chromatography dye.
 | |
| Fig. 1(a-b): | Mosaic observation sugarcane leaves in selected plantations, (a) Level of the mosaic symptom on leaves from all sugarcane plantations and (b) Appearances of mosaic symptoms in 10 plantations |
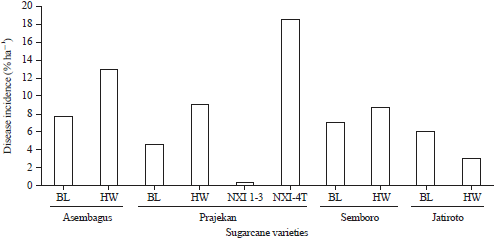 | |
| Fig. 2: | Estimation of disease incidence in several cultivars of sugarcane in the field and various mosaic symptoms |
DISCUSSION
In this study, the main symptom was recorded on sugarcane cultivars exhibiting a type of pattern of contrasting greenish within the island of green or yellowish chlorotic areas along with leaf especially at the base of the leaf3. There was a symptom of nutrition deficiency on the variety of HW Merah (HW) from Prajekan producing a mosaic covering the entire surface of the leaf, especially on the edge of the leaf.
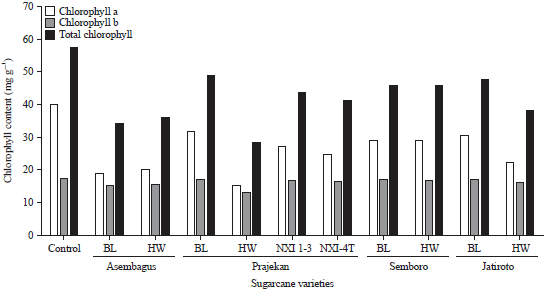 | |
| Fig. 3: | Estimation of chlorophyll a, chlorophyll b and total chlorophyll of non-symptomatic sugarcane leaves (control) and symptomatic sugarcane leaves from the plantation |
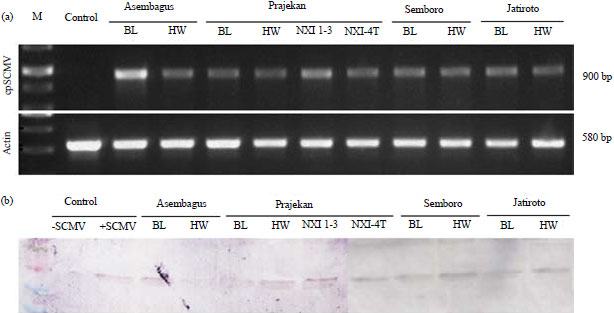 | |
| Fig. 4(a-b): | Confirmation of causative mosaic in symptomatic sugarcane leaf, (a) RT-PCR of the non-symptomatic leaf (line 1) and symptomatic leaf (line 2-11) producing a single band with a size of 900 bp (A, upper) responsible for coat protein of SCMV and 580 bp (A, lower) responsible for actin and (b) A serological test using the western blot technique showed the size of the protein band ~37 kDa |
Line 1: Negative control (non-symptomatic leaf), Line 2: Positive leaf infection and Line 3-12: Symptomatic leaves from sugarcane plantation (B) | |
Such symptoms are similar to the lack of water7 or nitrogen deficiency18, as well as on the variety of Bululawang (BL) from PG Jatiroto that exhibited mixed-symptoms similar to leaf rust19.
According to the disease incidence on NXI-4T (genetically modified sugarcane) compared to its ancestral sugarcane (Bululawang) indicated that the GM-sugarcane was more suffer than the ancestor (Fig. 2). It was studied that the variety of Bululawang resisted against mosaic disease20. However, when this cultivar is genetically modified into drought-resistant sugarcane by inserting choline dehydrogenase (betA) resulting moderate level in resistance status4. This possibly occurs since NXI-4T is drought-resistant genetically modified sugarcane expressed glycine cocamidopropyl betaine (an osmoprotectant compound) which increases the accumulation of glycine in leaves being responsible for defend mechanisms against environmental stress4,21. Nevertheless, the accumulation of glycine betaine on the leaves can chemically attract aphids (Aphis sp.), which is a vector of SCMV22. Furthermore, decreasing of chlorophyll content indicated that virion particles in mesophyll tissue damage chloroplast evoke mosaic pattern. A viral infection is suspected of causing structural changes in chloroplasts6, resulting in obstruction of chlorophyll synthesis23. This condition will result in changes in chlorophyll pigmentation, photosynthetic efficiency and accumulation of photosynthesis results3.
On the other hand, confirming the causative pathogen of mosaic in sugarcane could be done by various techniques such as by using serological approach like Western blot11, enzyme-linked immunosorbent assay (ELISA) or using molecular approach like reverse transcriptase-polymerase chain reaction (RT-PCR)3,11 continued by analysis of the capsid protein sequence3. In most cases, capsid protein-based gene is generally used to identify and detect the pathogen of mosaic diseases in the plant such as detection of SCMV on sugarcane in Pakistan, China and in Indonesia9,11,24.
Detection of the virus presence in the plant by using the RT-PCR technique is done through an approach to detect the presence of replicated viral RNA in the tissues of infected plants using a specific pair-primer for viral capsid protein. In case of detection of SCMV, a 900 bp of DNA fragment of capsid protein has been successfully amplified through RT-PCR procedure in confirming the presence of SCMV in sugarcane3. The similar result was also reported in detecting SCMV by using RT-PCR technique11,25,26. In addition, confirmation of virus-infected plant, in some cases could be done by western blot analysis using an immunoreaction of antibody and antigen. It has been widely used to confirm the presence of the virus based on the serological approach by using capsid protein-based antibody. Western blot has been successfully used in some reports to detect plant viruses. This technique has been successfully used to detect Maize chlorotic mottle virus (MCMV)13, to detect Citrus psorosis virus (CPsV) including to detect SCMV11,14. In addition, through this protocol, the protein ranged from 36.7-37 kDa representing the size of the capsid protein of SCMV11. In summary, SCMV is reported to infect both GM and non-GM-sugarcane. Interestingly, the disease incidence was high in GM-sugarcane than non-GM-sugarcane. It is interesting to confirm and reveal the molecular pathway or information of the relationship between GM-sugarcane (drought tolerant sugarcane) and the degree of mosaic incidence caused by the SCMV.
CONCLUSION
This study concluded that the NXI-4T (genetically modified sugarcane) only had a higher incidence of a mosaic than other varieties so the sugarcane NXI-4T was less resistant with the incidence above 20% against the SCMV. The SCMV was also decreasing the total chlorophyll content in infected leaves.
SIGNIFICANCE STATEMENT
This study for the first time reported the monitoring of SCMV on genetically modified sugarcane cultivated in Indonesia and discovered the useful information for farmer and industry to choose suitable sugarcane and develop new resistant sugarcane against SCMV.
ACKNOWLEDGMENT
This research was a part of partially funding research scheme by the Ministry of Research, Technology and Higher Education in addition to a Research grant funded by the Islamic Development Bank (IsDB) Project of the University of Jember with grant number of 200/UN25.7/PIU-IDB/2017 assigned to BS.
REFERENCES
- Huckett, B.I. and F.C. Botha, 1996. Progress towards a definitive diagnostic test for sugarcane mosaic virus infection. Proc. S. Afr. Sug. Technol. Ass., 70: 11-13.
Direct Link - Putra, L.K., H.J. Ogle, A.P. James and P.J. Whittle, 2003. Distribution of Sugarcane mosaic virus in sugarcane plants. Aust. Plant Pathol., 32: 305-307.
CrossRefDirect Link - Addy, H.S., Nurmalasari, A.H.S. Wahyudi, A. Sholeh and C. Anugrah et al., 2017. Detection and response of sugarcane against the infection of Sugarcane mosaic virus (SCMV) in Indonesia. Agronomy, Vol. 7, No. 3.
CrossRefDirect Link - Xie, X., W. Chen, Q. Fu, P. Zhang, T. An, A. Cui and D. An, 2016. Molecular variability and distribution of Sugarcane mosaic virus in Shanxi, China. PLoS One, Vol. 11.
CrossRefDirect Link - Zhao, J., X. Zhang, Y. Hong and Y. Liu, 2016. Chloroplast in plant-virus interaction. Front. Microbiol., Vol. 7.
CrossRefDirect Link - Ferreira, T.H., M.S. Tsunada, D. Bassi, P. Araujo and L. Mattiello et al., 2017. Sugarcane water stress tolerance mechanisms and its implications on developing biotechnology solutions. Front. Plant Sci., Vol. 8: 1077.
CrossRefDirect Link - Fu, W.L., S.R. Sun, H.Y. Fu, R.K. Chen, J.W. Su and S.J. Gao, 2015. A one-step real-time RT-PCR assay for the detection and quantitation of Sugarcane streak mosaic virus. BioMed. Res. Int., Vol. 2015.
CrossRefDirect Link - Xu, D.L., J.W. Park, T.E. Mirkov and G.H. Zhou, 2008. Viruses causing mosaic disease in sugarcane and their genetic diversity in southern China. Arch. Virol., 153: 1031-1039.
CrossRefDirect Link - Karuppaiah, R., R. Viswanathan and V.G. Kumar, 2013. Genetic diversity of Sugarcane bacilliform virus isolates infecting Saccharum spp. in India. Virus Genes, 46: 505-516.
CrossRefDirect Link - Darsono, N., N.N. Azizah, K.M. Putranty, N.T. Astuti, H.S. Addy, W. Darmanto and B. Sugiharto, 2018. Production of a polyclonal antibody against the recombinant coat protein of the Sugarcane mosaic virus and its application in the immunodiagnostic of sugarcane. Agronomy, Vol. 8.
CrossRefDirect Link - Chaves-Bedoya, G. and L.Y. Ortiz-Rojas, 2012. Evidence of different phylogenetic origins of two mexican Sugarcane mosaic virus (SCMV) isolates. Acta Agron., 61: 79-87.
Direct Link - Wang, Q., C. Zhang, C. Wang, Y. Qian, Z. Li, J. Hong and X. Zhou, 2017. Further characterization of Maize chlorotic mottle virus and its synergistic interaction with Sugarcane mosaic virus in maize. Sci. Rep., Vol. 7.
CrossRefDirect Link - Salem, R., I.A. Arif, M. Salama and G.E.H. Osman, 2018. Polyclonal antibodies against the recombinantly expressed coat protein of the Citrus psorosis virus. Saudi J. Biol. Sci., 25: 733-738.
CrossRefDirect Link - Kumar, R., A.K. Singh, D. Lavania, M.H. Siddiqui, M.H. Al-Whaibi and A. Grover, 2016. Expression analysis of ClpB/Hsp100 gene in faba bean (Vicia faba L.) plants in response to heat stress. Saudi J. Biol. Sci., 23: 243-247.
CrossRefDirect Link - Su, S., Y. Zhou, J.G. Qin, W. Yao and Z. Ma, 2010. Optimization of the method for Chlorophyll extraction in aquatic plants. J. Freshwater Ecol., 25: 531-538.
CrossRefDirect Link - Molazem, D., E.M. Qurbanov and S.A. Dunyamaliyev, 2010. Role of proline, Na and chlorophyll content in salt tolerance of corn (Zea mays L.). Am.-Eurasian J. Agric. Environ. Sci., 9: 319-324.
Direct Link - Hartt, C.E., 1970. Effect of nitrogen deficiency upon translocation of 14C in sugarcane. Plant Physiol., 46: 419-422.
CrossRefDirect Link - Briggs, G.C., Z. Nakhid, A.T. Alleyne, J. Ayats, J.O. Despradel and W. Elibox, 2014. First report of orange rust disease of sugarcane in the dominican republic. Plant Dis., 98: 1010-1010.
CrossRefDirect Link - Cheng, Y.J., X.P. Deng, S.S. Kwak, W. Chen and A.E. Eneji, 2013. Enhanced tolerance of transgenic potato plants expressing choline oxidase in chloroplasts against water stress. Bot. Stud., Vol. 54.
CrossRefDirect Link - Zúñiga, G.E. and L.J. Corcuera, 1987. Glycine-betaine accumulation influences susceptibility of water-stressed barley to the aphid Schizaphis graminum. Phytochemistry, 26: 367-369.
CrossRefDirect Link - Pazarlar, S., M. Gumus and G.B. Oztekin, 2013. The effects of Tobacco mosaic virus infection on growth and physiological parameters in some pepper varieties (Capsicum annuum L.). Not. Bot. Hortic. Agrobotanici Cluj-Napoca, 41: 427-433.
CrossRefDirect Link - Haider, M.S., S. Afghan, H. Riaz, M. Tahir, M.A. Javed, N. Rashid and J. Iqbal, 2011. Identification of two Sugarcane mosaic virus (SCMV) variants from naturally infected sugarcane crop in Pakistan. Pak. J. Bot., 43: 1157-1162.
Direct Link - Viswanathan, R., R. Karuppaiah and M. Balamuralikrishnan, 2010. Detection of three major RNA viruses infecting sugarcane by multiplex reverse transcription-polymerase chain reaction (multiplex-RT-PCR). Aust. Plant Pathol., 39: 79-84.
CrossRefDirect Link - Gonçalves, M.C., D.M. Galdeano, I.D.G. Maia and C.M. Chagas, 2011. Genetic variability of Sugarcane mosaic virus causing maize mosaic in Brazil. Pesquisa Agropecuária Bras., 46: 362-369.
CrossRefDirect Link