
Pankaj Sah
Applied Sciences Department, Applied Biology Section, Higher College of Technology (HCT), Al-Khuwair, P.O. Box 74, PC 133, Muscat,
Sultanate of Oman
Najwa Al-Hattali
Applied Sciences Department, Applied Biology Section, Higher College of Technology (HCT), Al-Khuwair, P.O. Box 74, PC 133, Muscat,
Sultanate of Oman
Hajer Al-Ajmi
Applied Sciences Department, Applied Biology Section, Higher College of Technology (HCT), Al-Khuwair, P.O. Box 74, PC 133, Muscat,
Sultanate of Oman
Iman Al-Bakari
Applied Sciences Department, Applied Biology Section, Higher College of Technology (HCT), Al-Khuwair, P.O. Box 74, PC 133, Muscat,
Sultanate of Oman
Journal of Applied Sciences
Year: 2016 | Volume: 16 | Issue: 7 | Page No.: 324-331







ABSTRACT
Background: Augmented anthropogenic activities are posing a direct threat to species diversity at regional and global scale for past many decades. Ecologists are very much alarmed about the serious repercussions of diversity-loss and predicting depauperate and poor functioning ecosystems in near future. However, the results of many such studies have been questioned too, on the basis of faulty inclusion of high productive species in experiments that rendered the relationship between diversity and ecosystem functioning debatable. Objective: Present study tries to find out the answer and deals exclusively with the effects of species diversity and richness on the productivity of plant communities in microcosms. Methodology: Pearson correlation and analysis of variance (ANOVA) carried out across all communities to observe the effect of species diversity and richness on herb productivity, indicated a highly significant and positive relationship (r = 0.85, F = 20.93, p<0.001), (r = 0.76, F = 11.23, p<0.01), respectively. Results: The results lend support to diversity-productivity hypothesis even at smaller scale ecosystems. This study comes to a new finding that at smaller scale ecosystems, the role of growth forms proves to be redundant and what matters most is species diversity and richness on the functioning of ecosystems. Conclusion: The present study accentuates the issues of ecological conservation and elucidates that more diverse and species rich areas are prerequisite for better functioning ecosystems. Therefore, this study recommends the conservation of biodiversity and that the productivity and functioning of an ecosystem can be enhanced by conserving and promoting its alpha diversity.
PDF Abstract XML References Citation
Received: April 28, 2016;
Accepted: May 15, 2016;
Published: June 15, 2016
How to cite this article
Pankaj Sah, Najwa Al-Hattali, Hajer Al-Ajmi and Iman Al-Bakari, 2016. Ecological Problem of Diversity-productivity Relationship
Elucidated Through Statistical Analysis of Experimental
Microcosmic Plant Communities. Journal of Applied Sciences, 16: 324-331.
DOI: 10.3923/jas.2016.324.331
URL: https://scialert.net/abstract/?doi=jas.2016.324.331
DOI: 10.3923/jas.2016.324.331
URL: https://scialert.net/abstract/?doi=jas.2016.324.331
INTRODUCTION
Augmented anthropogenic activities for past several decades have elevated concern for the existence of species and populations and its impact on ecosystems. These concerns have initiated a lot of observational and experimental studies on the relationship between species richness and ecosystem functioning and restoration ecology1-7. In many ecological studies, the impact of species richness on ecosystem functioning was investigated by comparing different ecosystem types varying in species numbers or alike ecosystems distributed at different geographical locations8,9. There are different parameters to study the functioning of an ecosystem, out of which productivity is considered as one of the excellent indicator of ecosystem functioning6,10. Topical studies which were performed in different ecosystems have shown that several communities and ecosystem processes are positively correlated with species diversity1,11,12. Yet many objections were also raised about the generality of these biodiversity effects13-15. As a result, it has been squabbled that the consequences of biodiversity loss are likely to be idiosyncratic, differing quantitatively and qualitatively between trophic groups and ecosystems16,17. Previous studies have shown that change in species equitability occurs much before does occur species extinction in ecosystem under anthropogenic influence. Therefore, it warrants increased and immediate attention18.
There are no exact available data about the current rates of species extinction. Yet some scientific estimates place it somewhere between two and three orders of magnitude higher than rates found in fossil records19,20. So, the concern over the effect of diversity-loss on ecosystem functioning is a hot topic in contemporary ecological studies.
In order to study the effects of community indices on ecosystem functioning and stability, experimental microcosms have been very successful. Many latest and significant developments in community ecology have been derived from experiments conducted in microcosms. Studies with microcosms have addressed a broad variety of phenomena, including climate change, biodiversity, assembly rules, habitat restoration, trophic dynamics and mycorrhizal associations. The common factor linking these studies are that they manipulate an individual environmental axis and explore the role that axis plays in structuring communities21. It has also been suggested that microcosms and mesocosms can be a useful approach for apparently intractable global problems, such as ecosystem responses to climate change or managing biodiversity through the design of nature reserve22.
The main objective of this study was to observe the effect of species diversity and richness on the functioning of experimental plant communities in microcosms. The study was designed in such a way that adjoining plots of a pair did not differ significantly in their immediate environmental conditions, whereas they differed significantly in their species richness and diversity values. Present experimental setup allowed us in correlating and understanding the relationship among diversity, richness, growth forms and microcosm functioning even with similar environmental conditions.
MATERIALS AND METHODS
Establishment of experimental microcosms: This study was conducted from May, 2011 to January, 2012 and a total of ten experimental plant communities with 300 seeded individuals in microcosms were raised in the greenhouse of Higher College of Technology, Muscat (Sultanate of Oman). The dimensions of microcosms were 36×22×5 cm3 and were almost same as designed and studied in a species richness and drought effect study in experimental plant communities23. Each microcosm was then filled with 500 g of commercial compost (Total N = 100 ppm, P2O5 = 185 ppm and K2O5 = 250 ppm). Furthermore, each microcosm was divided into two equal plots (18×22 = 396 cm2) by water impermeable barriers following23. In this way we maintained similar environmental conditions in all adjoining plots of pair in the studied microcosms (Fig. 1).
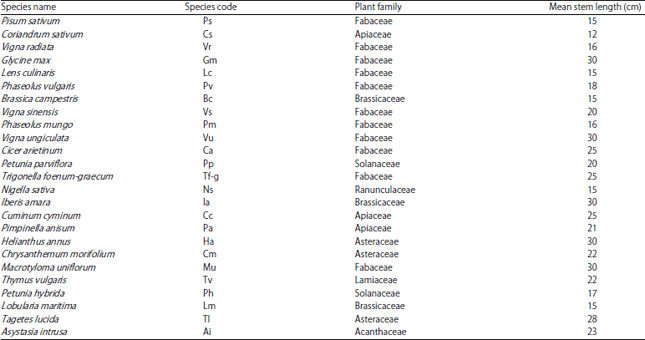
Species heterogeneity in plant communities (pairs of low and high species richness and diversities): For the establishment of plant communities, a total of 25 different herbs were used and chosen randomly for each plot. The herbs used for present study were of different mean stem length. Examples of studied herbs were Glycine max, Phaseolus vulgaris L., Vigna ungiculata (L). Walp., Cicer arientinum, Lens culinaris, Vigna radiata, Nigella sativa L., Cuminum cyminum, Pimpinella anisum L., Pisum sativum, Coriandrum sativum, Trigonella foenum-graecum, Macrotyloma uniflorum and Thymus vulgaris etc. (Table 1 and Fig. 1).
In each pair of microcosms, one plot was labeled as Low Species Richness (LSR) while other as High Species Richness (HSR). As a result, a total of 10 adjoining LSR and HSR plots were created in the experimental setup. Each plot was further divided into 30 smaller areas of 3.66×3.60 cm (13.2 cm2 area), having 1 seed/13.2 cm2. That’s how, this study could ensure that throughout all microcosms each individual was interacting with other in a defined area of available ecological resources.
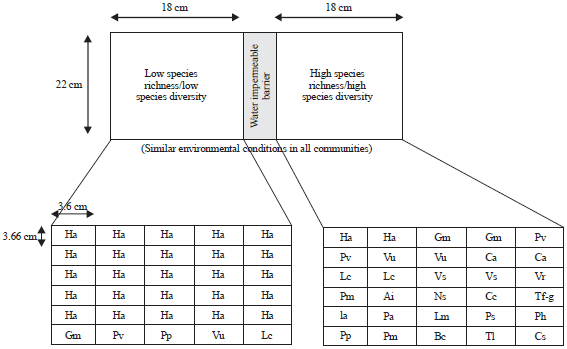 | |
| Fig. 1: | Schematic representation of experimental design showing two plant communities in a pair of plots. See the low species richness or diversity with tall stem class species i.e., Helianthus annus and high species richness or diversity with comparatively smaller stem class species. The diagram is just an example of two adjoining communities composition. Other communities were raised by various other species compositions as per the diversity gradient of present study |
| Table 1: | Description of herb species in experimental plant communities in microcosms |
 | |
In this way, 30 seeds of different species (30×13.2 = 396 cm2) were sown in each plot according to desired equitability or evenness to maintain adjoining low and high diverse ecosystems in each microcosmic pair (Table 2 and Fig. 1).
Growth form (stem length classes) and their significance: The stem lengths of each species was also recorded and distributed in 3 different growth form classes viz., short herbs (0-14 cm), medium herbs (15-20 cm) and tall herbs (21-30 cm).
| Table 2: | Paired t-test between High Species Richness (HSR) and adjoining Low Species Richness (LSR) communities in all 5 pairs of experimental microcosms |
 | |
The percent share of these growth forms was found to be 4, 40 and 56%, respectively among the species pool thus showing somewhat equal distribution between medium and tall herbs. The stem lengths of various herbs are of great significance in present microcosm study. In a recent study in experimental grassland, high community mean values of shoot length also contributed to high community biomass24. Since, it has been shown that the stem height or growth is positively related to aboveground biomass production25,26. This study took the stem length as an indicator of species biomass. The data were collected in order to observe the apparent effect of diversity, richness and growth forms on microcosm productivity. In some previous experimental studies, it was argued that higher diversity plots had more productive species which made it difficult to explain the effect of diversity on productivity. As according to a study of Huston27 in ECOTRON study1 the highly productive species were only included in the highest diversity treatment, rendering detection of a relationship between diversity and productivity inevitable. In the Cedar Creek diversity experiment28 the apparent diversity effect was because of greater probability of containing the most productive species in the high species richness treatment.
Species diversity and equitability: Diversity is the combination of two factors, species richness i.e., number of species and distribution among species, referred as equitability or evenness29. This study established ten different levels of diverse communities by manipulating the richness and equitability of different species per plot. Equitability or evenness was measured by equation30:
where, E is equitability or evenness index, H' is Shannon-Weiner diversity index and S is total species richness at the site. Species diversity of adjoining plant communities was calculated by Shannon-Weiner diversity index:
where, S is the total number of species or species richness, pi is relative abundance of each species and ln is natural log31. Each plot was irrigated at an interval of 48 h with an equal amount of distilled water (500 mL).
Estimation of aboveground community biomass: After 6 weeks of treatment, peak aboveground community biomass was calculated by harvest method, where clipping was done by hand with the help of sharp scissors. In order to obtain a valid estimation of the herbage, the vegetation was harvested very close to surface level32. The harvested plants were put in hot air oven at 80°C for 48 h to get biomass values6, which were expressed in terms of grams (g).
Statistical analysis of data: In order to understand the effect of community indices i.e., species diversity and richness on the functioning of microcosms, the data was statistically analyzed for Mean±Standard Error, linear regression, one way analysis of variance (ANOVA), Pearson bivariate correlation and paired t-test. All statistical analysis were performed using SigmaPlot (Systat Software, San Jose, California USA) and IBM SPSS Statistics for Windows, Version 20.0. Armonk, NY: IBM Corp.).
RESULTS AND DISCUSSION
Comparison between percent soil moisture and pH: Over the past few decades, accelerating rates of species extinction have prompted an increasing number of studies to reduce species diversity experimentally and examine how this alters the efficiency by which communities capture resources and convert those into biomass20,33,34. So, ecologically there is currently much interest in understanding how loss of biodiversity might alter ecological processes vital to the functioning of ecosystems35. As an alternative, ecologists have approached this problem by investigating how diversity influences stability and function within a multi-trophic setting in controlled microcosm experiments. The main advantage of microcosms is that they can easily be manipulated and replicated36. This study designed to keep the soil environment constant in each microcosm by using the commercial compost. This study also compared percent soil moisture and soil pH between adjoining plots of pairs in each microcosm before and at the end of experiment. For comparing the mean difference between abiotic components of high and low diverse plots of a pair we used paired t-test analysis. The results showed that percent soil moisture (t = -0.27, df = 4, α = 0.05, p = 0.40) and pH (t = 0.84, df = 4, α = 0.05, p = 0.22) did not differ significantly between high and low species richness plots across all microcosms. This was a good indicator to show that we were able to keep the abiotic environment constant among all studied plots of microcosms. In many earlier studies it was observed that soil nutrients were manipulated among various plant communities that increased variability in correlating diversity and ecosystem functioning37. Whereas, keeping the soil environment constant helped us to elucidate the apparent effect of community characteristics on ecosystem functioning.
Significant differences in species richness and diversity among all pairs of plots: Ecologists also found a strong effect of diversity on productivity and evidence suggestive of a simultaneous effect of composition and productivity4. The species composition (meaning the particular types and combinations of species present) has been reported to influence ecological processes to a much greater extent than the number of species27,38-42. The species richness between adjoining HSR and LSR plots was designed in such a way that all HSR plots (Mean = 15.2) had significantly greater species richness (t = 4.16, df = 4, α = 0.05, p<0.01) than their adjoining LSR plots (Mean = 4) (Table 2). However, the total numbers of individuals for each plot was kept constant at 30 individuals, this was done to observe the apparent effect of diversity and richness on productivity (Fig. 1). Shannon-Weiner index of species diversity between adjoining high and low species richness plots of all pairs showed highly significant differences. The high species diverse plots (Mean = 2.38) had significantly greater diversity (t = 7.85, df = 4, α = 0.05, p<0.001) than their adjoining low species diverse plots (Mean = 0.45). Species evenness or equitability values also exhibited similar trends, where high species even plots (Mean = 0.903) had significantly higher evenness values (t = 12.95, df = 4, α = 0.05, p<0.001) than their adjoining low species even plots (Mean = 0.326) (Table 2).
Relationship between species diversity and community productivity: In community ecological studies also, scientists previously thought species richness was mainly responsible for ecosystem functioning. However later on it was found that equitability and diversity were more influential than mere richness. Many previous studies on grassland plant communities12,28 and even experimental multi-trophic community1 were thought to show how productivity increases with species richness43. However, in later studies suggested what matters most is species diversity of functional groups, with species number per functional group being less important40. It has been suggested that due to human interference, abundance is disturbed first than richness i.e., diversity is disturbed first than richness, which ultimately will definitely affect ecosystem functioning. It was suggested that effects of diversity on productivity must come from interactions among individuals of different species3. This underlines the importance of evenness/equitability and abundance pattern among individuals, generating various combinations of species to species interactions, than mere species richness. A paired t-test analysis for biomass values showed that high species diverse community (Mean = 115.28 g) had significantly greater productivity (t = 4.41, df = 4, α = 0.05, p<0.01) than adjoining low diverse community (Mean = 60.87 g) (Table 2). Analysis of Variance (ANOVA), linear regression and correlation across all 10 adjoining plots showed that there is clear positive effect of species diversity on herb productivity, with a highly significant and positive relationship (r = 0.85, F = 20.93, α = 0.05, p<0.001) (Fig. 2).
Significant role of species richness and community productivity: In a heterogeneous habitat each species is a superior performer in only a part of site. As the heterogeneity or diversity increases, the magnitude of effect also increases, resulting in an increased coverage of the range of variations in the condition of habitat.
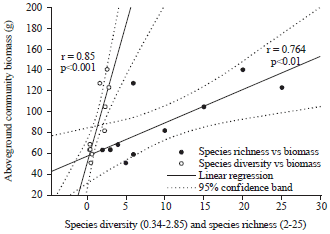 | |
| Fig. 2: | Linear regression analysis and pearson correlation show that both species diversity and richness are positively correlated with community biomass (r = 0.85, r = 0.76). The relationship is highly significant at 0.001 and 0.01 levels. The results are shown with 95% confidence band |
Increased diversity is expected to cause increased efficiency of resource capture and use, because chances increases for the presence of species that are better able to utilize existing conditions. Ecologists found that the significant difference in values of productivity in low species richness and high species richness plots suggests that there is the potential for increasing productivity of a site by realizing its potential α-diversity5. Increasing diversity leading to higher productivity may be attributed to ‘niche complementarity’ theory40,42. The niche complementarity theory39 predicts that differences among species in resource or environmental requirements would allow some combinations of species to more completely capture and use resources and thus have greater productivity than any individual species in monoculture, a phenomenon called over-yielding44. Studies investigating the relationship between species diversity and ecosystem functioning have advanced our basic understanding of community dynamics and may ultimately improve conservation by focusing attention on the processes critical to sustaining natural ecosystems41.
Species richness also had significant positive effect on community productivity in all plots. One way analysis of variance (ANOVA), linear regression and Pearson correlation between species richness and productivity showed that productivity tended to increase with increasing species richness across all 10 experimental plots, exhibiting a significantly positive relationship (r = 0.76, F = 11.23, α = 0.05, p<0.01) (Fig. 2).
Species diversity has greater role than species richness on productivity: In the present study, the results of experimental microcosms suggest that species diversity and richness both contribute significantly to microcosm productivity. However, species diversity (r = 0.85, p<0.001) has the lead over species richness (r = 0.76, p<0.01) in the functioning of terrestrial microcosms. Results presented in the current study are interesting as this suggests that at smaller levels such as of microcosms the role of growth forms seems to be redundant and what matters most is the diversity and equitability of species. This study found that in the experimental microcosms, percent share of tall herbs was significantly lower in high diversity plots than their adjoining low diversity plots, which reflects the apparent role of species diversity and richness on microcosm productivity. This can be explained by niche complementarity theory40,42 where more niche model supports more productivity by maximizing the usage of available resources. Similar findings were also reported by community ecologists even in natural ecosystems where a pair by pair survey of plots suggested that majority of HSR plots with higher biomass did not have a greater proportion of tall forbs5. It was concluded that when adjacent plots of natural communities are compared, the association between the presence of large and productive species of tall forbs and high species richness and diversity is weak5,6.
CONCLUSION
The findings of this microcosm study are important as it can be concluded that productivity of sites, even with similar environmental conditions can be increased by promoting and maintaining their alpha diversity. Although, both species diversity and richness contribute to microcosm functioning yet it is the diversity that ultimately regulates the functioning of microcosms and the growth forms may not have a larger role to play at the level of microcosms.
Therefore, this study recommends the need to conserve and maintenance of biological diversity for the better functional microcosms and ecosystems in a larger perspective. In conclusion, this study contributes to understand the unswerving effect of diversity, richness and growth forms on microcosm functioning and accentuates that encouragement must be given for the conservation of species in natural ecosystems.
ACKNOWLEDGMENTS
The authors are highly grateful to the Ministry of Manpower, Sultanate of Oman for providing necessary facilities and to the Dean, Heads of the Applied Science Department and Biology Section, Higher College of Technology, Muscat for their continuous support and encouragement. Authors are also thankful to all biology technicians for helping in laboratory and greenhouse arrangements.
REFERENCES
- Naeem, S., L.J. Thompson, S.P. Lawler, J.H. Lawton and R.M. Woodfin, 1994. Declining biodiversity can alter the performance of ecosystems. Nature, 368: 734-737.
CrossRefDirect Link - Tilman, D., 1996. Biodiversity: Population versus ecosystem stability. Ecology, 77: 350-363.
CrossRefDirect Link - Tilman, D., 1999. Diversity and production in european grasslands. Science, 286: 1099-1100.
CrossRefDirect Link - Hector, A., B. Schmid, C. Beierkuhnlein, M.C. Caldeira and M. Diemer et al., 1999. Plant diversity and productivity experiments in European grasslands. Science, 286: 1123-1127.
CrossRefPubMedDirect Link - Singh, S.P., P. Sah, V. Tyagi and B.S. Jina, 2005. Species diversity contributes to productivity-Evidence from natural grassland communities of the Himalaya. Curr. Sci., 89: 548-552.
Direct Link - Sah, P., K. Al-Badai and F. Al-Ajmi, 2016. A comparative study of in vitro seed germination ecology in two multipurpose leguminous tree species Albizia lebbeck (L.) Benth and Prosopis cineraria (L.) Druce under experimental saline stress. Ecol. Environ. Conserv., 22: 29-36.
Direct Link - Ricklefs, R. and D. Schluter, 1994. Species Diversity in Ecological Communities: Historical and Geographical Perspectives. 1st Edn., University of Chicago Press, Illinois, USA., ISBN-10: 0226718239.
Direct Link - Rosenzweig, M.L. and Z. Abramsky, 1994. How are Diversity and Productivity Related? In: Species Diversity in Ecological Communities: Historical and Geographical Perspectives, Rickleffs, R.E. and D. Schluter (Eds.). Chapter 5, University of Chicago Press, Chicago, IL., USA., ISBN-13: 9780226718231, pp: 52-64.
- Naeem, S., L.J. Thompson, S.P. Lawler, J.H. Lawton and R.M. Woodfin, 1995. Empirical evidence that declining species diversity may alter the performance of terrestrial ecosystems. Philos. Trans. R. Soc. London B: Biol. Sci., 347: 249-262.
CrossRefDirect Link - Tilman, D. and J.A. Downing, 1994. Biodiversity and stability in grasslands. Nature, 367: 363-365.
CrossRefDirect Link - Schwartz, M.W., C.A. Brigham, J.D. Hoeksema, K.G. Lyons, M.H. Mills and P.J. van Mantgem, 2000. Linking biodiversity to ecosystem function: Implications for conservation ecology. Oecogia, 122: 297-305.
Direct Link - Hooper, D.U., F.S. Chapin III, J.J. Ewel, A. Hector and P. Inchausti et al., 2005. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr., 75: 3-35.
CrossRefDirect Link - Srivastava, D.S. and M. Vellend, 2005. Biodiversity-ecosystem function research: Is it relevant to conservation? Annu. Rev. Ecol. Evol. Syst., 36: 267-294.
CrossRefDirect Link - Emmerson, M.C., M. Solan, C. Emes, D.M. Paterson and D. Raffaelli, 2001. Consistent patterns and the idiosyncratic effects of biodiversity in marine ecosystems. Nature, 411: 73-77.
CrossRefPubMedDirect Link - Covich, A.P., M.C. Austen, F. Barlocher, E. Chauvet and B.J. Cardinale et al., 2004. The role of biodiversity in the functioning of freshwater and marine benthic ecosystems. BioScience, 54: 767-775.
CrossRefDirect Link - Wilsey, B.J. and C. Potvin, 2000. Biodiversity and ecosystem functioning: Importance of species evenness in an old field. Ecol., 81: 887-892.
CrossRefDirect Link - Pimm, S.L., G.J. Russell, J.L. Gittleman and T.M. Brooks, 1995. The future of biodiversity. Science, 269: 347-350.
CrossRefPubMedDirect Link - Cardinale, B.J., D.S. Srivastava, J.E. Duffy, J.P. Wright, A.L. Downing, M. Sankaran and C. Jouseau, 2006. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature, 443: 989-992.
CrossRefDirect Link - Fraser, L.H. and P. Keddy, 1997. The role of experimental microcosms in ecological research. Trends Ecol. Evol., 12: 478-481.
CrossRefDirect Link - Benton, T.G., M. Solan, J.M.J. Travis and S.M. Sait, 2007. Microcosm experiments can inform global ecological problems. Trends Ecol. Evol., 22: 516-521.
CrossRefDirect Link - Nakamura, N., 2008. Species richness and aggregation effects on the productivity of ruderal plant communities under drought perturbation. Biosci. Horizons, 1: 128-135.
CrossRefDirect Link - Roscher, C., J. Schumacher, M. Gubsch, A. Lipowsky and A. Weigelt et al., 2012. Using plant functional traits to explain diversity-productivity relationships. PLoS ONE, Vol. 7.
CrossRefDirect Link - Schumacher, J. and C. Roscher, 2009. Differential effects of functional traits on aboveground biomass in semi-natural grasslands. Oikos, 118: 1659-1668.
CrossRefDirect Link - Thompson, K., A.P. Askew, J.P. Grime, N.P. Dunnett and A.J. Willis, 2005. Biodiversity, ecosystem function and plant traits in mature and immature plant communities. Funct. Ecol., 19: 355-358.
Direct Link - Huston, M.A., 1997. Hidden treatments in ecological experiments: re-evaluating the ecosystem function of biodiversity. Oecologia, 110: 449-460.
CrossRefDirect Link - Tilman, D., D. Wedin and J. Knops, 1996. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature, 379: 718-720.
CrossRefDirect Link - Raturi, G.P., 2012. Forest community structure along an altitudinal gradient of district rudraprayag of Garhwal Himalaya, India. Ecologia, 2: 76-84.
CrossRefDirect Link - Pielou, E.C., 1969. An Introduction to Mathematical Ecology. 1st Edn., Wiley-Interscience, New York, USA, ISBN: 0-471-68918-1, Pages: 286.
Direct Link - Chapin, III F.S., B.H. Walker, R.J. Hobbs, D.U. Hooper, J.H. Lawton, O.E. Sala and D. Tilman, 1997. Biotic control over the functioning of ecosystems. Science, 277: 500-504.
CrossRefDirect Link - Loreau, M., S. Naeem, P. Inchausti, J. Bengtsson and J.P. Grime et al., 2001. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science, 294: 804-808.
CrossRefDirect Link - Cardinale, B.J., J.P. Wright, M.W. Cadotte, I.T. Carroll and A. Hector et al., 2007. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proc. Natl. Acad. Sci., 104: 18123-18128.
CrossRefPubMedDirect Link - Lawton, J.H., 1995. Ecological experiments with model systems. Science, 269: 328-331.
CrossRefPubMedDirect Link - Tilman, D., 1993. Species richness of experimental productivity gradients: How important is colonization limitation? Ecology, 74: 2179-2191.
CrossRefDirect Link - Aarssen, L.W., 1997. High productivity in grassland ecosystems: Effected by species diversity or productive species? Oikos, 80: 183-184.
CrossRefDirect Link - Hooper, D.U. and P.M. Vitousek, 1997. The effects of plant composition and diversity on ecosystem processes. Science, 277: 1302-1305.
CrossRefDirect Link - Tilman, D., J. Knops, D. Wedin, P. Reich, M. Ritchie and E. Siemann, 1997. The influence of functional diversity and composition on ecosystem processes. Science, 277: 1300-1302.
CrossRefDirect Link - Chapin, III F.S., O.E. Sala, I.C. Burke, J.P. Grime and D.U. Hooper et al., 1998. Ecosystem consequences of changing biodiversity. BioScience, 48: 45-52.
CrossRefDirect Link - Loreau, M., 1998. Biodiversity and ecosystem functioning: A mechanistic model. Proc. Natl. Acad. Sci., 95: 5632-5636.
PubMedDirect Link - Tilman, D., 1999. The ecological consequences of changes in biodiversity: A search for general principles. Ecology, 80: 1455-1474.
CrossRefDirect Link