
Lawal Garba
Enzyme and Microbial Technology Research Centre, Faculty of Biotechnology and Biomolecular Sciences, University Putra Malaysia, 43400 UPM Serdang, Selangor Darul Ehsan, Malaysia
Mohd Shukuri Mohamad Ali
Enzyme and Microbial Technology Research Centre, Faculty of Biotechnology and Biomolecular Sciences, University Putra Malaysia, 43400 UPM Serdang, Selangor Darul Ehsan, Malaysia
Siti Nurbaya Oslan
Enzyme and Microbial Technology Research Centre, Faculty of Biotechnology and Biomolecular Sciences, University Putra Malaysia, 43400 UPM Serdang, Selangor Darul Ehsan, Malaysia
Raja Noor Zaliha Raja Binti Abdul Rahman
Enzyme and Microbial Technology Research Centre, Faculty of Biotechnology and Biomolecular Sciences, University Putra Malaysia, 43400 UPM Serdang, Selangor Darul Ehsan, Malaysia
Journal of Applied Sciences
Year: 2017 | Volume: 17 | Issue: 6 | Page No.: 282-295







ABSTRACT
Carbohydrates and proteins are polymers of polysaccharides and polypeptide residues, respectively. Conversely, lipids are made up of a wide range of compounds with tremendous differences in structures and lack building blocks. Therefore, fatty acids constitute the major components of various lipid classes such as glycerides and sphingolipids. Fatty acids are organic compounds containing a carboxylic acid group mostly at the end of an aliphatic chain and are categorized into saturated and unsaturated. Most fatty acids especially the so-called polyunsaturated fatty acids (PUFAs) and their derivatives play key biological roles in inflammatory response, cell division, control of lipid metabolism, as signaling molecules, supply of energy and protecting the biological membranes structure and function. In this study, a general overview of fatty acids, their biosynthesis mechanisms as well as biological importance have been discussed. Fatty acid desaturase enzymes and their sources have also been reviewed. Recent studies on the functional expression of different types of desaturase enzymes have also been discussed.
PDF Abstract XML References Citation
Received: December 07, 2016;
Accepted: February 06, 2017;
Published: May 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Lawal Garba, Mohd Shukuri Mohamad Ali, Siti Nurbaya Oslan and Raja Noor Zaliha Raja Binti Abdul Rahman, 2017. Review on Fatty Acid Desaturases and their Roles in Temperature Acclimatisation. Journal of Applied Sciences, 17: 282-295.
DOI: 10.3923/jas.2017.282.295
URL: https://scialert.net/abstract/?doi=jas.2017.282.295
DOI: 10.3923/jas.2017.282.295
URL: https://scialert.net/abstract/?doi=jas.2017.282.295
INTRODUCTION
Structurally, carbohydrates and proteins are made up of polymers (long chains) of polysaccharides (sugars) and polypeptides (amino acids) residues, respectively. However, lipids have a wide range of compounds with tremendous differences in structure. As lipids lack building blocks, fatty acids form the major components of their various classes including glycerides (phosphoglycerides and acylglycerols) and sphingolipids. Complex lipids are made up of fatty acids esterified to amino or alcohol groups and represent the predominant classes of lipids with fatty acids as their bulk constituents1,2.
Fatty acids are organic compounds containing a carboxylic acid group mostly at the end of an aliphatic chain. The chain may contain four and above carbon atoms, which are usually even numbered up to 24 and in some cases odd-numbered longer carbon atom chains are usually found (Fig. 1). Fatty acids may be saturated or unsaturated. The saturated fatty acids have a single bond in their aliphatic chains whereas the unsaturated ones have one or more double bonds in their chains. Most fatty acids are involved in key biological roles such as protecting the biological membranes structure and functions and supply of energy. Certain fatty acids and their derivatives known as polyunsaturated fatty acids (PUFA) are very crucial and perform several functions including inflammatory response, cell division, controlling lipid metabolism and serve as signaling molecules1-3.
Fatty acids nomenclature: Historically, many systems have been adopted to name fatty acids. However, the most frequently used and internationally recognised nomenclature system follows certain guidelines by the International Union of Pure and Applied Chemistry (IUPAC) in the Compendium of Chemical Terminology1. According to n-x , otherwise known as an omega system of nomenclature, C: Dn-x is used as general formula to describe fatty acids. The C stands for chain length, D is the number of double bonds and n-x (ωx) shows position of the first double bond located at the methyl end of the chain. Hence, 18:0 indicates a saturated fatty acid with an aliphatic chain of 18-carbon atoms. A monounsaturated fatty acid carrying a double bond at C9 position of its 18-carbon aliphatic chain can be presented as 18:1n-9 (18:1 ω9). The PUFA can be any fatty acid with two and above double bonds in their aliphatic chain such as 20:5n-3 (20:5 ω3), which is a fatty acid containing 20-carbon atoms with five double bonds within its aliphatic chain (Fig. 1)1.
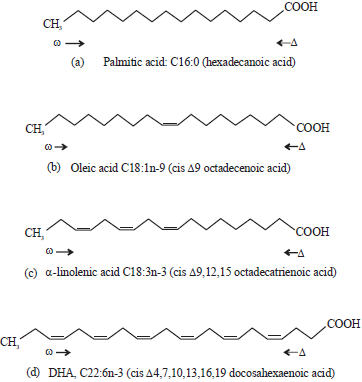 | |
| Fig. 1(a-d): | Examples of fatty acid nomenclature: (a) Saturated palmitic acid or hexadecanoic acid (16:0), (b) Monounsaturated fatty acid [oleic acid, cis Δ9-octadecenoic acid (18:1n-9)], (c) Polyunsaturated fatty acid [α-linolenic acid, cis Δ9,12,15-octadecatrienoic acid (18:3n-3)] and (d) Long-chain polyunsaturated fatty acid [docosahexaenoic acid, cis Δ4,7,10,13,16,19-docosahexaenoic acid (22:6n-3)]1 |
However, most fatty acid desaturase enzymes activities are indicated using the Δx (delta-x) system of nomenclature. Therefore, this system would be adopted to designate desaturase enzymes preferences for activities in this review. The system donates numbers of double bond counting from the carboxylic ends such as in 20:5Δ5,8,11,14,17, which can be presented as 20:5n-3. A fatty acid desaturase that produces a double bond at the C5 position of an aliphatic chain has Δ5 activity. Moreover, trivial names are also used to describe fatty acids, mostly indicating their key sources like oleic acid (18:1n-9) and palmitic acid (16:0) from olive and palm oils, respectively. Other useful system of naming fatty acids is referred to as semi-systematic, which names fatty acid by indicating the number of carbon atoms and their double bonds such as docosahexaenoic acid (DHA, 22:6n-3) and eicosapentaenoic acid (EPA, 20:5n-3)1.
Biosynthesis of fatty acids: Plants and animals owned the same lipogenic pathways of fatty acid biosynthesis. The first pathway is catalysed by two cytosolic enzyme systems, namely multienzyme Fatty Acid Synthase (FAS) complex and acetyl-CoA carboxylase. The FAS complex uses acetyl-CoA as source of carbon to synthesis saturated fatty acids essentially the 16:0 and 18:0 in animals and plants, respectively4. The FAS is broadly divided into two classes of type I systems (found mainly in animals and yeast, use a single large multifunctional polypeptide) and type II systems (found mostly in prokaryotes and plants, use a series of distinct monofunctional enzymes). The FAS mechanism has a series of decarboxylative Claisen condensation reactions in which condensation of malonyl-CoA produced by acetyl-CoA carboxylase from the growing acyl chain marks the key step. After every round of elongation, β-keto group is finally reduced to a fully saturated carbon chain via a chronological action by ketoacyl reductase, dehydrase and enoyl reductase activities. Acyl Carrier Protein (ACP) domain binds the growing acyl chain between its active sites by means of a covalent linkage to a phosphopantetheine prosthetic group and finally released it as a 16:0 by the action of a thioesterase. Primarily, acetyl-CoA is made in the mitochondria from two sources, protein and carbohydrate through oxidative catabolism of some amino acids and oxidative decarboxylation of pyruvate, respectively. This may lead to a citrate production (via the tri-carboxylic acid cycle) which is exported to the cytosol before a cytosolic acetyl-CoA is produced by the action of ATP citrate lyase. The NADPH is a reducing equivalent produced by the enzymes of carbohydrate metabolism including malic enzyme, tricarboxylic acid cycle (NADP-dependent isocitrate dehydrogenase and pentose phosphate pathway (glucose-6-phosphate dehydrogenase and 6-phosphogluconate dehydrogenase)1.
Elongation of fatty acid chains: Although C16:0 and C18:0 are the major fatty acids produced by FAS, other fatty acids of different chain lengths ranging from C20-C24 are very common however, carbon lengths greater than C24 are mostly found in eukaryotic organisms1,5. Fatty acid elongation is very crucial, serving as an alternative pathway of fatty acid production. Depending on the organism, many terms like Fatty Acid Chain Elongation System (FACES), elongase system and elongase have been used to explain enzymes capable of fatty acid elongation through the addition of two carbon atoms at the carboxyl end of the chain. In higher plants, fatty acid elongation is catalysed by an enzyme encoded by fatty acid elongase 1 (FAE1 gene) where only saturated and monounsaturated fatty acids are produced1,6. However, functional characterization of PUFA Fae1 has been reported from a marine parasitic protozoon Perkinsus marinus. A similar functional elongase enzyme that elongates PUFA has been demonstrated as IgASE1 from marine microalgae like Isochrysis galbana. More reports have confirmed that an active elongase produces PUFA from thraustochytrids1,7,8. Certain yeast species such as Saccharomyces cerevisiae and Mortierella alpina have been shown to produce different elongase enzymes. Saccharomyces cerevisiae produces mainly saturated and monounsaturated fatty acids has ELO1, ELO2 and ELO3 family of elongases. They differ in fatty acid chain length specificities1,9. Similarly, M. alpina synthesises arachidonic acid (ARA, 20:4n6) has two elongases comprising of GLELO and MAELO. The former enzyme elongates 18:3n-6 to 20:3n-6, perhaps the rate-limiting step in the biosynthesis of ARA whereas the latter has a preference for saturated and monounsaturated fatty acids elongation1,10.
However, plants and animals use four consecutive elongation reactions to produce fatty acids with chain lengths greater than C18 by simply adding 2-carbon unit taking place in the Endoplasmic Reticulum (ER) (Fig. 2). The reactions are similar to those involved in de novo fatty acid synthesis, particularly involving condensation, 1st reduction, dehydration and 2nd reduction. Four membrane-bound enzymes including very long fatty acids (Elovl) elongation proteins or condensing enzyme, β-ketoacyl-CoA reductase (Kar; 1st reduction), β-hydroxyacyl-CoA dehydrase (Hadc, dehydration) and trans-2-enoyl-CoA reductase (Ter, 2nd reduction) are known to catalyse these reactions (Fig. 2)1,11.
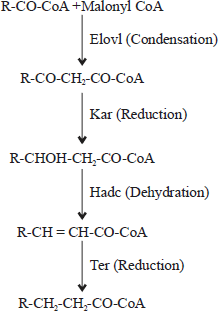 | |
| Fig. 2: | Pathway of fatty acid elongation in endoplasmic reticulum. The Elovl (elongase) catalyses condensation of malonyl-CoA that produces an activated fatty acid considered as the rate-limiting step of the pathway. Other enzymes associated with the pathway include 3-ketoacyl-CoA reductase (Kar), 3-hydroxyacyl-CoA dehydrase (Hadc) and trans-2, 3,-enoyl-CoA reductase (Ter)1 |
Previous studies have indicated that the condensing enzyme, Elovl belonging to elongation system was a rate-limiting. The enzyme, whose activity was regulated by the hormone and nutritional status was found to regulate certain elongation reactions based on the degree of unsaturation and chain length. However, other reports on cloning and functional characterization of a condensing enzyme revealed that the enzyme does not have any fatty acid specificity1,9. The ELOVL enzymes family has at least seven members as ELOVL1-7, further sub-divided into saturated and monounsaturated fatty acids elongases (ELOVL1, ELOVL3, ELOVL6 and ELOVL7) and elongases of PUFA designated as ELOVL2, ELOVL4 and ELOVL59,12.
Unsaturated fatty acids biosynthesis: Unsaturated fatty acids are carboxylic compounds having one or more double bonds within their acyl chains. The unsaturated fatty acids that contain only one double bond are called monounsaturated fatty acids whereas those with two and above double bonds are referred to as polyunsaturated fatty acids13,14. Generally, unsaturated fatty acids are synthesised through two major pathways, namely anaerobic and aerobic. The anaerobic pathway is performed by many bacterial species to create double bonds within the saturated acyl chains already produced at certain stage of fatty acids synthesis. A quite number of marine bacteria such as Shewanella sp. are well known EPA producers. Amino acid sequences analysis has revealed a strong relationship between the genes coding for EPA and complexes of microbial polyketide synthases (PKS) and FAS1,15,16. Such analysis was also performed on DHA-producing organisms including Moritella marina strain MP-1 and a thraustochytrid marine protist, Schizochytrium1.
The anaerobic pathway of unsaturated fatty acids synthesis has been well demonstrated in Escherichia coli where a dehyratase enzyme catalyses the removal of two hydrogen molecules with subsequent introduction of double bonds into saturated substrates before further elongation through the normal fatty acid biosynthesis machinery. The β-hydroxydecanoyl-ACP dehydrase (FabA) is a dehydratase enzyme, which catalyses the dehydration of β-hydroxydecanoyl-ACP, a saturated 10-carbon atoms intermediate to produce trans-2-decenoyl and water. In the reaction, a double bond is created at C7 and C8 position from the methyl end. The double bond may be hydrolysed by β-ketoacyl-ACP synthase (FabB) to produce saturated fatty acid through the typical fatty acid synthetic pathway. On the other hand, an isomerase enzyme (FabA) may be involved in isomerising the intermediate to cis-3-decenoyl, which is elongated by the β-ketoacyl-ACP synthase (FabB) to produce Palmitoleoyl-ACP (16:1ω7) and cis-vaccenoyl-ACP (18:1ω7)17,18 as shown in Fig. 3.
The aerobic pathway of unsaturated fatty acids synthesis is catalysed by the aerobic fatty acid desaturase enzymes in which full-length saturated substrates already synthesized either de novo from the anaerobic pathway, regioselective manner or obtained from the diets. This mechanism has been reported from virtually all higher organisms and prokaryotes. The desaturase enzymes utilized molecular oxygen, iron cofactors and reducing equivalents usually delivered by NADPH in two forms of electron transport systems depending on the cellular localization of the enzymes. Desaturase enzymes localized in the endoplasmic reticulum of plants, fungi and animals (acyl-CoA and acyl-lipid desaturases) derived the reducing equivalents from cytochrome b5 whereas acyl-lipid desaturases localised in the plants plastids and cyanobacteria obtained the reducing equivalents from ferredoxin (Fig. 4)19.
Biological importance of unsaturated fatty acids: Unsaturated fatty acids constitute the major components of membrane lipids of both prokaryotes and eukaryotic organisms.
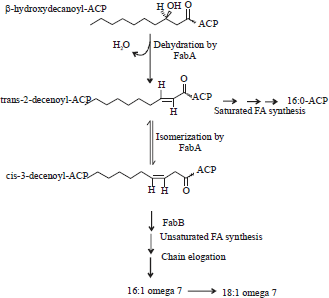 | |
| Fig. 3: | Anaerobic pathway of unsaturated fatty acid synthesis in Escherichia coli. FabA enzyme catalysed the removal of one molecule of water from β-hydroxydecanoyl-ACP intermediate to produce trans-2-decenoyl-ACP. Subsequently, critical step isomerisation reaction of unsaturated fatty acid synthesis produced cis-3-decenoyl-ACP. Elongation of cis-3-decenoyl-ACP catalysed by FabB produced two unsaturated fatty acids, namely palmitoleoyl-ACP (16:1 Δ9) and cis-vaccenoyl-ACP (18:1 Δ11). However, when trans-2-decenoyl-ACP is involved in the normal fatty acid synthesis catalysed by FabB enzyme, a saturated fatty acid is finally produced19 |
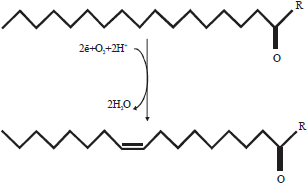 | |
| Fig. 4: | Desaturation reaction catalysed by the aerobic desaturase enzyme. The enzyme inserted a cis-double bond within saturated fatty acyl chain in the presence of 2e and O2 molecule. R can stand for CoA and phospholipid for acyl-CoA and acyl lipid desaturases, respectively20 |
They are primarily involved in regulating membrane lipids fluidity as a critical adaptive mechanism of psychrophilic organisms and some cold-blooded animals like reptiles to cold environments20,21. They also increase torpor bouts during hibernation in mammals, serve as signalling molecules in cell differentiation and DNA replication20,22.
The unsaturated fatty acids are increasingly gaining more recognition in the treatment of many diseases including autoimmune diseases, cancer, cardiovascular, heart, neurological and inflammatory disorders23-25. Therefore, the applications of long chain unsaturated fatty acids have been employed in health care system, food or nutraceutical industries24. For example, the ω-3 PUFAs promote the delivery of some drugs used in cancer treatment, provides protection against colorectal and breast cancer diseases26-29. Moreover, they have proven effectiveness in the management of different disorders like brain malfunction at old age, bone metabolism, joint pains or swelling and as antibacterial and anti-inflammatory agents28,30-32.
Dietary α-linolenic acid (ALA) is an unsaturated fatty acid, making the important ingredient of docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) production through the action of aerobic desaturase and elongase enzymes13. However, low level consumption and subsequent conversion of ALA to DHA and EPA requires supplementation through diets in human13,33-35.
FATTY ACID DESATURASE ENZYMES
Fatty acid desaturases belong to a group of enzymes family that essentially function in double bonds insertion at specified positions of fatty acyl chains. They require molecular oxygen and two electrons to accomplish their desaturation role. They are very crucial to all living organisms, playing a key role in unsaturated fatty acids synthesis necessary for membrane lipids fluidity. They are highly specific for substrate chain length and the location of double bond insertion within the fatty acyl chain36,37.
Fatty acid desaturase enzymes have been broadly classified into two evolutionary unrelated groups of soluble acyl-acyl carrier proteins and membrane-bound desaturases. The soluble group of desaturases has been abundantly found in plants plastids whereas the membrane-bound class of desaturases has been isolated from animals, fungi and bacteria. The membrane-bound class is further subdivided into acyl-coenzyme A (acyl-CoA) and acyl-lipid desaturases. The former has been reported from higher animals and fungi is generally involved in desaturation reactions at Δ9 position. The latter is predominantly found in plants and bacteria, catalyses desaturation of fatty acids linked to phospholipids. Besides their role in protecting membrane structure and function, fatty acid desaturases are very useful enzymes in polyunsaturated fatty acids synthesis including arachidonic and eicosapentaenoic acids due particularly to their ability to further introduce the second double bond at a position after the pre-existing Δ9 of unsaturated substrates36,38. Other beneficial roles of fatty acid desaturases in animals may include synthesis of essential omega-3 fatty acids, precursor molecules for pheromones, defensive fatty acids and functional hydrocarbons36,39.
Certain groups of fatty acid desaturases including soluble Δ9-stearoyl-ACP desaturases of plants, microsomal stearoyl-CoA desaturase (OLE1) of yeast and stearoyl-CoA desaturase of animals are very crucial enzymes of monounsaturated fatty acids synthesis like 18:1n-9 and 16:1n-71,40. Significant progress has been achieved in the study of stearoyl-CoA desaturases of animals as recent reports showed a successful crystallization and x-ray analysis of two stearoyl-CoA desaturases of mammals41. This indicated that the activity of these enzymes is universal in all organisms1,42,43.
However, in some organisms such as plants and algae, chloroplast membrane-bound desaturases can catalyse further desaturation of monounsaturated fatty acids to an array of unsaturated fatty acids. For example, linoleic acid (LA; 18:2n-6) is synthesised from 18:1n-9, then α-linolenic acid (ALA; 18:3n-3)1,44. Production of ALA from LA is catalysed either by ω3 or Δ15-desaturases45. Moreover, production of PUFA can be catalysed by Δ12 desaturases in some organisms like yeast whereas in others like protozoans and fungi, novel desaturase enzymes that function as both Δ12 and Δ15 have been reported1,46.
Contrary to PUFA production by certain organisms like plants, fungi and related aquatic organisms, some organisms can produce PUFA de novo. Introduction of double bond at Δ12 position of 18:1n-9 to produce LA has been reported from the axenic tissues of some insects such as cricket and cockroach1. According to some reports, many genes for Δ12-desaturase have been successfully isolated and characterised from insect. Two putative desaturase genes named as fat1 and fat2 were demonstrated from a nematode, Caenorhabditis elegans1,47,48. The fat1 codes for a newly ω3 desaturase, which is capable of desaturating both C18 and C20 substrates at C3 position from the methyl end whereas the fat2 gene codes for a Δ12-desaturase is active on both C16 and C18 substrates1. Recent report showed that this enzyme may be an active Δ15-desaturase. However, higher animals are unable to synthesize PUFA de novo due to their inability to produce Δ12/ω6 and Δ15/ω3 desaturses1,49.
SOURCES OF DESATURASES
Acyl-acyl carrier protein desaturases: Acyl-acyl carrier protein (ACP) desaturases consist of soluble enzymes associated with plant plastidial stroma and found in certain bacterial species like Mycobacterium and Streptomyces. The amino acids, D/EXXH are deemed significant for binding di-iron complex in these enzymes50. The ACP desaturases specifically catalyse the introduction of double bond within saturated acyl chains bound to ACP, producing cis-monounsaturated fatty acids in higher plants51. The de novo biosynthesis of fatty acids in higher plants may be preceded by malonyl-ACP (16:0-ACP) production from acetyl-CoA usually catalysed by acetyl-CoA carboxylase, then followed by malonyl-ACP production through the action of transacylase. Subsequent elongation of malonyl-ACP produces either palmitoyl-ACP (16:0-ACP) or stearoyl-ACP (18-ACP) after repeated cycles of 8-9 condensation-reduction-dehydration-reduction by complex Fatty Acid Synthase (FAS)51,52. The main products of FAS complexes in the plant plastidial tissue are 16:0-ACP and 18:0-ACP, which collectively constitute the major source of fatty acids in such tissues. However, heavy desaturation reactions that occur at different cellular localizations in plants results into a very low proportion of saturated fatty acids. The first desaturation reaction catalysed by a soluble Δ9 acyl-acyl carrier desaturase on aliphatic chains of plants fatty acids occurs in the chloroplasts or plastid tissues. It may also take place in the endoplasmic reticulum derivatives of very long chain CoA53,54. Therefore, these reactions usually produce oleoyl-ACP (18:1Δ9-ACP) from 18:0-ACP. However, in some instances, other precursors like 16:0-ACP may be desaturated to palmitoleoyl-ACP (16:1Δ9-ACP)51.
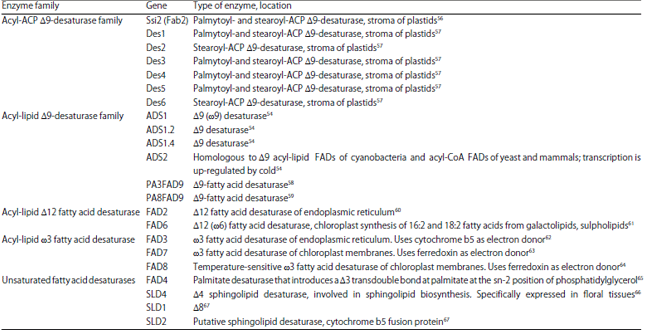
| Table 1: | Fatty acid desaturase enzymes and their encoding genes described from different organisms |
 | |
Three types of acyl-acyl carrier protein (ACP) desaturases described so far include Δ9-acyl-ACP, Δ6-acyl-ACP and Δ4-acyl-ACP desaturases. They differ in stereoselectivity and substrate specificity51. The Δ9-stearoyl-ACP desaturases are the most commonly reported members of soluble desaturases, isolated and characterized from various plant species such as soybean. However, acy-ACP of unusual specificities has been reported from Thunbergia alata (Blacked-eyed Susan vine) and Dolichandra unguis-cati L. (Cat’s claw)51,55.
Membrane-bound desaturases: Membrane-bound desaturases, which consist of acyl-lipid and acyl-CoA desaturases are the most abundant form of desaturase enzymes. The acyl-lipid desaturases specifically desaturate fatty acids attached to glycerolipids are commonly found in plants, cytoplasmic membranes of certain bacteria, thylakoid and plasmatic membranes of cyanobacteria whereas the acyl-CoA desaturases are predominantly found in fungi and animals, catalysing the desaturation of fatty acids related to endoplasmic reticulum. A group of three histidine-rich amino acids including ‘HXXXXH’, ‘HXXHH’ and ‘HXXHH’ has been very crucial as a presumptive active site of membrane desaturases50. Fatty acid desaturase enzymes of different stereoselectivity and cellular localisation reported from various organisms are summarised in Table 1.
Functional expression of soluble desaturases: The HaFAD7 is a gene coding for a plastid/chloroplast membrane omega-3 desaturase reported from sunflower. The gene codes for 443 amino acids protein with a predicted molecular mass of 50.8 kDa and hydrophobicity of four potential membrane-spanning regions. The RT-PCR analysis showed that the protein preferred expression in photosynthetically active tissues whereas heterologous expression in a unicellular cyanobacterium, Synechocystis sp., the PCC 6803 confirmed a functionally expressed omega-3 desaturase68. Arabidopsis thaliana was confirmed to have three genes coding omega-3 desaturases including FAD3, FAD7 and FAD8. However, expression of these genes was differently regulated as only FAD7 and FAD8 genes code for plastidial omega-3 desaturases. In another study, plastidial omega-3 desaturases with analogous pattern of expression was confirmed from Zea maize. Additionally, FAD7 and FAD8 genes expressions were induced by light and low temperature, respectively68-70.
A large number of genes coding stearoyl-ACP desaturases have been reported from various plants including Carthamus tinctorius, Ricinus communis, T. alata and Solanum tuberosum. Interestingly, the structure and functions of most stearoyl-ACP desaturases have been well demonstrated71-73. A coconut (Cocos nucifera L.) stearoyl-acyl carrier protein (ACP) desaturase (SAD) gene, CocoFAD was reported from the endosperm.
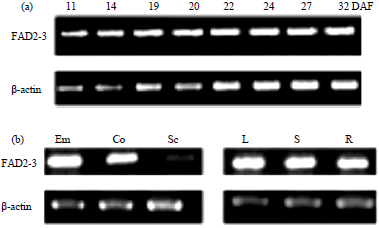 | |
| Fig. 5(a-b): | Profile of FAD2-3 expression in growing, (a) Soybean seeds and (b) Other tissues, spatial expression analysis embryo (Em), cotyledon (Co), seed coat (Sc), leaf (L), stem (S) and root (R)77 |
The gene was 1176 bp length, coding a 391 amino acids protein with more than 80% homology to certain plant Δ9-desaturase sequences. Real-time fluorescent quantitative PCR confirmed high yield of CocoFAD in the endosperm of 8-month-old coconut and leaf, reducing down to 50% in the older coconut of 15-month-old. Moreover, the gene was heterologously expressed in Saccharomyces cerevisiae and confirmed through GCMS analysis, which indicated a significant increase of palmitoleic acid (16:1) and oleic acid (18:1) showing that the coconut endosperm plastidial Δ9-desaturase was involved in fatty acid biosynthesis74.
Functional expression of membrane-bound desaturases: As membrane-bound desaturase enzymes are quite difficult to purify and crystallise, knowledge about their structure-function relationships is very limited75,76. Most of the previously reported membrane desaturases were studied through heterologous expression in suitable hosts followed by GCMS analysis confirmation of their in vivo activity. Oleate desaturase (FAD2) was isolated from plants Endoplasmic Reticulum (ER) and found to catalyse linoleic acid production. Soybean has three FAD2-like genes including FAD2-1A, FAD2-1B (restricted to seed) and FAD2-2 as a house keeping gene. Subsequently, FAD2-3 gene coding FAD2 isoform was reported. Heterologous expression of FAD2-3 gene in recombinant yeast cells has revealed a significant increase of linoleic acid (18:2) over the control cells indicating that FAD2-3 gene codes for a functional FAD2 protein. Moreover, semi-quantitative RT-PCR and insilico analysis further confirmed the constitutive expression of the FAD2-3 gene in both developing seeds and vegetative tissues77 (Fig. 5).
Physaria fendleri has many ORFs with tremendous similarity to oleate Δ12-desaturase of Arabidopsis. Out of 62, 59 clones were characterised as bifunctional oleate Δ12-hydroxylase/desaturase. Heterologous expression in yeast confirmed the successful isolation of full-length ORFs, designated as PfFAD2 from the remaining three clones and was the first report among all Physaria species encoding oleate Δ12-desaturases. The PfFAD2 gene was able to catalyse the production of 16:1 Δ9 and 18:1 Δ9 upon expression in both developing seed and leaf78.
Significant difference in the levels of seed α-linolenic acid (ALA) was observed among various linseeds. Ten varieties of Indian linseed plus one mutant variety were studied for fatty acids at different developmental stages. The varieties were grouped as high and low ALA content classes. Six microsomal desaturase genes, which act chronologically in fatty acid desaturation pathway were identified as SAD1, SAD2, FAD2, FAD2-2, FAD3A and FAD3B through real-time PCR. Pattern of gene specific, temporal and differential expression profiles of all the desaturase enzymes correlated well with differences in FA composition of the two groups. Additionally, the enzymes actively converted intermediate fatty acids like stearic, oleic and linoleic acids to ALA so much in high ALA class than the low ALA class. Sequence analysis of the six desaturase genes showed that difference in amino acid sequences has no influence on the disparity of ALA accumulation79. In another study, a PpSAD gene that codes for stearoyl-ACP desaturase (SAD) was isolated from the seeds of Pongamia pinnata, an oil producing plant. The gene has an open reading frame of 1182 bp coding 393 amino acids and a predicted molecular weight of 45.04 kDa. The Seeds of the P. pinnata were analysed for PpSAD gene expression through quantitative real-time PCR, which showed a clear gene expression at different seed developmental stages. Moreover, the expression analysis combined with previous studies suggests that stearoyl-ACP desaturase may take part in regulating the growth of plant seed and development73.
The ω3-fatty acid desaturase protein that catalyses n-3 polyunsaturated fatty acids (PUFAs) biosynthesis was reported from Mortierella alpina 1S-4. The protein is encoded by MAW3 gene, heterologously expressed in yeast and demonstrated de novo synthesis of two unusual fatty acids as n-4 hexadecadienoic acid (16:29cis,12cis) and n-1 hexadecatrienoic acid (16:39cis,12cis,15) confirmed by GCMS and 1H NMR analyses. Apart from the previously known ω3 activity of the protein on 18- and 20-carbon PUFAs, the enzyme introduced more double bonds at C12 and C15-positions of the endogenous palmitoleic acid (16:19cis) producing 16:2, 9cis,12cis and 16:3, 9cis,12cis,15, respectively confirming the protein as a bifunctional Δ12/Δ15-desaturase for 16-carbon fatty acids. This was the first study to demonstrate that fatty acid desaturase (MAW3) has Δ12-and Δ15-desaturation activity on 16-carbon fatty acid with preferred ω3-desaturation activity80.
Many bacteria have been studied for functional fatty acid desaturases of different specificities. One of the earliest reports involved identification and characterization of des gene which codes for Δ5-fatty acid desaturase in Bacillus subtilis. The Δ5-fatty acid desaturase has 352 amino acids with three conserved histidine and two membrane spanning domains common to most membrane-bound desaturases. Heterologous expression of the gene in E. coli revealed a functional Δ5-fatty acid desaturase which desaturated palmitic acid of the E. coli membrane phospholipids to produce a new monounsaturated fatty acid cis-5-hexadecenoic acid81.
Psychrobacter urativorans was found to produce high amount of monounsaturated fatty acids with unsaturation bond at C9 position. A novel gene, PuFAD9 was reported from the P. urativorans which has an ORF of 1,455 bp coding 484 amino acids. Functional expression of the gene in E. coli was confirmed by GCMS and indicated a functionally expressed protein capable of changing the ratio of palmitic to palmitoleic acid in the recombinant E. coli cells14. In a similar study, an active Δ9-fatty acid desaturase gene designated as PhFAD9 with an ORF of 1134 bp was isolated from Pseudoalteromonas sp. MLY15. The gene codes for 377 amino acids with a predicted molecular mass of 43.4 kDa. Functional analysis of E. coli cells transformed with the gene confirmed a functionally expressed membrane-bound Δ9-fatty acid desaturase which increased the E. coli cellular palmitoleic acids to about 91.7%. Moreover, the enzyme was able to desaturate stearic acid added into the E. coli growth medium to produce oleic acid82.
Recently, two novel genes designated as PA3FAD9 and PA8FAD9 were reported from Antarctic Pseudomonas sp. A3 and cold-tolerant Pseudomonas sp. A8, respectively. Each gene has an ORF of 1185 bp coding for a membrane-bound Δ9-fatty acid desaturase protein with 394 amino acids and a predicted molecular weight of 45 kDa. Functional expression in E. coli transetta (DE3) was confirmed by GCMS analysis, which showed active 9-fatty acid desaturase proteins that increased the overall cellular palmitoleic acid content of the recombinant E. coli. However, the E. coli expressing Δ9-fatty acid desaturase of Pseudomonas sp. A8 produced higher amount of palmitoleic acids (21%) more than the cells expressing Δ9-fatty acid desaturase of Pseudomonas sp. A3 (10.91 %). Moreover, both enzymes were able to desaturate exogenous stearic acid incorporated into the E. coli membrane phospholipid to produce 5% oleic acid further confirming functional Δ9-fatty acid desaturase enzymes with dual activity57,58.
ROLES OF FATTY ACID DESATURASES IN TEMPERATURE ACCLIMATISATION
As temperatures below the optimum requirements for organisms cause membrane lipids rigidity and abnormal cellular activities, poikilothermic organisms like bacteria, plants and fish respond to such environmental changes by remodelling the lipid composition of their membrane50. The amount of molecular disorder and molecular motion in a lipid bilayer is known as the membrane fluidity. Temperature stress tends to affect membrane fluidity of an organism. Cold temperature usually leads to a decreased membrane fluidity, which can be regained through desaturation of fatty acids already associated with the membrane lipids by fatty acid desaturases83. Compared to phospholipid containing high amount of saturated fatty acids, phospholipids rich in unsaturated fatty acids often have a much lower transition temperature. This is due particularly to the fact that saturated acyl chains are assembled tightly in the membrane lipids whereas the unsaturated fatty acids provide stearic hindrance due to a rigid kink of cis-double bonds that leads to a much poorer packing of the unsaturated fatty acids chain. Therefore, the optimum membrane lipids fluidity may regain its original state or somewhat close to it, restoring the normal cellular activity at lower temperature84.
The relationship between membrane fluidity and temperature changes has been well studied in many organisms including bacteria, cyanobacteria and fish83 through fluorescence polarization of 1,6-diphenyl-1,3,5 hexatriene (DPH).
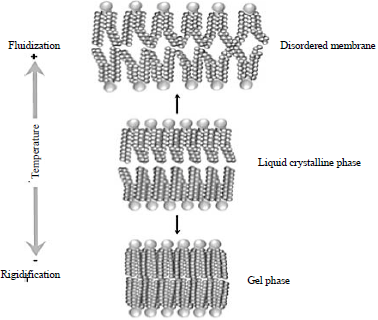 | |
| Fig. 6: | Changes in membrane structure and behavior of lipid bilayers at low and high temperatures. Low temperature increases lipid orderliness and membranes rigidity whereas high temperature decreases lipid orderliness but increases membranes fluidization86 |
The DPH was introduced parallel to the membranes lipids acyl chains. Weak depolarization was observed when the DPH stably interacts with rigidified membranes upon fluorescence but the depolarization increases with fluidized membranes. Heat shock increases membranes fluidity and may cause disintegration of lipid bilayer83,85 (Fig. 6).
CONCLUSION
Fatty acid desaturase enzymes introduce unsaturation bonds at specified positions of fatty acyl chains. The enzymes have been isolated from all groups of organisms and play significant role in regulating membrane fluidity. Class of soluble desaturases has been extensively studied. Membrane-bound class of desaturases lacks sufficient information on their structure-function relationships due largely to their difficult nature to purify. Membrane-bound desaturase of Mortierella alpine has been successfully expressed in a strain of Pichia pastoris and purified to some extent. However, detailed in vitro analysis and structural information of membrane desaturases are still in progress.
SIGNIFICANCE STATEMENT
Fatty acid desaturase enzymes are involved in the desaturation reactions that create double bonds within fatty acyl chains to produce unsaturated fatty acids. Unsaturated fatty acids have several beneficial applications in nutraceutical industries. The enzymes play a vital role in maintaining the right membrane fluidity. Soluble desaturases of plants have been studied in detail whereas, the membrane-bound desaturases of bacteria are poorly understood due largely to their difficult nature to purify. However, the only membrane-bound desaturases studied in detail so far were reported from rat and human.
ACKNOWLEDGMENT
The authors wish to acknowledge Putra grant, University Putra Malaysia for supporting this research (GP-IPS/2016/9471000).
REFERENCES
- Castro, L.F.C., D.R. Tocher and O. Monroig, 2016. Long-chain polyunsaturated fatty acid biosynthesis in chordates: Insights into the evolution of Fads and Elovl gene repertoire. Progr. Lipid Res., 62: 25-40.
CrossRefDirect Link - Schmitz, G. and J. Ecker, 2008. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipids Res., 47: 147-155.
CrossRefPubMedDirect Link - Garba, L., W. Latip, M.S.M. Ali, S.N. Oslan and R.N.Z.B.R. Abd Rahman, 2016. Unsaturated fatty acids in antarctic bacteria. Res. J. Microbiol., 11: 146-152.
CrossRefDirect Link - Wakil, S.J., J.K. Stoops and V.C. Joshi, 1983. Fatty acid synthesis and its regulation. Ann. Rev. Biochem., 52: 537-579.
CrossRefPubMedDirect Link - Poulos, A., 1995. Very long chain fatty acids in higher animals-a review. Lipids, 30: 1-14.
CrossRefDirect Link - Lemieux, B., M. Miquel and C. Somerville, 1990. Mutants of Arabidopsis with alterations in seed lipid fatty acid composition. Theor. Applied Genet., 80: 234-240.
CrossRefDirect Link - Harwood, J.L. and I.A. Guschina, 2009. The versatility of algae and their lipid metabolism. Biochimie, 91: 679-684.
CrossRefDirect Link - Venegas-Caleron, M., F. Beaudoin, O. Sayanova and J.A. Napier, 2007. Co-transcribed genes for long chain polyunsaturated fatty acid biosynthesis in the protozoon Perkinsus marinus include a plant-like FAE1 3-ketoacyl coenzyme A synthase. J. Biol. Chem., 282: 2996-3003.
CrossRefDirect Link - Jakobsson, A., R. Westerberg and A. Jacobsson, 2006. Fatty acid elongases in mammals: Their regulation and roles in metabolism. Prog. Lipid Res., 45: 237-249.
CrossRefDirect Link - Parker-Barnes, J.M., T. Das, E. Bobik, A.E. Leonard and J.M. Thurmond et al., 2000. Identification and characterization of an enzyme involved in the elongation of n-6 and n-3 polyunsaturated fatty acids. Proc. Natl. Acad. Sci., 97: 8284-8289.
CrossRefDirect Link - Guillou, H., D. Zadravec, P.G.P. Martin and A. Jacobsson, 2010. The key roles of elongases and desaturases in mammalian fatty acid metabolism: Insights from transgenic mice. Prog. Lipid Res., 49: 186-199.
CrossRefPubMedDirect Link - Naganuma, T., Y. Sato, T. Sassa, Y. Ohno and A. Kihara, 2011. Biochemical characterization of the very long-chain fatty acid elongase ELOVL7. FEBS Lett., 585: 3337-3341.
CrossRefDirect Link - Gupta, A., J. Vongsvivut, C.J. Barrow and M. Puri, 2012. Molecular identification of marine yeast and its spectroscopic analysis establishes unsaturated fatty acid accumulation. J. Biosci. Bioeng., 114: 411-417.
CrossRefDirect Link - Li, Y., M. Dietrich, R.D. Schmid, B. He, P. Ouyang and V.B. Urlacher, 2008. Identification and functional expression of a Δ9-fatty acid desaturase from Psychrobacter urativorans in Escherichia coli. Lipids, 43: 207-213.
CrossRefDirect Link - Allen, E.E. and D.H. Bartlett, 2002. Structure and regulation of the omega-3 polyunsaturated fatty acid synthase genes from the deep-sea bacterium Photobacterium profundum strain SS9. Microbiology, 148: 1903-1913.
Direct Link - Orikasa, Y., A. Yamada, R. Yu, Y. Ito and T. Nishida et al., 2004. Characterization of the eicosapentaenoic acid biosynthesis gene cluster from Shewanella sp. strain SCRC-2738. Cell. Mol. Biol. (Noisy-le-Grand), 50: 625-630.
PubMedDirect Link - Feng, Y. and J.E. Cronan, 2011. Complex binding of the FabR repressor of bacterial unsaturated fatty acid biosynthesis to its cognate promoters. Mol. Microbiol., 80: 195-218.
CrossRefDirect Link - Zhu, L., J. Cheng, B. Luo, S. Feng and J. Lin et al., 2009. Functions of the Clostridium acetobutylicium FabF and FabZ proteins in unsaturated fatty acid biosynthesis. BMC Microbiol., Vol. 9.
CrossRefDirect Link - Contreras, C., M. Franco, N.J. Place and R.F. Nespolo, 2014. The effects of poly-unsaturated fatty acids on the physiology of hibernation in a South American marsupial, Dromiciops gliroides. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., 177: 62-69.
CrossRefDirect Link - Hulbert, A.J., N. Turner, L.H. Storlien and P.L. Else, 2005. Dietary fats and membrane function: implications for metabolism and disease. Biol. Rev. Camb. Philos. Soc., 80: 155-169.
CrossRefDirect Link - Aguilar, P.S. and D. de Mendoza, 2006. Control of fatty acid desaturation: A mechanism conserved from bacteria to humans. Mol. Microbiol., 62: 1507-1514.
CrossRefDirect Link - Ruxton, C.H.S., S.C. Reed, M.J.A. Simpson and K.J. Millington, 2004. The health benefits of omega-3 polyunsaturated fatty acids: A review of the evidence. J. Hum. Nutr. Diet., 17: 449-459.
CrossRefPubMedDirect Link - Xing, H., X. Zhang, Q. Yang, R. Liu and Z. Bao et al., 2014. Separation of long chain fatty acids with different number of unsaturated bonds by fractional extraction: Experimental and COSMO-RS study. Food Chem., 143: 411-417.
CrossRefDirect Link - Yi, C., J. Shi, J. Kramer, S. Xue and Y.M. Jiang et al., 2009. Fatty acid composition and phenolic antioxidants of winemaking pomace powder. Food Chem., 114: 570-576.
CrossRefDirect Link - Chajes, V., G. Torres-Mejia, C. Biessy, C. Ortega-Olvera and A. Angeles-Llerenas et al., 2012. ω-3 and ω-6 polyunsaturated fatty acid intakes and the risk of breast cancer in Mexican women: Impact of obesity status. Cancer Epidemiol. Biomarkers Prev., 21: 319-326.
CrossRefDirect Link - De Lorgeril, M. and P. Salen, 2012. New insights into the health effects of dietary saturated and omega-6 and omega-3 polyunsaturated fatty acids. BMC Med., Vol. 10.
CrossRefDirect Link - Dillon, J.T., J.C. Aponte, R. Tarozo and Y. Huang, 2013. Purification of omega-3 polyunsaturated fatty acids from fish oil using silver-thiolate chromatographic material and high performance liquid chromatography. J. Chromatogr. A, 1312: 18-25.
CrossRefDirect Link - Patterson, R.E., S.W. Flatt, V.A. Newman, L. Natarajan and C.L. Rock et al., 2011. Marine fatty acid intake is associated with breast cancer prognosis. J. Nutr., 141: 201-206.
Direct Link - Guedes, A.C., H.M. Amaro and F.X. Malcata, 2011. Microalgae as sources of high added-value compounds-a brief review of recent work. Biotechnol. Prog., 27: 597-613.
CrossRefDirect Link - Huang, C.B. and J.L. Ebersole, 2010. A novel bioactivity of omega-3 polyunsaturated fatty acids and their ester derivatives. Mol. Oral Microbiol., 25: 75-80.
CrossRefDirect Link - Pereira, H., L. Barreira, F. Figueiredo, L. Custodio and C. Vizetto-Duarte et al., 2012. Polyunsaturated fatty acids of marine macroalgae: Potential for nutritional and pharmaceutical applications. Mar. Drugs, 10: 1920-1935.
CrossRefDirect Link - Anderson, B.M. and D.W. Ma, 2009. Are all n-3 polyunsaturated fatty acids created equal? Lipids Health Dis., Vol. 8.
CrossRefDirect Link - Burdge, G.C. and P.C. Calder, 2005. Conversion of α-linolenic acid to longer-chain polyunsaturated fatty acids in human adults. Reprod. Nutr. Dev., 45: 581-597.
CrossRefDirect Link - Innis, S.M., 2007. Fatty acids and early human development. Early Hum. Dev., 83: 761-766.
CrossRefDirect Link - Haritos, V.S., I. Horne, K. Damcevski, K. Glover and N. Gibb, 2014. Unexpected functional diversity in the fatty acid desaturases of the flour beetle Tribolium castaneum and identification of key residues determining activity. Insect Biochem. Mol. Biol., 51: 62-70.
CrossRefDirect Link - Shanklin, J. and E.B. Cahoon, 1998. Desaturation and related modifications of fatty acids. Ann. Rev. Plant Physiol. Plant Mol. Biol., 49: 611-641.
CrossRefDirect Link - Sperling, P., P. Ternes, T. Zank and E. Heinz, 2003. The evolution of desaturases. Prostaglandins Leukot. Essential Fatty Acids, 68: 73-95.
CrossRefPubMedDirect Link - Haritos, V.S., I. Horne, K. Damcevski, K. Glover, N. Gibb, S. Okada and M. Hamberg, 2012. The convergent evolution of defensive polyacetylenic fatty acid biosynthesis genes in soldier beetles. Nat. Commun., Vol. 3.
CrossRefDirect Link - Dobrzyn, A. and J.M. Ntambi, 2005. The role of stearoyl-CoA desaturase in the control of metabolism. Prostaglandins Leukot. Essential Fatty Acids, 73: 35-41.
CrossRefDirect Link - Bai, Y., J.G. McCoy, E.J. Levin, P. Sobrado, K.R. Rajashankar, B.G. Fox and M. Zhou, 2015. X-ray structure of a mammalian stearoyl-CoA desaturase. Nature, 524: 252-256.
CrossRefDirect Link - Castro, L.F.C., J.M. Wilson, O. Goncalves, S. Galante-Oliveira, E. Rocha and I. Cunha, 2011. The evolutionary history of the stearoyl-CoA desaturase gene family in vertebrates. BMC Evol. Biol., Vol. 11.
CrossRefDirect Link - Wang, H., M.G. Klein, H. Zou, W. Lane and G. Snell et al., 2015. Crystal structure of human stearoyl-coenzyme a desaturase in complex with substrate. Nat. Struct. Mol. Biol., 22: 581-585.
CrossRefDirect Link - Guschina, I.A. and J.L. Harwood, 2006. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res., 45: 160-186.
CrossRefPubMedDirect Link - Kotajima, T., Y. Shiraiwa and I. Suzuki, 2014. Functional screening of a novel Δ15 fatty acid desaturase from the coccolithophorid Emiliania huxleyi. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids, 1841: 1451-1458.
CrossRefDirect Link - Sayanova, O., R. Haslam, I. Guschina, D. Lloyd, W.W. Christie, J.L. Harwood and J.A. Napier, 2006. A bifunctional Δ12,Δ15-desaturase from Acanthamoeba castellanii directs the synthesis of highly unusual N-1 series unsaturated fatty acids. J. Biol. Chem., 281: 36533-36541.
CrossRefDirect Link - Vanhercke, T., P. Shrestha, A.G. Green and S.P. Singh, 2011. Mechanistic and structural insights into the regioselectivity of an acyl-CoA fatty acid desaturase via directed molecular evolution. J. Biol. Chem., 286: 12860-12869.
CrossRefDirect Link - Zhou, X.R., I. Horne, K. Damcevski, V. Haritos, A. Green and S. Singh, 2008. Isolation and functional characterization of two independently-evolved fatty acid Δ12-desaturase genes from insects. Insect Mol. Biol., 17: 667-676.
CrossRefDirect Link - Tocher, D.R., M.J. Leaver and P.A. Hodgson, 1998. Recent advances in the biochemistry and molecular biology of fatty acyl desaturases. Prog. Lipid Res., 37: 73-117.
CrossRefPubMedDirect Link - Rodriguez, M.F.R., A. Sanchez-Garcia, J.J. Salas, R. Garces and E. Martinez-Force, 2015. Characterization of soluble acyl-ACP desaturases from Camelina sativa, Macadamia tetraphylla and Dolichandra unguis-cati. J. Plant Physiol., 178: 35-42.
CrossRefDirect Link - Ohlrogge, J.B. and J.G. Jaworski, 1997. Regulation of fatty acid synthesis. Ann. Rev. Plant Physiol. Plant Mol. Biol., 48: 109-136.
CrossRefDirect Link - Ohlrogge, J., M. Pollard, X. Bao, M. Focke and T. Girke et al., 2000. Fatty acid synthesis: From CO2 to functional genomics. Biochem. Soc. Trans., 28: 567-574.
CrossRefDirect Link - Smith, M.A., M. Dauk, H. Ramadan, H. Yang and L.E. Seamons et al., 2013. Involvement of arabidopsis acyl-coenzyme a desaturase-like2 (At2g31360) in the biosynthesis of the very-long-chain monounsaturated fatty acid components of membrane lipids. Plant Physiol., 161: 81-96.
Direct Link - Zhang, P., J.W. Burton, R.G. Upchurch, E. Whittle, J. Shanklin and R.E. Dewey, 2008. Mutations in a Δ9-stearoyl-ACP-desaturase gene are associated with enhanced stearic acid levels in soybean seeds. Crop Sci., 48: 2305-2313.
CrossRefDirect Link - Kachroo, P., J. Shanklin, J. Shah, E.J. Whittle and D.F. Klessig, 2001. A fatty acid desaturase modulates the activation of defense signaling pathways in plants. Proc. Nat. Acad. Sci., 98: 9448-9453.
CrossRefDirect Link - Kachroo, A., J. Shanklin, E. Whittle, L. Lapchyk, D. Hildebrand and P. Kachroo, 2007. The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis. Plant Mol. Biol., 63: 257-271.
CrossRefDirect Link - Garba, L., M.S.M. Ali, S.N. Oslan and R.N.Z.R.A. Rahman, 2016. Heterologous expression of PA8FAD9 and functional characterization of a Δ9-fatty acid desaturase from a cold-tolerant Pseudomonas sp. A8. Mol. Biotechnol., 58: 718-728.
CrossRefDirect Link - Garba, L., M.S.M. Ali, S.N. Oslan and R.N.Z.R. Abd Rahman, 2016. Molecular cloning and functional expression of a Δ9-fatty acid desaturase from an antarctic Pseudomonas sp. A3. PLoS ONE Vol. 11, No. 8.
CrossRefDirect Link - Okuley, J., J. Lightner, K. Feldmann, N. Yadav and E. Lark, 1994. Arabidopsis FAD2 gene encodes the enzyme that is essential for polyunsaturated lipid synthesis. Plant Cell, 6: 147-158.
Direct Link - Falcone, D.L., S. Gibson, B. Lemieux and C. Somerville, 1994. Identification of a gene that complements an arabidopsis mutant deficient in chloroplast ω6 desaturase activity. Plant Physiol., 106: 1453-1459.
Direct Link - Arondel, V., B. Lemieux, I. Hwang, S. Gibson, H.M. Goodman and C.R. Somerville, 1992. Map-based cloning of a gene controlling omega-3 fatty acid desaturation in Arabidopsis. Science, 258: 1353-1355.
CrossRefPubMedDirect Link - Iba, K., S. Gibson, T. Nishiuchi, T. Fuse and M. Nishimura et al., 1993. A gene encoding a chloroplast omega-3 fatty acid desaturase complements alterations in fatty acid desaturation and chloroplast copy number of the fad7 mutant of Arabidopsis thaliana. J. Biol. Chem., 268: 24099-24105.
Direct Link - Gibson, S., V. Arondel, K. Iba and C. Somerville, 1994. Cloning of a temperature-regulated gene encoding a chloroplast [omega]-3 desaturase from Arabidopsis thaliana. Plant Physiol., 106: 1615-1621.
Direct Link - Gao, J., I. Ajjawi, A. Manoli, A. Sawin and C. Xu et al., 2009. Fatty acid desaturase4 of Arabidopsis encodes a protein distinct from characterized fatty acid desaturases. Plant J., 60: 832-839.
CrossRefDirect Link - Michaelson, L.V., S. Zauner, J.E. Markham, R.P. Haslam and R. Desikan et al., 2009. Functional characterization of a higher plant sphingolipid Δ4-desaturase: Defining the role of sphingosine and sphingosine-1-phosphate in Arabidopsis. Plant Physiol., 149: 487-498.
Direct Link - Ryan, P.R., Q. Liu, P. Sperling, B. Dong, S. Franke and E. Delhaize, 2007. A higher plant Δ8 sphingolipid desaturase with a preference for (Z)-isomer formation confers aluminum tolerance to yeast and plants. Plant Physiol., 144: 1968-1977.
Direct Link - Venegas-Caleron, M., A.M. Muro-Pastor, R. Garces and E. Martinez-Force, 2006. Functional characterization of a plastidial omega-3 desaturase from sunflower (Helianthus annuus) in cyanobacteria. Plant Physiol. Biochem., 44: 517-525.
CrossRefDirect Link - Berberich, T., M. Harada, K. Sugawara, H. Kodama, K. Iba and T. Kusano, 1998. Two maize genes encoding ω-3 fatty acid desaturase and their differential expression to temperature. Plant Mol. Biol., 36: 297-306.
CrossRefDirect Link - Horiguchi, G., H. Iwakawa, H. Kodama, N. Kawakami, M. Nishimura and K. Iba, 1996. Expression of a gene for plastid ω‐3 fatty acid desaturase and changes in lipid and fatty acid compositions in light‐and dark‐grown wheat leaves. Physiol. Plant., 96: 275-283.
CrossRefDirect Link - Davydov, R., B. Behrouzian, S. Smoukov, J. Stubbe, B.M. Hoffman and J. Shanklin, 2005. Effect of substrate on the diiron(III) site in stearoyl acyl carrier protein Δ9-desaturase as disclosed by cryoreduction electron paramagnetic resonance/electron nuclear double resonance spectroscopy. Biochemistry, 44: 1309-1315.
CrossRefDirect Link - Lindqvist, Y., W. Huang, G. Schneider and J. Shanklin, 1996. Crystal structure of delta9 stearoyl-acyl carrier protein desaturase from castor seed and its relationship to other di-iron proteins. EMBO J., 15: 4081-4092.
Direct Link - Ramesh, A.M., V. Kesari and L. Rangan, 2014. Characterization of a stearoyl-acyl carrier protein desaturase gene from potential biofuel plant, Pongamia pinnata L. Gene, 542: 113-121.
CrossRefDirect Link - Gao, L., R. Sun, Y. Liang, M. Zhang, Y. Zheng and D. Li, 2014. Cloning and functional expression of a cDNA encoding stearoyl-ACP Δ9-desaturase from the endosperm of coconut (Cocos nucifera L.). Gene, 549: 70-76.
CrossRefDirect Link - Li, S.F., L.Y. Song, G.J. Zhang, W.B. Yin, Y.H. Chen, R.R.C. Wang and Z.M. Hu, 2011. Newly identified essential amino acid residues affecting Δ8-sphingolipid desaturase activity revealed by site-directed mutagenesis. Biochem. Biophys. Res. Commun., 416: 165-171.
CrossRefDirect Link - Na-Ranong, S., K. Laoteng, P. Kittakoop, M. Tanticharoen and S. Cheevadhanarak, 2006. Targeted mutagenesis of a fatty acid Δ6-desaturase from Mucor rouxii: Role of amino acid residues adjacent to histidine-rich motif II. Biochem. Biophys. Res. Commun., 339: 1029-1034.
CrossRefDirect Link - Li, L., X. Wang, J. Gai and D. Yu, 2007. Molecular cloning and characterization of a novel microsomal oleate desaturase gene from soybean. J. Plant Physiol., 164: 1516-1526.
CrossRefDirect Link - Lozinsky, S., H. Yang, L. Forseille, G.R. Cook, I. Ramirez-Erosa and M.A. Smith, 2014. Characterization of an oleate 12-desaturase from Physaria fendleri and identification of 5'UTR introns in divergent FAD2 family genes. Plant Physiol. Biochem., 75: 114-122.
CrossRefDirect Link - Rajwade, A.V., N.Y. Kadoo, S.P. Borikar, A.M. Harsulkar, P.B. Ghorpade and V.S. Gupta, 2014. Differential transcriptional activity of SAD, FAD2 and FAD3 desaturase genes in developing seeds of linseed contributes to varietal variation in α-linolenic acid content. Phytochemistry, 98: 41-53.
CrossRefDirect Link - Kikukawa, H., E. Sakuradani, S. Kishino, S.B. Park and A. Ando et al., 2013. Characterization of a trifunctional fatty acid desaturase from oleaginous filamentous fungus Mortierella alpina 1S-4 using a yeast expression system. J. Biosci. Bioeng., 116: 672-676.
CrossRefDirect Link - Aguilar, P.S., J.E. Cronan Jr. and D. de Mendoza, 1998. A Bacillus subtilis gene induced by cold shock encodes a membrane phospholipid desaturase. J. Bacteriol., 180: 2194-2200.
Direct Link - Li, Y., X. Xu, M. Dietrich, V.B. Urlacher, R.D. Schmid, P. Ouyang and B. He, 2009. Identification and functional expression of a Δ9 fatty acid desaturase from the marine bacterium Pseudoalteromonas sp. MLY15. J. Mol. Catal. B: Enzymatic, 56: 96-101.
CrossRefDirect Link - Los, D.A., K.S. Mironov and S.I. Allakhverdiev, 2013. Regulatory role of membrane fluidity in gene expression and physiological functions. Photosynth. Res., 116: 489-509.
CrossRefDirect Link - Mansilla, M.C. and D. de Mendoza, 2005. The Bacillus subtilis desaturase: A model to understand phospholipid modification and temperature sensing. Arch. Microbiol., 183: 229-235.
CrossRefDirect Link - Horvath, I., A. Glatz, H. Nakamoto, M.L. Mishkind and T. Munnik et al., 2012. Heat shock response in photosynthetic organisms: Membrane and lipid connections. Progr. Lipid Res., 51: 208-220.
CrossRefDirect Link - Los, D.A. and N. Murata, 2004. Membrane fluidity and its roles in the perception of environmental signals. Biochimica et Biophysica Acta (BBA)-Biomembranes, 1666: 142-157.
CrossRefDirect Link