
Yalda Modirrahmati
Department of Medical Laboratory Technology, Faculty of Health Sciences, Universiti Teknologi MARA (UiTM), Puncak Alam Campus, Puncak Alam, 42300 Selangor, Malaysia
Mohd Fahmi Mastuki
Department of Medical Laboratory Technology, Faculty of Health Sciences, Universiti Teknologi MARA (UiTM), Puncak Alam Campus, Puncak Alam, 42300 Selangor, Malaysia
Roslinah Mohamad Hussain
Department of Medical Laboratory Technology, Faculty of Health Sciences, Universiti Teknologi MARA (UiTM), Puncak Alam Campus, Puncak Alam, 42300 Selangor, Malaysia
Journal of Applied Sciences
Year: 2015 | Volume: 15 | Issue: 6 | Page No.: 874-883







ABSTRACT
Dissemination of S. aureus within the host is facilitated by its ability to survive neutrophils (PMNs) clearance mechanisms. Identifying agents from natural sources that can boost neutrophil killing ability is therefore beneficial. We investigated effects of P. betle ethanolic extract on Myeloperoxidase (MPO) and killing potential of PMNs. Staphylococcus aureus suppresses MPO expression in PMNs with corresponding loss of MPO activity and 19.23% protein reduction. Piper betle extract alleviated this repressor effect with threefold (3.106 mU mL-1 μg-1 protein) increase in specific MPO activity in 30 min and 13.44% increase in protein concentration PMNs treated with S. aureus together with extract increased activity even higher (3.629 mU mL-1 μg-1 protein) with a corresponding 19.23% increase in protein concentration. The RT-qPCR analyses confirmed that S. aureus down-regulates MPO causing reduced expression (0.053 fold), whereas, treating PMNs with S. aureus with extract increased MPO expression by 2.29 fold. Staphylococcus aureus showed 79.24% survival in PMNs, whereas, addition of extract enhanced neutrophil killing ability as reflected by a marked reduction in cell survival to 7.14%. Piper betle ethanolic extract overcomes the inhibitory effects of Staphylococcus aureus on myeloperoxidase, resulting in increased clearance of intracellular cells.
PDF Abstract XML References Citation
Received: March 02, 2015;
Accepted: May 06, 2015;
Published: May 23, 2015
How to cite this article
Yalda Modirrahmati, Mohd Fahmi Mastuki and Roslinah Mohamad Hussain, 2015. Alleviation of Inhibitory Effects of Staphylococcus aureus on MPO Expression and Myeloperoxidase Activity in Human Neutrophils in vitro by Piper betle Linn. Ethanolic Extract. Journal of Applied Sciences, 15: 874-883.
DOI: 10.3923/jas.2015.874.883
URL: https://scialert.net/abstract/?doi=jas.2015.874.883
DOI: 10.3923/jas.2015.874.883
URL: https://scialert.net/abstract/?doi=jas.2015.874.883
INTRODUCTION
The search for novel therapeutic agents with better efficacies to combat infections and tumors has led to numerous investigations involving plant products as alternative sources. We investigated the effect of ethanolic extract of P. betle on an important component in neutrophils (PMNs), namely Myeloperoxidase (MPO) which contributes to its killing ability. Staphylococcus aureus is a tenacious pathogen that causes various types of infections and is problematic due to its multidrug-resistance trait. It produces an armament of proteins including catalase and alkylhydroxyperoxidase, that protect it from oxidative damage (Kobayashi et al., 2010). When confronted with the hostile environment within phagosomes, genes encoding these proteins; amongst others, are upregulated to counteract the deleterious effects presented by the human host. In addition, S. aureus circumvents host innate immune system and studies show that some pathogenic strains not only overcome neutrophil (PMNs) elimination but also induce neutrophil destruction (Gresham et al., 2000). Further, S. aureus is capable of surviving within PMNs which facilitates its dissemination within the host, thus contributing to infection (Nauseef, 2007).
Polymorphonuclear neutrophils (PMNs) are essential white blood cells of the innate immune system which characterizes the initial protection of the human host against invading microorganisms (Segal, 2005). Following phagocytic events, neutrophils degranulate releasing microbicidal enzymes and undergo oxidative burst resulting in production of potent oxidizing Reactive Oxygen Species (ROS) that kill invading microorganisms (Dahlgren et al., 2007). Myeloperoxidase is a heterodimeric heme-containing enzyme that is found in abundance in azurophilic granules in PMNs and consists of 2 alpha chains (each 60 kDa) and 2 beta chains (each 15 kDa). It generates hypochlorous acid (HOCl) from H2O2 and chloride ion which is a strong antioxidant that plays an essential role in killing of phagocytosed bacteria (Klebanoff, 2005). Individuals with myeloperoxidase deficiency show increased susceptibility to infections (Kutter et al., 2000) and increased susceptibility to candidal infections in patients with diabetes (Aratani et al., 1999).
Piper betle leaves are consumed in many traditions for treatment of a wide range of ailments and diseases. Extracts from Piper betle Linn possesses various bioactivities including immunomodulatory, anti-inflammatory, antimicrobial and antioxidant properties (Kanjwani et al., 2008; Tan and Chan, 2014). Piper betle leaves are chewed in combination with lime and areca nut for the prevention of oral malodour and heated leaves are used to reduce cough and asthma (Ramji et al., 2002). Further, P. betle contains considerable amounts of polyphenols and terpenoids such as hydroxychavicol, allylpyrocatechol and eugenol (Rathee et al., 2006) which contribute towards its free radical scavenging capability and inhibition of lipid peroxidation (Bhattacharya et al., 2007). Crude ethanolic extract of P. betle shows strong antimicrobial activity against numerous tested pathogenic bacterial strains including Staphylococcus aureus and Escherichia coli (Chahal et al., 2011).
Myeloperoxidase deficient in neutrophils have retarded killing ability of S. aureus (Peterson, 2011) although the exact mechanism has not been elucidated. We investigated the effects of ethanolic P. betle leaf extract on MPO expression and myeloperoxidase activity and its consequence on the killing ability of neutrophils that were challenged with S. aureus. We show that P. betle ethanolic extract alleviates the suppressor effects of S. aureus on MPO in human neutrophils, resulting in increased myeloperoxidase activity and killing ability of PMNs, suggesting its ability to enhance clearance of phagocytosed bacteria.
MATERIALS AND METHODS
Preparation of ethanolic extract: Piper betle leaves were collected from University Agriculture Park, University Putra Malaysia and authenticated at the Forest Research Institute Malaysia (PID 521113-29). Leaves were washed with distilled water and oven dried overnight at 60°C and ground (100 g) in a blender and then soaked in ethanol (95% w/v) for seven days at room temperature. The mixture was filtered and the filtrate was completely dried using a rotary evaporator at reduced pressure (60°C). The residue was dissolved in 10% dimethyl sulfoxide (Merck, Germany) and stored (Pin et al., 2009). The MIC (5 mg mL-1) of ethanolic P. betle extract against S. aureus was previously determined (Hussain et al., 2013).
Characterization of major phytoconstituent by HPLC NMR: Allylpyrocatechol was previously identified as the major constituent in Piper betle ethanolic extract using analytical HPLC UV (Abdullah and Hussain, 2015) which has been recently confirmed using optimized HPLC NMR (in review).
Staphylococcus aureus inoculum preparation: Staphylococcus aureus (ATCC25923) was maintained in frozen culture and propagated on sheep blood agar for experimental use. Species confirmation was performed by gram stain and conventional catalase and coagulase tests. Inoculum standardization was performed in reference to a standard growth curve performed in BHI broth and cells harvested for each assay were standardized (1×108 cells mL-1) at exponential phase. No ethics approval was obtained for use of this control strain (ATCC collection) and this study did not involve any genetic manipulations. Staphylococcus aureus ATCC 25923 strain used in this study was a non-clinical isolate and non-virulent control strain but capable of evading PMN killing (Gresham et al., 2000).
Neutrophil isolation and cell viability: Ethics approval for collection and handling of human blood was obtained from the Main Ethics Committee, Universiti Teknologi MARA (Ref: 600-RMI (5/1/6) and the study was conducted in accordance with approved ethical standards. Neutrophils were pooled from blood freshly collected from ten healthy donors. Six milliliter of blood was added to equal volume of PBS and neutrophils were harvested by dual density Ficoll gradient centrifugation as described by Venaille et al. (1994). Histopaque 1077 (3 mL) was overlayed with equal volume of Histopaque 1119 and topped with 6 mL of diluted blood and centrifuged (700×g, 30 min) at room temperature. Cells from the interphase layer, containing enriched neutrophils, were harvested, washed and resuspended in cold PBS and counted. Neutrophil viability counts were performed as described by Strober (2001) using 0.4% trypan blue. Cells were counted after 3-5 min of mixing and percentage of viable cells was calculated. Cell morphology was confirmed using the Wright stain.
Treatment of neutrophils with S. aureus and P. betle ethanolic extract: For the assays, 1 mL of neutrophils (4.36×104) and 100 μL of the serum were added to each milliliter of unopsonized bacteria. Four sample tubes were prepared consisting of, neutrophils only (negative control), neutrophils with S. aureus (positive control), neutrophils with MIC (5 mg mL-1) P. betle ethanolic extract and neutrophils with S. aureus and 5 mg mL-1 P. betle ethanolic extract. Tubes were incubated for 1, 2 and 3 h (37°C) followed by centrifugation (4000 rpm, 5 min). One milliliter ice cold PBS was added to each tube, centrifuged at 100×g (5 min), supernatant discarded and pellets were washed twice with ice cold PBS. The pellet was resuspended in 1 mL PBS containing Triton X-100 and incubated at room temperature (5 min) to lyse neutrophils.
RT-qPCR analysis: Total RNA from each sample was isolated using RNeasy mini kit (Qiagen, USA) according to manufacturer’s instructions. The cDNA synthesis was performed using Quanti Tect Reverse Transcription Kit (Qiagen, USA) according to manufacturer’s instructions. The RNA purity and concentration were confirmed using biophotometer (Eppendorf, USA).
| Table 1: | Primers and probe designated targeting the MPO and housekeeping gene (HPRT1) of neutrophils |
 | |
Complete MPO sequence (human neutrophils) was accessed from NCBI Gen Bank database. For probe and primers, sequences were each aligned using Sci Tools Real Time PCR software (Integrated DNA Technologies, USA) to identify unique regions that are suitable for amplification of the MPO (Table 1). Specificity of primers and probes were confirmed using primer blast and Nucleotide blast tool (NCBI). The probe was labeled with 6-carboxy-fluorescein (FAM) at the 5’ end (reporter) and with the 6-carboxy-tetramethylrhodamine (TARMA) at the 3’ end (quencher). All the steps were repeated for the designated reference housekeeping gene (HPRT1) for selection of forward and reverse primers and probe (Table 1).
Reactions were performed in 20 μL final reaction volume containing 2X Quanti Nova Probe PCR Mastermix, respective cDNA, primers and probes in RNase-free water. Real-time PCR was performed (Biorad, USA) according to manufacturer’s recommendations (2 min at 95°C, 5 sec at 95°C and 5 sec at 60°C for 40 cycles). The PCR reaction for each gene was performed in triplicate and gene expressions were analyzed using the Livak method. Data generated in the bar graph were derived using the Livak or Ct method (2‾ΔΔCt) (Livak and Schmittgen, 2001) also known ΔΔCt as defined by the following equation:
2‾ΔΔCt = 2-[(Ct target (Treated sample)–Ct HKG)–(Ct target (Untreated sample)–Ct HKG)]
Bradford protein assay: Protein concentrations in lysates were determined using the Bradford assay by spectrophotometric analysis (595 nm) according to manufacturer’s instruction and referenced against a standard plotted curve (BSA) for total protein content.
TNB consumption and myeloperoxidase activity assay: Myeloperoxidase (MPO) activity assay was performed using the colorimetric NWLSSTM assay kit (Sigma Aldrich, Malaysia) according to manufacturer’s instructions (Northwest Life Science Specialties, 2014). The 5-thio-2-nitrobenzoic acid (TNB) was first used to develop a standard curve and ΔA412 of each lysate was compared to the standard curve to determine TNB consumption (nM) which was then used to calculate MPO activity in samples as follows:
![]()
where, B is amount (nM) of TNB consumed, V is Sample volume (mL) added to each well.
One unit MPO activity is defined as the amount of enzyme that catalyzes sufficient HOCL production resulting in formation of 1 nM TauNHCl (pH 6.5, 25°C) over 30 min in the presence of 100 mM chloride and 100 μM H2O2.
Specific TNB consumption and myeloperoxidase activity were calculated using the following equation (Philip, 2007):
![]()
Neutrophil killing assay: For the neutrophil killing assay, bacterial cells were first opsonized using Phosphate Buffered Saline (PBS) containing 10% serum (1:10) and incubated at 37oC (20 min). The assay was performed using two tubes; consisting of optimized S. aureus with PMN and S. aureus with P. betle ethanolic extract and PMNs, respectively. Tubes were incubated for 1 h (37°C) and samples were washed and centrifuged (100×g) twice in cold PBS to remove extracellular cells. The pellets were resuspended in lysing buffer (Triton-X100, SDS and NaCl), vortexed for 1 min and incubated for 15 min (37oC) to lyse neutrophils and release intracellular cells. Lysates were inoculated on BHI agar and incubated overnight (37oC). The number of viable intracellular bacteria in each lysate was determined by colony counts, in triplicate, after overnight (37oC) incubation.
RESULTS
It is uncertain how P. betle ethanolic extract affects interactions between organism and phagocytic cells. Therefore, we investigated its effects on human Myeloperoxidase (MPO), an important component in neutrophils that is involved in killing of phagocytosed bacteria. Neutrophils used in this study were freshly harvested for each test procedure and.
MPO expression: The effect of P. betle ethanolic extract on MPO expression in neutrophil was investigated using real-time PCR to determine changes in the targeted gene. The RT-qPCR analyses were performed in triplicate together with the housekeeping gene, HPRT1. Amplification efficiencies for both genes (MPO and HPRT1) were approximately 100%.
The effect of P. betle ethanolic extract on expression of MPO in neutrophil was investigated using real-time PCR to determine changes in the targeting gene. Expression of MPO was determined after treatment (in triplicate) with 5 mg mL-1 P. betle ethanolic extract for 1 h.
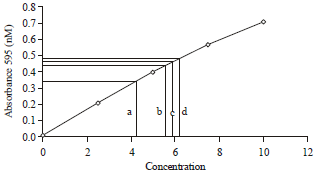 | |
| Fig. 1: | BSA standard protein curve showing total protein concentrations in lysate prepared from a: PMNs+S. aureus, b: PMNs, c: PMNs+P. betle and d: PMNs+P. betle+S. aureus |
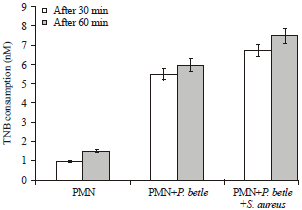 | |
| Fig. 2: | TNB consumption (nM) by each lysate after 30 and 60 min incubation |
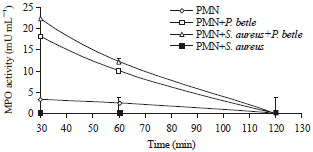 | |
| Fig. 3: | Myeloperoxidase activity assay (mU mL-1) |
The relative expression of MPO gene in treated and untreated samples with HPRT1 reference gene was determined using the 2-ΔΔCt (Livak method). RT-PCR analyses were performed in triplicate and the amplification efficiencies for both genes (MPO and HPRT1) were near 100%. According to the amplification graphs (Fig. 1-3), MPO in treated PMNs with P. betle extract amplified higher than untreated PMNs and treated PMNs with S. aureus with indicating a higher expression of the gene in treated PMNs with P. betle extract to untreated PMNs and treated PMNs with S. aureus.
First RT-PCR run: Analysis of obtained data from first RT-PCR run showed that S. aureus suppresses MPO expression in PMNs causing approximately nil (0.06 fold) expression compared to untreated PMNs (baseline).
| Table 2: | Quantification data of target gene (MPO) and reference gene (HPRT1) in treated and untreated samples of neutrophils for the first, second and third RT-PCR run |
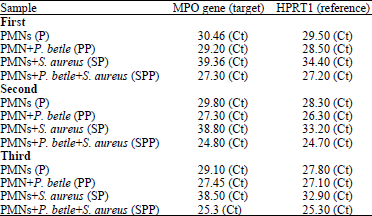 | |
| *Ct or threshold cycle is the number of cycles required for the fluorescent signal to cross the threshold | |
Increased MPO expression was observed in PMNs treated with P. betle ethanolic extract (1.20 fold) and interestingly, PMNs exposed to S. aureus showed higher MPO expression in presence of P. betle extract (1.82 fold) compared to baseline (untreated PMNs). Evaluated data showed that P. betle extract overcomes the inhibitory effect of S. aureus on MPO expression which elevated from 0.06 fold in PMNs treated with S. aureus to 1.82 fold in PMNs treated with both S. aureus and P. betle extract (Fig. 4a, Table 2).
Relative quantification of MPO expression using Livak method or ΔΔCt:
| • | PMNs+P. betle | |
| • | ΔCT (P) = Ct (target, P)-Ct (reference, P) = 30.46-29.50 = 0.96 | |
| • | ΔCT (PP) = Ct (target, PP)-Ct (reference, PP) = 29.20+28.50 = 0.7 | |
| • | ΔΔCt = ΔCt (PP)-ΔCt (P) = 0.7-0.96 = -0.26 | |
| • | Normalized expression ratio = 2‾ΔΔCT = 2‾(-0.26) = 1.20 | |
| • | PMNs treated with P. betle ethanolic extract expressed MPO at 1.20 fold higher than untreated PMNs | |
| • | PMNs+S. aureus | |
| • | ΔCT (P) = Ct (target, P)-Ct (reference, P) = 30.46- 29.50 = 0.96 | |
| • | ΔCT (SP) = Ct (target, SP)-Ct (reference, SP) = 39.36 -34.40 = 4.96 | |
| • | ΔΔCt = ΔCt (PP)-ΔCt (P) = 4.96-0.96 = 4 | |
| • | Normalized expression ratio = 2‾ΔΔCT = 2‾4 = 0.06 | |
| • | PMNs treated with S. aureus expressed MPO at 0.06 fold lower than untreated PMNs | |
| • | PMNs+P. betle+S. aureus | |
| • | ΔCT (P) = Ct (target, P)-Ct (reference, P) = 30.46- 29.50 = 0.96 | |
| • | ΔCT (SPP) = Ct (target, SPP)-Ct (reference, SPP) = 27.30-27.20 = 0.1 | |
| • | ΔΔCt = ΔCt (SPP)-ΔCt (P) = 0.1-0.96 = -0.86 | |
| • | Normalized expression ratio = 2‾ΔΔCT = 2‾(-0.86) = 1.82 | |
| • | PMNs treated with P. betle ethanolic extract and S. aureus expressed MPO at 1.82-fold higher than untreated PMNs | |
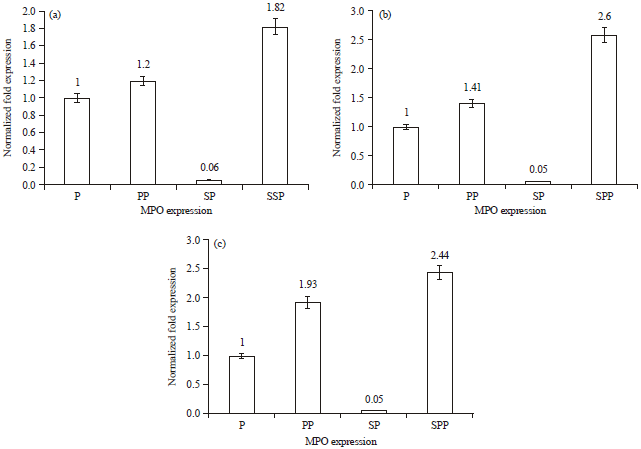 | |
| Fig. 4(a-c): | Gene fold expression of MPO in PMNs (P), PMNs+P. betle (PP), PMNs+S. aureus (SP) and PMNs+P. betle+S. aureus (SPP) for the, (a) First RT-PCR run, (b) Second RT-PCR run and (c) Third RT-PCR run |
Second RT-PCR run: Analysis of obtained data from first RT-PCR run showed that S. aureus suppresses MPO expression in PMNs causing approximately nil (0.05 fold) expression compared to untreated PMNs (baseline). Increased MPO expression was observed in PMNs treated with P. betle ethanolic extract (1.41 fold) and interestingly, PMNs exposed to S. aureus showed higher MPO expression in presence of P. betle extract (2.60 fold) compared to baseline (untreated PMNs). Evaluated data showed that P. betle extract overcomes the inhibitory effect of S. aureus on MPO expression which elevated from 0.05 fold in PMNs treated with S. aureus to 2.6 fold in PMNs treated with both S. aureus and P. betle extract (Fig. 4b, Table 2).
Relative quantification by using Livak method or ΔΔCt:
| • | PMNs+P. betle | |
| • | ΔCT (P) = Ct (target, P)-Ct (reference, P) = 29.80- 28.30 = 1.5 | |
| • | ΔCT (PP) = Ct (target, PP)-Ct (reference, PP) = 27.30 +26.30 = 1 | |
| • | ΔΔCt = ΔCt (PP)-ΔCt (P) = 1 -1.5 = -0.05 | |
| • | Normalized expression ratio = 2‾ΔΔCT = 2‾(-0.5) = 1.41 | |
| • | PMNs treated with P. betle ethanolic extract expressed MPO at 1.41 fold higher than untreated PMNs | |
| • | PMNs+S. aureus | |
| • | ΔCT (P) = Ct (target, P)-Ct (reference, P) = 29.80- 28.30 = 1.5 | |
| • | ΔCT (SP) = Ct (target, SP)-Ct (reference, SP) = 38.80 -33.20 = 5.6 | |
| • | ΔΔCt = ΔCt (SP)-ΔCt (P) = 5.6-1.5 = 4.1 | |
| • | Normalized expression ratio = 2‾ΔΔCT = 2‾4.1 = 0.05 | |
| • | PMNs treated with S. aureus expressed MPO at 0.05 fold lower than untreated PMNs | |
| • | PMNs+P. betle+S. aureus | |
| • | ΔCT (P) = Ct (target, P)-Ct (reference, P) = 29.80- 28.30 = 1.5 | |
| • | ΔCT (SPP) = Ct (target, SPP)-Ct (reference, SPP) = 24.80-24.70 = 0.1 | |
| • | ΔΔCt = ΔCt (SPP)-ΔCt (P) = 0.1-1.5 = -1.4 | |
| • | Normalized expression ratio = 2‾ΔΔCT = 2‾(-1.4) = 2.6 | |
| • | PMNs treated with P. betle ethanolic extract and S. aureus expressed MPO at 2.6 fold higher than untreated PMNs | |
Third RT-PCR run: Analysis of obtained data from first RT-PCR run showed that S. aureus suppresses MPO expression in PMNs causing approximately nil (0.05 fold) expression compared to untreated PMNs (baseline). Increased MPO expression was observed in PMNs treated with P. betle ethanolic extract (1.93 fold) and interestingly, PMNs exposed to S. aureus showed higher MPO expression in presence of P. betle extract (2.44 fold) compared to baseline (untreated PMNs).
| Table 3: | Mean normalized expression ratio data for the overall three RT-PCR runs of target gene MPO for untreated and treated PMNs with (5 mg mL-1) P. betle ethanolic extract and (1×108 cell mL-1) S. aureus. Ct or threshold cycle is the number of cycles required for the fluorescent signal to cross the threshold |
 | |
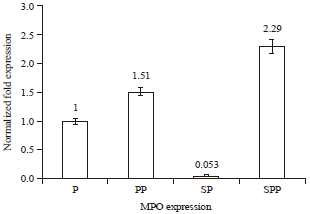 | |
| Fig. 5: | Averaged (three trials) MPO expression o in PMNs (P), PMNs+P. betle (PP), PMNs+S. aureus (SP) and PMNs+P. betle+S. aureus (SPP) |
Evaluated data showed that P. betle extract overcomes the inhibitory effect of S. aureus on MPO expression which elevated from 0.05 fold in PMNs treated with S. aureus to 2.44 fold in PMNs treated with both S. aureus and P. betle extract (Fig. 4c, Table 2).
Relative quantification by using Livak method or ΔΔCt:
| • | PMNs+P. betle | |
| • | ΔCT (P) = Ct (target, P)-Ct (reference, P) = 29.10- 27.80 = 1.30 | |
| • | ΔCT (PP) = Ct (target, PP)-Ct (reference, PP) = 27.45 -27.10 = 0.35 | |
| • | ΔΔCt = ΔCt (PP) – ΔCt (P) = 0.35-1.30 = -0.95 | |
| • | Normalized expression ratio = 2‾ΔΔCT = 2‾(-0.95) = 1.93 | |
| • | PMNs treated with P. betle ethanolic extract expressed MPO at 1.93 fold higher than untreated PMNs | |
| • | PMNs+S. aureus | |
| • | ΔCT (P) = Ct (target, P)-Ct (reference, P) = 29.10- 27.80 = 1.30 | |
| • | ΔCT (SP) = Ct (target, SP)-Ct (reference, SP) = 38.50 -32.90 = 5.6 | |
| • | ΔΔCt = ΔCt (SP)-ΔCt (P) = 5.6-1.30 = 4.3 | |
| • | Normalized expression ratio = 2‾ΔΔCT = 2‾4.3 = 0.05 | |
| • | PMNs treated with S. aureus expressed MPO at 0.05 fold lower than untreated PMNs | |
| • | PMNs+P. betle+S. aureus | |
| • | ΔCT (P) = Ct (target, P)-Ct (reference, P) = 29.10- 27.80 = 1.30 | |
| • | ΔCT (SPP) = Ct (target, SPP)-Ct (reference, SPP) = 25.31-25.30 = 0.01 | |
| • | ΔΔCt = ΔCt (SPP)-ΔCt (P) = 0.01-1.30 = -1.29 | |
| • | Normalized expression ratio = 2‾ΔΔCT = 2‾(-1.29) = 2.44 | |
| • | PMNs treated with P. betle ethanolic extract and S. aureus expressed MPO at 2.44 fold higher than untreated PMNs | |
Averaged result of all three RT-PCR runs: Based on the data quantification for the three trials (Table 3, Fig. 5), treated PMNs with P. betle and PMNs+P. betle+S. aureus showed increased MPO expression at 1.51 and 2.29 fold higher than untreated PMNs and treated PMNs with S. aureus (1×108 cell mL-1) showed decreased MPO expression at 0.053 fold lower than untreated PMNs (Fig. 5). It is confirmed that P. betle ethanolic extract relieves the suppressive effect of S. aureus on MPO expression in human neutrophils.
According to the amplification data for all three trial runs, PMNs treated with S. aureus (1×108 cells/mL) showed decreased MPO expression at 0.053 fold lower than untreated PMNs. However, MPO expression in PMNs treated with P. betle extract was higher than in untreated PMNs and PMNs treated with S. aureus indicating higher expression of the gene in the presence of the extract. Based on the quantification data for the three trials (Table 3), PMNs treated with P. betle and with both P. betle and S. aureus showed increased MPO expressions at 1.51 and 2.29 folds higher respectively than untreated PMNs (Fig. 5). This verifies that P. betle ethanolic extract relieves the suppressive effect of S. aureus on MPO expression in human neutrophils.
Total protein content: Total proteins in the lysates prepared from neutrophils were increased after exposure to MIC (5 mg mL-1) of P. betle ethanolic extract (6.2 μg mL-1) which represents a 19.2% increase compared to untreated PMNs (5.2 μg mL-1) (Fig. 1). In contrast, lysate obtained from neutrophils exposed to S. aureus alone showed decreased protein concentration (4.2 μg mL-1) corresponding to approximately 19.2% reduction compared to untreated cells. Further, the presence of P. betle extract caused an approximately 13.5% increase over untreated cells in total protein concentration in the neutrophils inspite of presence of S. aureus (5.9 μg mL-1).
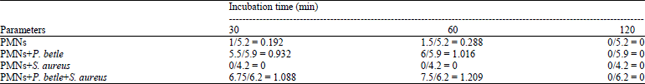
| Table 4: | Specific TNB consumption (nM mL-1 μg-1 protein) by lysates against baseline protein concentrations |
 | |
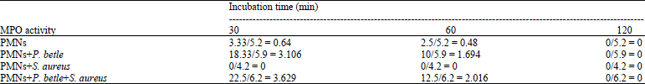
| Table 5: | Specific myeloperoxidase activity (mU mL-1 μg-1 protein) against baseline protein concentrations |
 | |
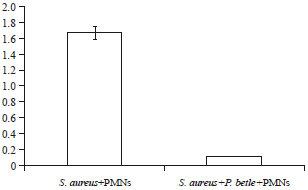 | |
| Fig. 6: | Neutrophil killing assay of S. aureus (1×108 cell mL-1) Graph showing colony counts (CFU) of S. aureus+PMNs and S. aureus+P. betle+PMNs |
TNB consumption: To determine MPO activity assay, the amount of 5-thio-2-nitrobenzoic acid (TNB) consumed by each lysate preparation was first measured. The TNB consumption is proportional to myeloperoxidase activity in test samples (Fig. 2). Neutrophils (PMNs) that were challenged with S. aureus (1×108 CFU mL-1) had no consumption of TNB whereas the presence of MIC (5 mg mL-1) of P. betle ethanolic extract significantly increased TNB consumption indicating elevated myeloperoxidase activity in PMNs above baseline (control) levels.
Specific TNB consumption (nM mL-1 μg-1 protein): Specific TNB consumption (nM mL-1 μg-1 protein) calculations were performed to standardize TNB consumption by myeloperoxidase to baseline protein concentration in the different test samples which revealed that TNB consumption by PMNs treated with P. betle was 0.932 and 1.016 at 30 and 60 min, respectively (Table 4). No TNB consumption was observed in PMNs treated with S. aureus. An increase in TNB consumption by MPO was observed in PMNs treated with P. betle and S. aureus (1.088 and 1.209) compared to untreated PMNs (0.192 and 0.288) at 30 and 60 min, respectively.
Myeloperoxidase (MPO) activity: Correspondingly, neutrophils that were challenged with S. aureus completely lost MPO activity compared to unchallenged neutrophils (Fig. 3). A marked increase in MPO activities was evident when the crude extract was added to PMNs in the presence and absence of S. aureus although a more significant increase was observed without presence of the organism. The decline in MPO activities beyond 30 min in all samples are as expected presumably due to deterioration of MPO protein subunits that occurs consistently in all samples.
Specific myeloperoxidase (MPO) activity (mU mL-1 μg protein): Specific MPO activity calculation was performed to standardize to baseline protein concentration which revealed that MPO activity in PMNs treated with P. betle were 3.106 and 1.694 at 30 and 60 min incubation, respectively (Table 5). No MPO activity was observed in PMNs treated with S. aureus and similarly, significant increases in MPO activities were observed in PMNs treated with P. betle alone or P. betle with S. aureus (3.629 and 2.016) compared to untreated PMNs.
Neutrophil killing assay: The PMNs that were challenged with opsonized S. aureus (2.12×108) were competent to perform killing of the pathogen whereupon after 60 min incubation, which allowed for phagocytotic events, the number of S. aureus cells recovered from PMN lysate was 1.68×108 CFU mL‾1 which represents 20.75% reduction (Fig. 6). However, PMNs that were challenged with the same number of S. aureus in the presence of P. betle ethanolic extract showed reduced recovery of viable organisms (0.12×108 cell mL-1) corresponding to 94.33% decrease in cell number.
DISCUSSION
Efficient killing of S. aureus by human neutrophils via the H2O2-dependent pathway is enhanced by the presence of myeloperoxidase (Humphreys et al., 1989). The NADH oxidase catalyses formation of cell damaging O2‾ which is subsequently converted to the highly toxic H2O2 that induces cell death. Myeloperoxidase catalyzes oxidation of chloride by H2O2 to form hypochlorous acid (HOCl) which contributes to microbicidal properties of PMNs (Lau and Baldus, 2006). The HOCl formed within the phagosomes react with ingested bacteria to generate tyrosyl radicals which catalyses cross-linking of proteins (Heinecke et al., 1993) and oxidises bacterial methionine residues in proteins that contribute to microbicidal effect (Rosen et al., 2009).
Reactive radicals such as O2‾, H2O2 and OH- are capable of killing organisms in the absence of myeloperoxidase (Roos et al., 2003; Urban et al., 2006). However, recent studies suggest a more prominent role of MPO in microbial killing within phagosomes that appears to be especially significant in overwhelming infections (Klebanoff et al., 2013). Myeloperoxidase-deficient mice for instance, are shown to be particularly more susceptible to candidiasis (Aratani et al., 1999). It has also been observed that neutrophils deficient in MPO kill many species of pathogenic microbes more slowly than do normal neutrophils and mice deficient in MPO succumb more readily to certain pathogens than do normal animals (Gresham et al., 2000; Palazzolo-Balance et al., 2008).
In overcoming and to survive the damaging effects presented by PMNs during phagocytosis (Rigby and DeLeo, 2013), S. aureus is known to regulate expression of genes encoding proteins that are involved in stress-response, metabolism and virulence, amongst others. Specifically, genes encoding γ-hemolysins (hlgA, hlgB and hlgC) and exotoxins that destroy leukocytes are upregulated during phagocytosis by neutrophils. In addition, S. aureus also expresses two pore-forming toxins, namely the α-hemolysin and bicomponent leukotoxins (Luk) that aids in destruction of neutrophils (Voyich et al., 2005). In this study, we show that S. aureus down-regulates MPO expression, probably contributing to the observed reduction in myeloperoxidase activity and its increased survival in human neutrophils. As these observations have not been previously documented in literature, therefore, an additional mechanism by which this pathogen overcomes killing by neutrophils is suggested. Majumdar et al. (2003) reported that P. betle ethanolic extract increased both superoxide dismutase and catalase activities NSAID induced ulcers in albino rats but effects of S. aureus on MPO expression has not been documented. P. betle ethanolic increased MPO expression, thereby alleviating the repressor effect of S. aureus to increase myeloperoxidase activity. The RT-qPCR analyses verified upregulation of MPO by P. betle ethanolic leaf extract above baseline, despite presence of the organism. Increasing MPO expression and myeloperoxidase activity, most likely contributes to ability of neutrophils to resist and combat against damages caused by S. aureus (Hampton et al., 1996). It is not clear why P. betle extract expressed MPO much higher in presence of S. aureus and this requires elucidation on the mechanisms involved during transcription of the gene in presence and absence of the organism.
Presence of S. aureus also showed corresponding decrease in myeloperoxidase activity, possibly contributing to the observed debilitating effect on neutrophil killing potential, thus resulting in increase in survival of the organism in PMNs. In absence of extract, the organism suppressed myeloperoxidase activity with concomitant increase in survival of the organism up to 80%. Presence of P. betle ethanolic leaf extract increased myeloperoxidase activity which corresponded with decreased S. aureus survival as reflected by reduced number of colonies as determined by the neutrophil killing assay. Although correlation between myeloperoxidase activity and cell survival within neutrophils was not directly established in this study, the observations are highly suggestive. Results of neutrophil killing assay are acceptable because S. aureus cells were opsonized with bovine serum albumin to enhance phagocytosis by PMNs and sufficient incubation time (1 h) was allowed for phagocytic activity to occur effectively (Deby-Dupont et al., 1999). In addition, after treatment of PMNs with S. aureus, the cell suspension was pelletted and washed twice with ice cold PBS to remove extracellular S. aureus cells. Therefore, the growth of cells observed after lysing treated PMNs were assuredly sourced from within (intracellular) the PMNs where extracellular unphagocytosed bacteria were removed (Haq et al., 1995).
Taken together, our observations confirm that P. betle ethanolic extract increases MPO activity by upregulating expression of the MPO in human neutrophils. Possible interference by ethanol on the observed effects is discounted due to the use of the rotary evaporator during extract preparation. Rotary evaporation vaporises residual ethanol at 60°C and condenses it under reduced pressure, where it is converted into liquid ethanol which was collected in a separate collection plate and discarded (Voravuthikunchai and Mitchell, 2008). Therefore, the extract is devoid of ethanol. The 10% DMSO is often used as a cyroprotectant to optimize cell survival in vitro and in vivo (Randhawa, 2006). Dimethyl sulfoxide was used in this study at 10% concentration which had no cytotoxic and inhibitory effect on S. aureus and neutrophils. In addition, relevant PMN and S. aureus controls that were included this study provided appropriate baseline data that reduced possible errors. Furthermore, our results showed enhanced enzyme activity and increased protein concentrations in the presence of the extract which negates the possible cytotoxic and inhibitory effects normally imposed by DMSO and ethanol.
Piper betel ethanolic extract has been previously characterized and its phytoconstituents and their related pharmacological properties identified (Bhalerao et al., 2013). We recently verified Allylpyrocatechol (APC) as the major constituent in P. betle ethanolic extract using HPLC UV and NMR, albeit at reduced purity (Abdullah and Hussain, 2015). APC was previously shown to exhibit anti-inflammatory activities by inhibition of the NF-КB pathway in LPS-induced macrophages (Sarkar et al., 2008). Its chemical structure which reveals two hydroxyl groups each attached to a benzene ring, facilitates its extraction as the major constituent in the more polar solvent, ethanol by given its immunomodulatory potential, further investigations using standard APC are warranted for confirmation of its direct contribution towards the observed effects.
CONCLUSION
In conclusion, this study showed that S. aureus inhibits MPO expression possibly contributing to the corresponding decrease in myeloperoxidase activity leading to high survival of the organism in PMNs. This observation suggests another mechanism by which S. aureus overwhelms and survives neutrophil killing. Piper betle ethanolic leaf extract alleviates the suppressive effect induced by S. aureus causing a high increase in MPO expression resulting in increased MPO activity and correspondingly, decreased survival of phagocytosed S. aureus. As we have previously established allylpyrocatechol as the major constituent in the extract, therefore, pending investigations using standard allylpyrocatechol of known purity, we expect to verify it as the major component in P. betle ethanolic extract that is responsible for the observed effects on neutrophils.
ACKNOWLEDGMENT
The authors thank the Postgraduate Student Division, Faculty of Health Sciences, Universiti Teknologi MARA for the assistance and funds provided to complete this study.
REFERENCES
- Abdullah, N. and R.M. Hussain, 2015. Isolation of allylpyrocatechol from Piper betle L. leaves by using high-performance liquid chromatography. J. Liquid Chromatogr. Related Technol., 38: 289-293.
CrossRefDirect Link - Aratani, Y., H. Koyama, S.I. Nyui, K. Suzuki, F. Kura and N. Maeda, 1999. Severe impairment in early host defense against Candida albicans in mice deficient in myeloperoxidase. Infect. Immunity, 67: 1828-1836.
Direct Link - Bhattacharya, S., S.R. Chaudhuri, S. Chattopadhyay and S.K. Bandyopadhyay, 2007. Healing properties of some Indian medicinal plants against indomethacin-induced gastric ulceration of rats. J. Clin. Biochem. Nutr., 41: 106-114.
CrossRefDirect Link - Bhalerao, S.A., D.R. Verma, R.V. Gavanka, N.C. Teli, Y.Y. Rane and V.S. Didwana and A. Trikannad, 2013. Phytochemistry, pharmacological profile and therapeutic uses of Piper betle Linn-An overview. Res. Rev.: J. Pharmacogn. Phytochem., 1: 10-19.
Direct Link - Chahal, J., R. Ohlyan, A. Kandale, A. Walia and S. Puri, 2011. Introduction, phytochemistry, traditional uses and biological activity of genus Piper: A review. Int. J. Curr. Pharm. Rev. Res., 2: 130-144.
Direct Link - Dahlgren, C., A. Karksson and J. Bylund, 2007. Measurement of respiratory burst products generated by professional phagocytes. Methods Mol. Biol., 412: 349-363.
PubMed - Humphreys, J.M., B. Davies, C.A. Hart and S.W. Edwards, 1989. Role of myeloperoxidase in the Killing of Staphylococcus aureus by human neutrophils: Studies with the myeloperoxidase inhibitor salicylhrdroxamic acid. J. Gen. Microbiol., 135: 1187-1193.
PubMed - Hampton, M.B., A.J. Kettle and C.C. Winterbourn, 1996. Involvement of superoxide and myeloperoxidase in oxygen-dependent killing of Staphylococcus aureus by neutrophils. Infect. Immunity, 64: 3512-3517.
Direct Link - Heinecke, J.W., W. Li, G.A. Francis and J.A. Goldstein, 1993. Tyrosyl radical generated by myeloperoxidase catalyzes the oxidative cross-linking of proteins. J. Clin. Invest., 91: 2866-2872.
CrossRefDirect Link - Kanjwani, D.G., T.P. Marathe, S.V. Chiplunkar and S.S. Sathaye, 2008. Evaluation of immunomodulatory activity of methanolic extract of Piper betel. Scand. J. Immunol., 67: 589-593.
CrossRefPubMedDirect Link - Klebanoff, S.J., 2005. Myeloperoxidase: Friend and foe. J. Leukocyte Biol., 77: 598-625.
CrossRefPubMedDirect Link - Kobayashi, S.D., K.R. Braughton, A.M. Palazzolo-Ballance, A.D. Kennedy and E. Sampaio et al., 2010. Rapid neutrophil destruction following phagocytosis of Staphylococcus aureus. J. Innate Immunity, 2: 560-575.
Direct Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - Lau, D. and S. Baldus, 2006. Myeloperoxidase and its contributory role in acute coronary syndrome and inflammatory heart disease. Pharmacol. Ther., 111: 16-26.
CrossRefDirect Link - Majumdar, B., S.G. Ray Chaudhuri, A. Ray and S.K. Bandyopadhyay, 2003. Effect of ethanol extract of Piper betle Linn leaf on healing of NSAID-induced experimental ulcer: A novel role of free radical scavenging action. Indian J. Exp. Biol., 41: 311-315.
PubMed - Nauseef, W.M., 2007. How human neutrophils kill and degrade microbes: An integrated view. Immunol. Rev., 219: 88-102.
CrossRef - Pin, K.Y., T.G. Chuah, A.A. Rashih, C.L. Law, M.A. Rasadah and T.S.Y. Choong, 2009. Drying of betel leaves (Piper betle L.): Quality and drying kinetics. Drying Technol., 27: 149-155.
Direct Link - Ramji, N., N. Ramji, R. Iyer and S. Chandrasekaran, 2002. Phenolic antibacterials from Piper betle in the prevention of halitosis. J. Ethnopharmacol., 83: 149-152.
CrossRefPubMedDirect Link - Randhawa, M.A., 2006. The effect of dimethyl sulfoxide (DMSO) on the growth of dermatophytes. Japanese J. Med. Mycol., 47: 313-318.
CrossRef - Rathee, J.S., B.S. Patro, S. Mula, S. Gamre and S. Chattopadhyay, 2006. Antioxidant activity of Piper betel leaf extract and its constituents. J. Agric. Food Chem., 54: 9046-9054.
CrossRefPubMedDirect Link - Rigby, M. and F.R. DeLeo, 2013. Neutrophils in innate host defense against Staphylococcus infections. Semin Immunopathol., 34: 237-259.
PubMed - Hussain, R.M., N.A.M. Din and N.A.N.M. Nasir, 2013. Piper betle ethanolic extract reduces Neutrophil scavenging ability and catalase activity in S.aureus. Int. J. Pharm. Sci. Rev. Res., 22: 35-40.
Direct Link - Roos, D., R. van Bruggen and C. Meischl, 2003. Oxidative killing of microbes by neutrophils. Microbes Infect., 5: 1307-1315.
CrossRef - Rosen, H., S.J. Klebanoff, Y. Wang, N. Brot, J.W. Heinecke and X. Fu, 2009. Methionine oxidation contributes to bacterial killing by the myeloperoxidase system of neutrophils. Proc. Nat. Acad. Sci., 106: 18686-18691.
CrossRef - Sarkar, D., P. Saha, S. Gamre, S. Bhattacharjee and C. Hariharan et al., 2008. Anti-inflammatory effect of allylpyrocatechol in LPS-induced macrophages is mediated by suppression of iNOS and COX-2 via the NF-κB pathway. Int. Immunopharmacol., 8: 1264-1271.
CrossRef - Strober, W., 2015. Trypan blue exclusion test of cell viability. Curr. Protocols Immunol., Vol. 21.
CrossRefDirect Link - Tan, Y.P. and E.W.C. Chan, 2014. Antioxidant, antityrosinase and antibacterial properties of fresh and processed leaves of Anacardium occidentale and Piper betle. Food Biosci., 6: 17-23.
CrossRefDirect Link - Urban, C.F., S. Lourido and A. Zychlinsky, 2006. How do microbes evade neutrophil killing? Cell. Microbiol., 8: 1687-1696.
CrossRefDirect Link - Venaille, T.J., N.L.A. Misso, M.J. Phillips, B.W.S. Robinson and P.J. Thompson, 1994. Effects of different density gradient separation techniques on neutrophil function. Scandinavian J. Clin. Laboratory Invest., 54: 385-391.
Direct Link - Voravuthikunchai, S.P. and H. Mitchell, 2008. Inhibitory and killing activities of medical plants against multiple antibiotic-resistant Helicobacter pylori. J. Health Sci., 54: 78-88.
Direct Link - Deby-Dupont, G., C. Deby and M. Lamy, 1999. Neutrophil myeloperoxidase revisited: It's role in health and disease. Intensivmedizin und Notfallmedizin, 36: 500-513.
CrossRefDirect Link - Kutter, D., P. Devaquet, G. Vanderstocken, J.M. Paulus, V. Marchal and A. Gothot, 2000. Consequences of total and subtotal myeloperoxidase deficiency: Risk or benefit ? Acta Haematol., 104: 10-15.
CrossRefPubMedDirect Link