
Kiam Beng Yeo
Materials and Minerals Research Unit, Faculty of Engineering, Universiti Malaysia Sabah, Jalan UMS, Kota Kinabalu, 88400, Sabah, Malaysia
Wong Wei Loong
Materials and Minerals Research Unit, Faculty of Engineering, Universiti Malaysia Sabah, Jalan UMS, Kota Kinabalu, 88400, Sabah, Malaysia
Kenneth Tze Kin Teo
Materials and Minerals Research Unit, Faculty of Engineering, Universiti Malaysia Sabah, Jalan UMS, Kota Kinabalu, 88400, Sabah, Malaysia
Journal of Applied Sciences
Year: 2014 | Volume: 14 | Issue: 23 | Page No.: 3242-3248







ABSTRACT
Fundamental fish swimming kinematic by sinusoidal fit consisting of smooth amplitude modulated travelling wave h(x,t) along the fish body from the nose to tail being a construe of wave amplitude envelope f(x) and a fish tail oscillation function g(t). Improved fundamental parameters by previous workers, such as the body wave speed (C), wavelength of body wave (λ), swimming speed (U), tail beat amplitude (A) and Strouhal-Number (St) were utilized in the simulation for designing the biomimetic robotic fish. Effects of these parameters on the swimming kinematic model had been developed for comparison between the Blacktip shark and biomimetic robot fish over a single cycle with 18 iteration of full fish body motion from nose to tail. Results showed that Blacktip shark could be seen to produce a more elongated body displacement shape as compared to the biomimetic robotic fish. Doubling the swimming speed (U) had also showed insignificant body motion change though increase in tail beat frequency and reduction of tail beat amplitude (A) and wavelength (λ) was obvious. The effect of doubling the tail beat frequency had also shown similar behavior as doubling the swimming speed.
PDF Abstract XML References Citation
Received: April 23, 2014;
Accepted: July 23, 2014;
Published: September 13, 2014
How to cite this article
Kiam Beng Yeo, Wong Wei Loong and Kenneth Tze Kin Teo, 2014. Biomimetic Robot Fish Modelling Base on Shark Swimming Kinematics. Journal of Applied Sciences, 14: 3242-3248.
DOI: 10.3923/jas.2014.3242.3248
URL: https://scialert.net/abstract/?doi=jas.2014.3242.3248
DOI: 10.3923/jas.2014.3242.3248
URL: https://scialert.net/abstract/?doi=jas.2014.3242.3248
INTRODUCTION
Increasing interest in biomimetic robotics attracted scientists and researchers in various applications for military and marine purposes such as undersea mission, search and rescue, cable inspection and nature disaster monitoring. In Mother Nature, fishes showed excellent performance in speed and maneuverability to swim in underwater environments. Thus, the inspirations to a huge pool of studies in fish locomotion in underwater environment.
However, studies on fish locomotion mainly focused on bony fishes that have skeleton made of bones which could be more closely modelled through the fish kinematics, morphology and functionality. Shark also categorized as a fish, with the exception of its skeletons being cartilages instead of bones. Its flexible tissue is durable and strong like bones, yet lighter and more flexible then bones. Shark swimming motion could be classified as between Anguilliform mode and Carangiform mode and so defined as a Subcarangiform swimmer.
In Massachusetts Institute of Technology (MIT), Tuna fish had inspired the RoboTuna development of a thunniform swimmer. The dimension and swimming kinematics of RoboTuna was based on the true Tuna fish (Barrett et al., 1999). In MIT, Kumph (2000) had also developed the autonomous robotic fish based on the Pike fish call RoboPike. The purpose of these robotic fish was to establish fluid mechanics interaction of fish swimming in liquid environment and the optimization performance of operation time and energy consumption.
Lighthill (1960) proposed a kinematic model for Carangiform propulsion and this model had been widely used in fish robot research. Liu and Hu (2010) design of a multi-joint Carangiform swimming robotic fish was such a case that utilizes the kinematic model to develop a tail motion function describing the fish tail motion relative to the head as discretized into a series of tail posture over time. As well as Yu and Wang (2005) numerically calculated the optimal link-length-ratio using improved constrained cyclic variable method applied to develop a four linked robotic fish.
SHARK SWIMMING MOTION
Webb and Keyes (1982) had recorded six species of shark swimming movement on video tape and analyses the video frame by frame manually to resolve the movement within 1/60 of a second or 17 m sec. They found that one of the Blacktip shark species had forward swimming movement swims at speed of 0.22 m sec-1 with a total body length of 0.97±0.5 m as illustrated in the Fig. 1.
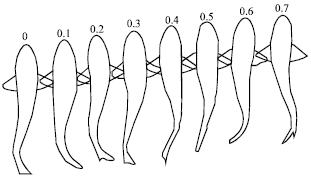 | |
| Fig. 1: | Blacktip shark forward swimming movement at speed of 0.22 m sec-1 (Webb and Keyes, 1982) |
The speed of the wave traveling backwards over the body was found greater than the swimming speed (Webb and Keyes, 1982).
Webb and Keyes (1982) showed that sharks did not swim at low speed but instead increases swimming speed by increasing the tail beat frequency while reducing the body movement amplitude. However, different species of sharks may swim at various speeds, tail beat frequency and body movement amplitude, their analysis found five important kinematic parameters, governing the shark from the nose tip to the tail tip with respect to a total body length (L). The parameters were the swimming speed (U/L), tail beat frequency (f), tail beat amplitude (A/L), stride length (S/L) and the wave length (λ/L). The measurement data being summarize in Table 1 for various shark types ranked according to their morphology and swimming mode from Anguilliform to Carangiform. Figure 2 also illustrated the relationship of kinematic parameters specific to Blacktip shark.
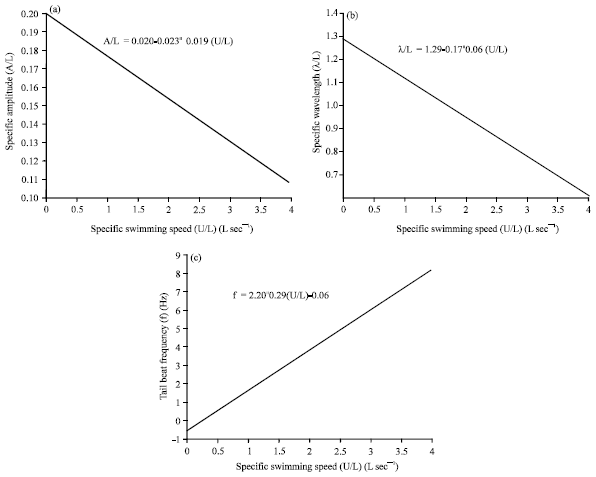 | |
| Fig. 2(a-c): | Relationships of (a) Tail beat amplitude (A/L), (b) Wavelength (λ/L) and (c) Tail-beat frequency (f) and swimming speed (U/L) of Blacktip shark |
| Table 1: | Summary of mean kinematic parameters for various species of shark |
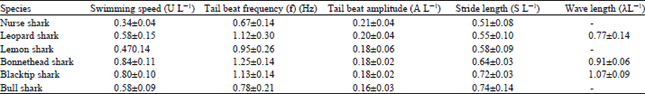 | |
| (Webb and Keyes, 1982) | |
KINEMATIC MODEL
Fishes produced a dynamic body motion to recreate a flow field effective for the fish to swim in the underwater environment. Thus, if the flow field could be reproduced for an undulated body and an oscillating tail fin of a biomimetic robot fish, then we could in principle achieve the performance of a real fish. However, the difficulties in developing this motion lie in two areas. First, the actual shape of the biological fish had never been measured accurately enough to use as a direct input to the biomimetic robotic, fish body control system. Even, if it had the biomimetic robotic fish body can only be a replicate copy of the biological fish shape. Secondly, attempting to derive the exact shape by mathematically analysis shall be limited by the complexity of the interaction of the biomimetic robot fish body with the surrounding fluid.
Therefore, to understand the fish swimming locomotion in the fluid environment, the fish swimming kinematic could be curve fitted as sinusoidal consisting of smooth amplitude modulated travelling wave along the fish bodies from the nose towards the tail illustrate in the Fig. 3. Gray (1933) discovered that the waves travel along the fish body could cause faster backward wave movement then the forward velocity of the fish and the amplitude of the wave was different for each section of the fish body.
As such, Lighthill (1960) and Wu (1971) had suggested the shape of the wave as following in Eq. 1:
| (1) |
where, h (x,t) is the traveling wave along the fish body and g(t) is the fish tail oscillation function. From Lighthill perspective, f(x) being the wave amplitude envelope which limits the large motion of the anterior section of the fish. In addition, he predicted that the function of f(x) being to minimise the momentum losses when the fish under wake swimming condition through keeping the body wave speed C close to the swimming speed.
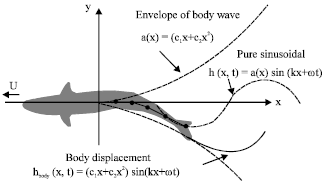 | |
| Fig. 3: | Top view of fish swimming kinematics |
Based on the Eq. 1 and the Fig. 3, Lighthill developed the following equations:
| (2) |
| (3) |
| (4) |
The kinematic model originally was developed from pure sinusoidal function consisting of wave peak amplitude along the body length. The function a(x) being the body wave amplitude envelope, such that C1 being chosen to be independent and C2 as to double the amplitude of the motion, while both C1 and C2 were adjustable parameters. According to Barrett (1996), the sinusoidal wave travel from right to left within the bounds of the second order amplitude envelope. However, in this study, the calculation of the swimming kinematic model had been based on sinusoidal wave traveling from left to right.
Combining the sinusoidal equation and the body wave amplitude envelope equation shall yield the body displacement function h(x, t) of the fish swimming motion. The motion was measured from the fixed body position until the end of the tail along the axis. Barratt had improved on Lighthill’s kinematic model by combining idea from Dewar and Graham (1994) and Triantafyllou et al. (1993). Important parameters were collected from their works, such as the speed of the body wave (C), wavelength of body wave (λ), swimming speed (U), tail beat amplitude (A) and Strouhal-Number (St). It is worth noting that the derivation of governing equation was calculated based on sinusoidal wave travelling from left to right and the parameters had been changed to suite this kinematic model.
The relationships of these parameters of the swimming kinematics were found by evaluating Eq. 4 at horizontal axis or at x = the fish body length lbody, such that:
| (5) |
| (6) |
| (7) |
At this level, five parameters were obtained experimentally; U, St, A, λ and C1. From the definition, seven other related parameters could be identified:
![]()
| (8) |
| (9) |
| (10) |
According to Webb and Keyes (1982), changes of shark swimming speed affects the tail beat frequency, body amplitude and wavelength. Therefore, Eq. 5 could be expanded to obtain the relationship among the parameters shown in the Fig. 3:
| (11) |
Where, hbody is a function of C1, C2, lbody, k, t and ω only could be determined except for the C2. Therefore, to obtain the C2, the Eq. 6 could be utilized by solving numerically to yield:
| (12) |
RESULTS AND DISCUSSION
The fundamental parameters used by the kinematic model was developed to describe the fish swimming motion which was required for designing the biomimetic robotic fish included {C1, λ, St, f, U, A}. The C1 being the linear coefficient of the wave amplitude envelope traveling along the body, λ is wavelength of the fish traveling body wave, St is Strouhal-Number of the fish tail, f is oscillation frequency generate from fish body to actuate the tail, U is fish forward swimming speed and A is the tail beat amplitude of the fish. Effect of the parameters on the swimming kinematic model has been presented in two sets of graph to display the comparison between Blacktip shark and biomimetic robot fish for a single cycle with 18 iteration of full fish body motion from nose to tail. In Table 2, the swimming kinematic parameters for designing the biomimetic robotic fish being based on the true experiment of the Blacktip shark.
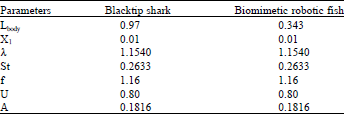
Figure 4 showed the Blacktip shark and biomimetic robot fish swimming motion by using base parameters which produces a body wave speed of 1.3386 L sec-1 for a wave period of 0.8621 sec. The Blacktip shark could be seen to produce a more elongated body displacement shape as compared to the biomimetic robotic fish.
Due to the natural characteristic of real shark by doubling the swimming speed (U) as in Fig. 5, increases the tail beat frequency to 2.29 Hz could be observed and a reduction of tail beat amplitude A to 0.1632 and wavelength λ to 1.0180 took place. In addition, the body wave speed had doubled to 2.9726 L sec-1 and the wave period reduced to 0.3425 sec. Yet the body motion still remain the same even the swimming speed had been doubled.
In the Fig. 6, doubling the tail beat frequency f had increased the swimming speed to 1.3507 L sec-1 and the body wave speed to 0.4310 L sec-1 within a period of 0.4310 sec. In fact, increased of tail beat frequency had caused the tail beat amplitude to reduce to 0.1689 and wavelength decrease to 1.0689.
| Table 2: | Parameters of Blacktip shark and biomimetic robotic fish for kinematic simulation |
 | |
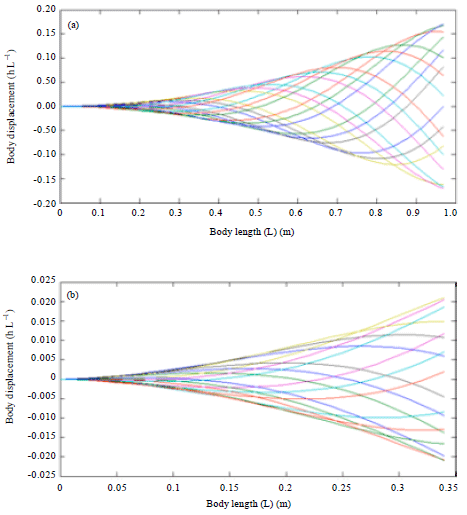 | |
| Fig. 4(a-b): | Body kinematic simulation using base parameter of (a) Black tip shark and (b) Robotic fish |
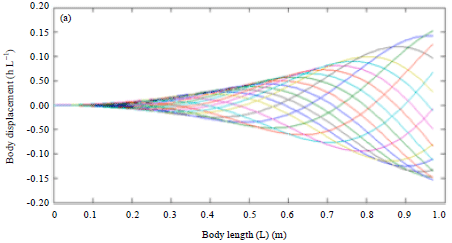 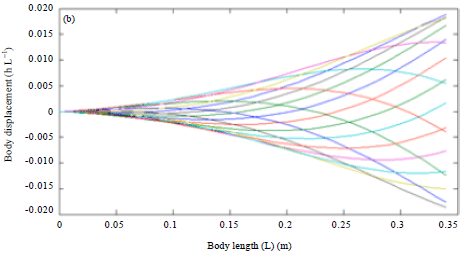 | |
| Fig. 5(a-b): | Effect of double swimming speed on body kinematics of (a) Blacktip shark body kinematics and (b) Robot fish |
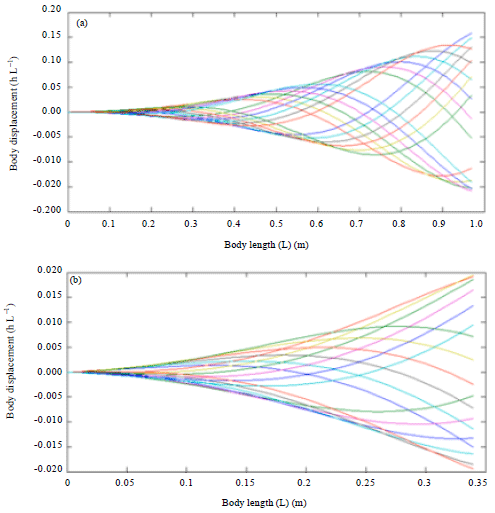 | |
| Fig. 6(a-b): | Effect of double tail beat frequency on body kinematics of (a) Blacktip shark body kinematics and (b) Robot fish |
The effect of doubling the tail beat frequency showed similar effect of doubling the swimming speed.
CONCLUSION
The equation of kinematic model presented in this study had shown that it could be utilized for simulating both real shark and biomimetic robot fish swimming performance. Results showed that Blacktip shark could be seen to produce a more elongated body displacement shape as compared to the biomimetic robotic fish. The fish body motion was also found to remain the same even the swimming speed had been doubled. As well as the effect of doubling the tail beat frequency showed similar effect as doubling the swimming speed. However, other aspects of fish hydrodynamic drag force, shape and muscle performance which could limit the accuracy of the kinematic model had not been considered. Nevertheless, the equation of motion could be used as design and simulation references for biomimetic robotic fish the performance prediction.
ACKNOWLEDGMENT
The authors expressed their appreciation to the Material and Minerals Research Unit, Faculty of Engineering of Universiti Malaysia Sabah for the support of this study.
NOMENCLATURE
| A | = | Tail Beat amplitude |
| C | = | Body wave speed |
| c1 | = | Linear body wave amplitude envelope |
| c2 | = | Quadratic body wave amplitude envelope |
| f | = | Tail beat frequency |
| hbody | = | Body displacement |
| k | = | Body wave number |
| L | = | Shark body length |
| lbody | = | Body length |
| St | = | Strouhal number |
| T | = | Period body wave |
| U | = | Swimming speed |
| ω | = | Body wave frequency |
| x | = | Displacement along main axis |
| λ | = | Body wave length |
REFERENCES
- Barrett, D.S., M.S. Triantafyllou, D.K.P. Yue, M.A. Grosenbaugh and M.J. Wolfgang, 1999. Drag reduction in fish-like locomotion. J. Fluid Mech., 392: 183-212.
CrossRefDirect Link - Dewar, H. and J. Graham, 1994. Studies of tropical tuna swimming performance in a large water tunnel-energetics. J. Exp. Biol., 59: 45-59.
Direct Link - Lighthill, M.J., 1960. Note on the swimming of slender fish. J. Fluid Mech., 9: 305-317.
CrossRefDirect Link - Liu, J. and H. Hu, 2010. Biological inspiration: From carangiform fish to multi-joint robotic fish. J. Bionic Eng., 7: 35-48.
CrossRef - Triantafyllou, G.S., M.S. Triantafyllou and M.A. Grosenbaugh, 1993. Optimal thrust development in oscillating foils with application to fish propulsion. J. Fluids Struct., 7: 205-224.
CrossRefDirect Link - Wu, T.Y.T., 1971. Hydromechanics of swimming propulsion. Part 3. Swimming and optimum movements of slender fish with side fins. J. Fluid Mech, 46: 545-568.
Direct Link - Yu, J. and L. Wang, 2005. Parameter optimization of simplified propulsive model for biomimetic robot fish. Proceedings of the IEEE International Conference on, Robotics and Automation, April 18-22, 2005, Barcelona, Spain, pp: 3306-3311.
CrossRefDirect Link