
Saad Farouk
Department of Agricultural Botany, Faculty of Agriculture, Mansoura University, Egypt
LiveDNA: 20.2011
Abu Bakr Al-Jilani Sanusi
Department of Plant Production, Faculty of Agriculture, Sirte University, Libya
Journal of Applied Sciences
Year: 2019 | Volume: 19 | Issue: 4 | Page No.: 270-278







ABSTRACT
Background and Objective: wheat is an annual crop that needs cold and long day to transition to flowering that is essential in cultivated cereal to maximize yield. But little is reported in flowering inducing substances on flowering induction in spring wheat. Materials and Methods: Two field experiments were done in the Agric Bot. Dept., Mansoura Univ., Egypt, to assess the role of vernalization (VER) and flowering inducing substances 'FIS' (ascorbic acid 'AsA' and benzyl adenine 'BA') application as well as their interaction on wheat growth, flowering yield and grain quality. Results: Thirty days VER treatment increased plant growth, improved physiological characteristics, induced flowering by modulating plant phytohormone concentration and finally increased yield and grain quality. Exogenous application of 75 mg L–1 AsA plus 50 μM BA under normal or VER treatment increased all studied characters. Conclusion: Application of 75 mg L–1 AsA plus 50 μM BA with 30 days VER three times 30, 40 and 50 days from sowing is suggested to achieve the uppermost growth and yield.
PDF Abstract XML References Citation
Received: January 10, 2019;
Accepted: January 30, 2019;
Published: April 12, 2019
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Saad Farouk and Abu Bakr Al-Jilani Sanusi, 2019. Potent Induction of Wheat Flowering and its Related to Yield Components. Journal of Applied Sciences, 19: 270-278.
DOI: 10.3923/jas.2019.270.278
URL: https://scialert.net/abstract/?doi=jas.2019.270.278
DOI: 10.3923/jas.2019.270.278
URL: https://scialert.net/abstract/?doi=jas.2019.270.278
INTRODUCTION
Wheat (Triticum aestivum L.) is the premier staple crop worldwide and counted among the 'big three' cereal crops, accounting 216.8 million ha with a production of 651.4 million t being harvested annually1. In Egypt and Libya, wheat is considered the first strategic food crop for human and animals. The magnitude of wheat lies in it's vital nutritional and industrial commodity in which proteins and carbohydrates, it provides 37% of the total calories for people and 40% of the protein in the Egyptian diet. Remarkable increment in wheat production from 19.64 million t ha–1 in 2009 to 22.88 million t ha–1 in 2015, was achieved by raising the wheat area from 1.32 to 1.46 million ha year–1 and grain yield from 6.45 to 6.60 t fed–1 in the same period. There is a gap between wheat consumption and production, hence, using suitable agricultural practices such as vernalization (VER) and foliar spraying with flowering inducing substances (FIS) may have an effect on wheat productivity and grain quality.
During plant development, the timing of the transition from vegetative to reproductive growth is crucial for reproductive success and maximizing grain yield. Recently, vernalization has been examined widely at the biochemical and molecular levels2. Gardner and Barnett3 classified wheat cultivars in response to VER into three types of qualitative (6-8 weeks at 6°C), quantitative (2-4 weeks at 2°C) and neutral. However, according to Wang et al.4, the classification relies further on cultivation date, reliable with a cultivar, rather than the nature of the VER response of that cultivar, thus some spring varieties may require VER or in contrary, winter varieties may necessitate an extended VER. Plant responses to VER have been reported in some field crops and it has been assessed that VER accelerate plant growth and development as well as improves biochemical attributes, raising the yield of several plants5-7.
Ascorbic acid (AsA) regulates various crucial aspects of physiological processes like cellular division, growth and phytohormone-mediated signaling processes throughout the flowering stage8. The beneficial effects of ascorbic acid upon growth and productivity as well as some biochemical attributes have been reported on several field crops9-11. In this concern, Hamed and Abd Elgawas12 proved that there were a progressive increase in growth, yield and its attributes, grain quality and photosynthetic pigments by the AsA application.
Cytokinins regulate flowering genes in shoot apical meristem, leading to floral stimulus13 and play a vital role in the greater partitioning of photosynthates towards reproductive sink thus raising the yield14. Generally, during flowering transition and development, there is a massive concentration of cytokinin observed in the apical meristem of many plants15. Benzylaminopurine (BAP) or 6-benzyladenine (6-BA) was shown to increase plant growth and biochemical attributes as well as yield components16.
In higher plants, flowering time is tightly manipulated by multi-internal or external factors. The timing of flowering affects total yield as well as fruit quality. There are no applicable data in the compartment effect of FIS and VER on spring wheat flowering. Understanding the regulatory mechanisms by which these factors modulate flowering time is very helpful when developing management strategies for wheat crop production.
The aim of the present investigation was to clarify the role of FIS with or without VER treatments on vegetative growth, some physiological attributes, flowering and finally yield and grain quality in wheat.
MATERIALS AND METHODS
Plant materials and experimental location: Two field experiments were done at Agricultural Botany Department Experimental Farm, Mansoura University, during 2014/2015 and 2015/2016 winter season to assess the influence of vernalization (VER), ascorbic acid (AsA), benzyladenine (BA) and their interaction on wheat crop growth, earliness and yield and grain quality. Wheat grains (Triticum aestivum L. cv Gemmeza 9) were secured from the Field Crop Research Institute, ARC, Egypt. Random soil samples were collected from the experimental soil at the depth of 0-30 cm before planting for the physical and chemical properties of soil according to methods delineated by Motsara and Roy17. Data of physical and chemical analysis are shown in Table 1.
Layout of the experimental design and plant management: A split-plot design with three replicates was followed VER treatment occupying the main plots, whereas foliar spraying with flowering inducing substances (FIS) was allocated in the sub-plots. The plot size was 3 m2 (1.5 m in length and 2 m width) including 13 rows.
The experimental soil was regularly prepared and fertilized according to the recommendation of the Ministry of Agriculture and Land Reclamation, Egypt. Nutrient doses of 119 and 119 kg P2O5 ha–1 as superphosphate (15.5% P2O5) or potassium sulfate (48% K2O) were added previously to sowing, while nitrogen was supplied at a rate of 238 kg N ha–1 as ammonium nitrate (33.5% N) four times, the first one 21 days from sowing (DFS) and then every 10 days.
| Table 1: | Physical and chemical analysis of the experimental soil in the first and second season |
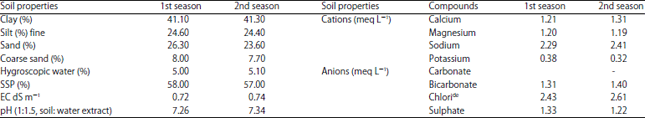 | |
For VER treatment, the grains were divided into three groups, the first one stored in the lab conditions, while the other two groups were placed in wet peat in perforated plastic bags, then exposed to low temperatures at 4-5°C in the refrigerator for 30 or 45 days before sowing date. Wheat grains were sown at the rate of 168 kg ha–1 on the 10th and 15th November in 15 cm row to row distances by the single row handheld drill.
The plants were sprayed three times at 30, 40 and 50 DFS once adding 0.05% (v/v) Tween 20 as a wetting agent, according to the following treatments (1) Control (water); (2)75 mg L–1AsA, (3)50 μM BA and (4)75 mg L–1 AsA plus 50 μM BA.
Sampling and measurements: Random samples of 10 plants were chosen from every experimental plot at 90 DFS in each season for assessing the growth characters and some physiological attributes.
Vegetative growth characteristics:
| • | Plant height (cm) was measured from the soil surface to the tip of the largest linear blade in the plant |
| • | Leaf area per plant (cm2) was estimated using the leaf area meter, AM 300 150 (ADC Bioscientific Ltd) |
| • | Shoot dry weight (g) |
| • | No of tillers per plant |
Physiological trails in the second season: The method of Lichtenthaler and Wellburn18 was used for the estimation of photosynthetic pigments in the flag leaf using methanol supplemented with a trace of potassium carbonate as extraction solvent.
Total and soluble carbohydrates were extracted and determined according to the method of Sadasivam and Manickam19.
Ion percentage was estimated once wet digesting. Total nitrogen was estimated by the micro-Kjeldahl method, potassium was determined by flame photometry17, total phosphorus was assayed by the ammonium molybdate and ascorbic acid method20 using a spectrophotometer. Flag leaf relative water content was estimated by the procedure of Bajji et al.21.
Membrane permeability (%) was estimated following the method of Wang et al.22.
Endogenous phytohormones concentration in the flag leaf was determined by the method of Shindy and Smith23.
Yield and its components: At harvest, the yield and its components were estimated as spike length (cm), grain yield/plant, number of grains per spike, grain yield per plant and grain index (1000 grain weight). Additionally, determination of grain protein and carbohydrate percent19.
Statistical analysis: Data were presented as mean±SD of 10 replicates and subjected to two-way analysis of variance using CoHort Software, 2008 statistical package. For means comparisons, analysis was used Duncan's multiple range test methods (DMRT) at p<0.05 significance level.
RESULTS
Plant growth: Data in Table 2 shows that growth characters of wheat plants were significantly influenced by either vernalization "VER" or foliar application of flowering inducing substances "FIS" and their interaction. The data indicated that VER treatment significantly increased all plant growth in both season compared with control plants. On the other hand, all growth characters increased under FIS foliar application, in special, application of 75 mg L–1 AsA plus 50 μM BA which increased plant height by 14 and 8%, tillers number per plant by 58 and 45%; leaf area per plant by 25 and 19% and shoot dry weight by 35 and 41% in the first and second season respectively. Regarding the interaction effect, the data in the same table suggest that exogenous application of FIS has an additive effect under VER treatment.
Photosynthetic pigment and carbohydrates concentration: Grain pre-treatment with VER, in special, for 30 days before sowing, significantly raised the total chlorophyll, total carotenoids, total carbohydrates and total soluble carbohydrates concentration and decreased the ratio between chlorophyll a to chlorophyll b in the flag leaf relative to un-vernalized treatments (Table 3).
| Table 2: | Wheat plant growth as affected by vernalization, flowering inducing substances and their interaction at 90 days from sowing in both seasons |
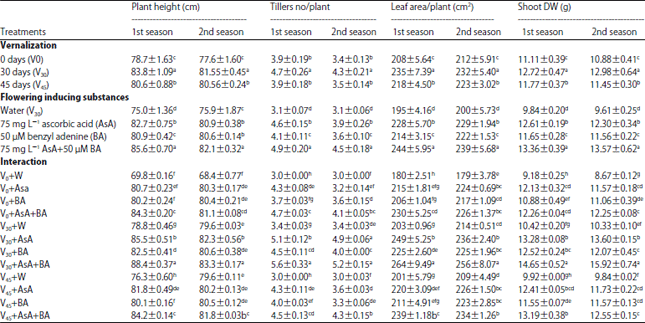 | |
| Values are given as Mean±SD of three replicate. Means in columns with different letters are significantly different at p<0.05 (Duncan's multiple range test) | |
| Table 3: | Photosynthetic pigments, total and soluble carbohydrates of wheat flag leaf as affected by vernalization, flowering inducing substances and their interaction at 90 days from sowing in the second season |
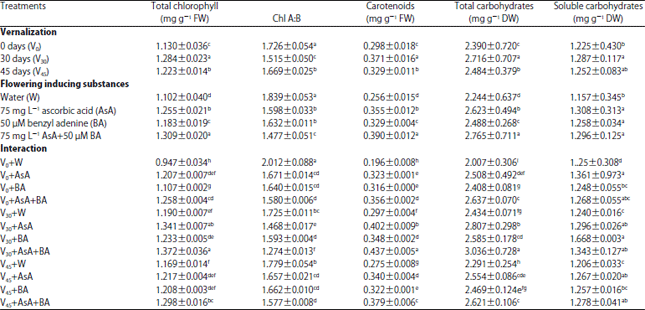 | |
| Values are given as Mean±SD of three replicate. Means in columns with different letters are significantly different at p<0.05 (Duncan's multiple range test) | |
Data also showed that the highest concentration of chlorophyll, carotenoids, total carbohydrate and soluble carbohydrate were obtained under foliar application of 75 mg L–1 AsA plus 50 μM BA which raised total chlorophyll, total carotenoids, total carbohydrates and total soluble sugars by 18 m, 52 m, 23 m and 12% relative to control, respectively. This treatment also decreased the ratio between chlorophyll a to chlorophyll b by 19%. Regarding the interaction effects, the data in Table 3 clearly indicated that the most effective interaction that gave the highest values relative to control was the foliar application of 75 mg L–1 AsA plus 50 μM BA combined with 30 days VER.
| Table 4: | Nitrogen (N), phosphorous (P), potassium (K), relative water content (RWC) and membrane permeability (MP) percentage of wheat flag leaf as affected by vernalization, flowering inducing substances and their interaction at 90 days from sowing in the second season |
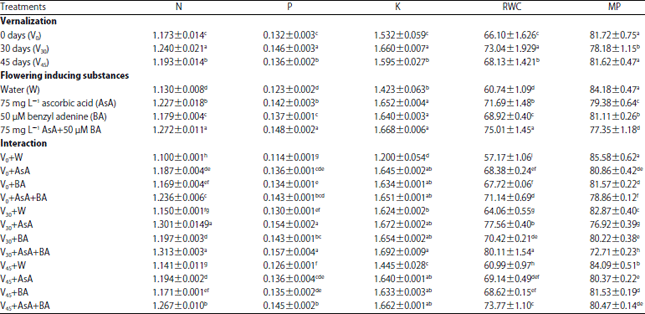 | |
| Values are given as Mean±SE of three replicate. Means in columns by different letters are significantly different at p<0.05 by (Duncan's Multiple Range Test) | |
| Table 5: | Flag leaf endogenous phytohormones concentration (ng g–1 FW) and days to heading of wheat as affected by vernalization, flowering inducing substances and their interaction at 90 days from sowing in the second season |
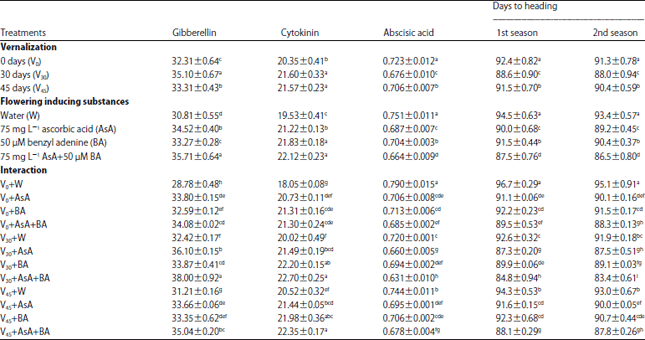 | |
| Values are given as Mean±SE of three replicate. Means in columns by different letters are significantly different at p<0.05 by (Duncan's Multiple Range Test) | |
Ion, relative water content and membrane permeability percentages: Ion and relative water content percentages significantly raised by 30 days VER treatment which increased N, P, K and RWC (%) by 5, 10, 5 and 10%, respectively.
Alternatively, VER treatment decreased membrane permeability percent (Table 4). Concerning the role of FIS on ion and RWC (%), Table 4 verified that foliar spraying of 75 mg L–1 AsA plus 50 μM BA gave the highest percent of N, P, K and RWC relative to untreated plants reaching to 12, 20, 17 and 23%, but decreased membrane permeability percent by 8%.
Endogenous phytohormones and number of days to heading: Data in Table 5 indicated that the number of days to heading significantly decreased by VER treatment in special 30 days, that was associated to increased the concentration of GA and cytokinin likewise as decreased ABA concentration in flag leaf of wheat plants. The data in the Table 5 showed that exogenous application of FIS increased the concentration of GA and cytokinin, however, decreased ABA resulting in shortage the time requisite to heading. The most effective treatment in this concern was 75 mg L–1 AsA plus 50 μM BA that decreased the time for heading by about 7 days relative to untreated control plants. This decrease in the time needed for heading was associated with increasing GA and cytokinin by 15 and 13%, also decreasing the ABA concentration by 11%. Compared to untreated plants, exogenous application of FIS in special 75 mg L–1 AsA plus 50 μM BA treatment considerable decline the number of days to heading by about 11 days. This decrease resulted from increasing the concentration of either GA or cytokinin and declines the concentration of ABA by 32, 25 and 20%, respectively.
Yield and its components: Yield and grain quality were significantly increased under vernalization treatment, in special for 30 days which gave the highest values compared with not-vernalized plants in each experimental season (Table 6). Exogenous application of FIS in special, 75 mg L–1 AsA plus 50 μM BA, significantly increased yield and grain quality, relative to untreated plants (Table 6). Data in the same Table showed that the interaction impact between vernalization and FIS foliar application gave an accumulative effect.
DISCUSSION
The current investigation verified that application of flowering inducing substances (FIS) with or without vernalization (VER), accelerates flowering induction of spring wheat and increased plant growth and yield and improved grain quality. These findings are in agreement with earlier reported5,6,9,10,12,16. The improvement in yield and grains quality of wheat under the present investigation may be a result of their responding increased in leaf area and tiller numbers per plant, photosynthetic pigment concentration, enhanced photosynthetic rate, coupled with an increment in sub-stomatal CO2, as well as increased rates of assimilate transport from the source to the developing grains24. However, increase yield might be due to more grains per spike and increased assimilate partitioning from leaves to grains, as suggested by the 1000-grain weight5,6. Additionally, all experimental treatment increased the concentration of GA and/or cytokinin and decreased ABA as well as increasing the availability of photoassimilate transport to developing grains, leading to raised fruit set and improved grain quality25. All experimental treatments improved considerably grain quality represented as increasing protein and carbohydrate percent in the wheat grains relative to the untreated ones. In this concern, Mohamed26 proved that application of AsA improved wheat grain quality as compared to control plants. Application of BA increased grain protein and total carbohydrates percentage27. It can be suggested here that the application of AsA or BA encourages the absorption of nitrogen from the soil and/or improved the photosynthetic processes through their impact on some enzymatic action. The activation of these processes might cause the rise in protein buildup and carbohydrate accumulation in the developing grains.
Throughout plant development, the timing of the transition from vegetative to reproductive phase is important, in cultivated cereals, to maximize grain production. Upon starting this transition and thus have evolved a series of mechanisms by which flowering time can be specifically synchronized with environmental cues. However, according to Wang et al.,4 the classification of wheat cultivars as spring and winter, relies further on cultivation date consistent with a cultivar, rather than the nature of the VER response of that cultivar, so some spring varieties may need VER or in contrary, winter varieties may require an extended VER.
Gardner and Barnett3 calculated VER of wheat varieties in terms of a quantity called the chilling unit and declared that the amount of chilling unit required to fulfill VER requirement is natural in the field. According to these researchers3, high fluctuation in temperatures in the field can increase the effect of coldness on the fulfillment of VER requirement of wheat, this occurs due to an increase in chilling units for fulfilling VER requirement in the refrigerator (compared to the field). In temperate cereals, such as wheat and barley, the main genes regulating the VER response are VERNALIZATION1 (VRN1)6, VERNALIZATION2 (VRN2)28 and VERNALIZATION3 (VRN3)29. These genes are interconnected by complex epistatic combinations whereby an allele for spring growth habits every gene is epistatic to the alleles for winter growth habit in the other two genes28. The induction of VRN1 transcription by VER coincides with cold-induced changes in histone methylation and acetylation patterns in its regulatory regions30.
| Table 6: | Yield and yield components of wheat plant in both season as well as grain quality in the second season as affected by vernalization, flowering inducing substances and their interaction at harvesting time |
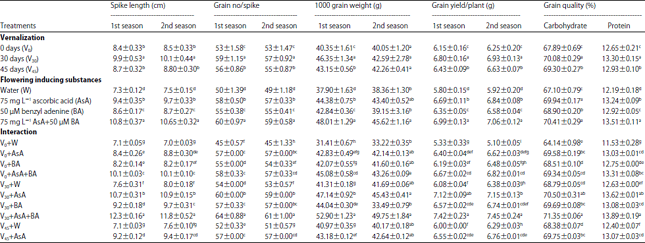 | |
| Values are given as Mean±SD of three replicate. Means in columns with different letters are significantly different at p<0.05 (Duncan's multiple range test) | |
The up regulation of VRN1 in the leaves is important to maintain low transcript levels of the flowering repressor VRN2 in the spring31, which facilitates the up-regulation of VRN3 and the initiation of the transition to the reproductive phase28. It was shown that winter wheat, rye and barley cultivars can quickly turn to heading only if germinated grains or plantlets were exposed to low temperature for a certain time at the onset of development.
AsA and/or BA and/or VER may play a crucial role in floral induction, through alterations in phytohormone levels, like decreasing ABA and enhanced GA and endogenous cytokinin. ABA may contribute to the down-regulation of LEAFY and thus to the late flowering phenotype, while GA plays a major role in promoting flowering. The GAs are a family of tetracyclic diterpenoid carboxylic acids that comprise several bioactive forms that regulation numerous aspects of plant's development, counting the initiation and promotion of flowering32. In wheat, GA has previously been reported to accelerate flowering in spring wheat under long days33. Moreover, circumstantial evidence that floret fertility was linked to an increment in the level of soluble carbohydrates in the shoot and spike that may accelerate the bio-synthesis of a floral stimulus that's transported through vascular tissues to the shoot apical meristem and subsequently induces flower initiation34. These finding verified that the exogenous application of AsA or BA may accelerate flower induction by raising photo-assimilate movement.
The increment of photosynthetic pigments due to FIS could be attributed to inhibition of chlorophyll degradation and/or the promotion of chlorophyll synthesis and/or accelerated the development and modification of the chloroplast35. Moreover, the increment in photosynthetic pigment under FIS might be attributed to efficient scavenging of ROS by antioxidant-enzymes and molecules; that would have destroyed chlorophylls36. Application of AsA, BA or VER decreased the Chl a:b ratio that plays a critical role in stabilizing photosynthetic processes. Such a decline in the Chla:b ratio seems to be a conflict with the fact that Chl a is comparatively stable throughout senescence, however chlorophyll b is almost labile.
Functionally, the decline in Chla:b ratio induced the capture of far-red radiation and helps to sustain an energy balance between PSI and PSII, resulting in optimal functioning37.
CONCLUSION
The present investigation proved that, spring wheat might be flowering earlier and gave the greatest grain yield by foliar spraying three times 30, 40 and 50 days from sowing with 75 mg L–1 AsA plus 50 μM BA under 30 days vernalization.
SIGNIFICANCE STATEMENT
The timing of flowering affects total yield and grain quality. The genetic elements and molecular mechanisms that rule in flowering process have been studied in diverse crops. But, reports on the responsibility of flowering inducing substances with or without vernalization for induction flowering in spring wheat are inadequate. The current investigation provides insight into how crops might be managed to increase wheat production by FIS and VER. Also the performed experiment is one of several steps on the way to explain the flowering mechanisms of spring wheat.
REFERENCES
- Kim, D.H., M.R. Doyle, S. Sung and R.M. Amasino, 2009. Vernalization: Winter and the timing of flowering in plants. Annu. Rev. Cell Dev. Biol., 25: 277-299.
CrossRefDirect Link - Gardner, F.P. and R.D. Barnett, 1990. Vernalization of wheat cultivars and a triticale. Crop Sci., 30: 166-169.
CrossRefDirect Link - Wang, S.Y., R.W. Ward, J.T. Ritchie, R.A. Fischer and U. Schulthess, 1995. Vernalization in wheat I. A model based on the interchangeability of plant age and vernalization duration. Field Crops Res., 41: 91-100.
CrossRefDirect Link - Steinfort, U., B. Trevaskis, S. Fukai, K.L. Bell and M.F. Dreccer, 2017. Vernalisation and photoperiod sensitivity in wheat: Impact on canopy development and yield components. Field Crops Res., 201: 108-121.
CrossRefDirect Link - Steinfort, U., S. Fukai, B. Trevaskis, D. Glassop, A. Chan and M.F. Dreccer, 2017. Vernalisation and photoperiod sensitivity in wheat: The response of floret fertility and grain number is affected by vernalisation status. Field Crops Res., 203: 243-255.
CrossRefDirect Link - Fukuda, M., Y. Yanai, Y. Nakano and T. Higashide, 2018. Differences in vernalisation responses in onion cultivars. J. Hortic. Sci. Biotechnol., 93: 316-322.
CrossRefDirect Link - Gallie, D.R., 2013. L-ascorbic acid: A multifunctional molecule supporting plant growth and development. Scientifica,.
CrossRefDirect Link - Farouk, S., 2011. Ascorbic acid and α-Tocopherol minimize salt-induced wheat leaf senescence. J. Stress Physiol. Biochem., 7: 58-79.
Direct Link - Hussein, M.M. and A.K. Alva, 2014. Effects of zinc and ascorbic acid application on the growth and photosynthetic pigments of millet plants grown under different salinity. Agric. Sci., 5: 1253-1260.
CrossRefDirect Link - Hamed, B.A. and H. Abd Elgawad, 2018. Differential effects of some ascorbates on Phaseolus vulgaris L. in response to salinity stress. Egypt. J. Bot., 58: 23-36.
Direct Link - Lindsay, D.L., V.K. Sawhney and P.C. Bonham-Smith, 2006. Cytokinin-induced changes in CLAVATA1 and WUSCHEL expression temporally coincide with altered floral development in Arabidopsis. Plant Sci., 170: 1111-1117.
CrossRefDirect Link - Sharma, K.M., D.D. Sharma, K.B. Shukla and B. Upadhyay, 2008. Growth partitioning and productivity of wheat as influenced by fertilization and foliar application of bio-regulators. Indian J. Plant Physiol., 13: 387-393.
Direct Link - Corbesier, L., E. Prinsen, A. Jacqmard, P. Lejeune, H.V. Onckelen, C. Perilleux and G. Bernier, 2003. Cytokinin levels in leaves, leaf exudate and shoot apical meristem of Arabidopsis thaliana during floral transition. J. Exp. Bot., 54: 2511-2517.
CrossRefDirect Link - Motsara, M.R. and R.N. Roy, 2008. Guide to Laboratory Establishment for Plant Nutrient Analysis. 1st Ed. Food and Agriculture Organization of the United Nations, Rome, Italy, ISBN: 9789251059814, Pages: 204.
Direct Link - Lichtenthaler, H.K. and A.R. Wellburn, 1985. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biol. Soc. Trans., 11: 591-592.
CrossRefDirect Link - Bajji, M., S. Lutts and J.M. Kient, 2001. Water deficit effects on solute contribution to osmotic adjustment as a function of leaf ageing in three durum wheat (Triticum durum Desf.) cultivars performing differently in arid conditions. Plant Sci., 160: 669-681.
Direct Link - Wang, Q., X. Liang, Y. Dong, L. Xu, X. Zhang, J. Hou and Z. Fan, 2013. Effects of exogenous nitric oxide on cadmium toxicity, element contents and antioxidative system in perennial ryegrass. Plant Growth Regul., 69: 11-20.
CrossRefDirect Link - Shindy, W.W. and O.E. Smith, 1975. Identification of plant hormones from cotton ovules. Plant Physiol., 55: 550-554.
CrossRefDirect Link - Ren, B., J. Zhang, S. Dong, P. Liu and B. Zhao, 2018. Exogenous 6‐benzyladenine improves antioxidative system and carbon metabolism of summer maize waterlogged in the field. J. Agron. Crop Sci., 204: 175-184.
CrossRefDirect Link - Mohamed, N.E.M., 2013. Behaviour of wheat Cv. Masr-1 plants to foliar application of some vitamins. Nat. Sci., 11: 1-5.
Direct Link - Ibrahim, S.M.M., S.T. Lobna and M.M. Farahat, 2010. Vegetative growth and chemical constituents of Croton plants as affected by foliar application of Benzyl adenine and Gibberellic acid. J. Am. Sci., 6: 126-130.
Direct Link - Distelfeld, A., C. Li and J. Dubcovsky, 2009. Regulation of flowering in temperate cereals. Curr. Opin. Plant Biol., 12: 178-184.
CrossRefDirect Link - Nitcher, R., A. Distelfeld, C. Tan, L. Yan and J. Dubcovsky, 2013. Increased copy number at the HvFT1 locus is associated with accelerated flowering time in barley. Mol. Genet. Genomics, 288: 261-275.
CrossRefDirect Link - Oliver, S.N., E.J. Finnegan, E.S. Dennis, W.J. Peacock and B. Trevaskis, 2009. Vernalization-induced flowering in cereals is associated with changes in histone methylation at the VERNALIZATION1 gene. Proc. Natl. Acad. Sci., 106: 8386-8391.
CrossRefDirect Link - Chen, A. and J. Dubcovsky, 2012. Wheat TILLING mutants show that the vernalization gene VRN1 down-regulates the flowering repressor VRN2 in leaves but is not essential for flowering. PLoS Genet., Vol. 8.
CrossRefDirect Link - Pearce, S., L.S. Vanzetti and J. Dubcovsky, 2013. Exogenous gibberellins induce wheat spike development under short days only in the presence of vernalization. Plant Physiol., 163: 1433-1445.
CrossRefDirect Link - Evans, L.T., C. Blundell and R.W. King, 1995. Developmental responses by tall and dwarf isogenic lines of spring wheat to applied gibberellins. Aust. J. Plant Physiol., 22: 365-371.
CrossRefDirect Link - Bernier, G. and C. Perilleux, 2005. A physiological overview of the genetics of flowering time control. Plant Biotechnol. J., 3: 3-16.
CrossRefDirect Link - Sudria, C., J. Palazon, R. Cusido, M. Bonfill, M.T. Pinol and C. Morales, 2001. Effect of benzyladenine and indolebutyric acid on ultrastructure, glands formation and essential oil accumulation in lavandula dentata plantlets. Biol. Planta., 44: 1-6.
CrossRefDirect Link - Davey, M.W., M.V. Montagu, D. Inze, M. Sanmartin and A. Kanellis et al., 2000. Plant L-ascorbic acid: Chemistry, function, metabolism, bioavailability and effects of processing. J. Sci. Food Agric., 80: 825-860.
CrossRefDirect Link