
Thanchanok Kangwankraiphaisan
Center of Excellence for Environmental and Hazardous Waste Management, Chulalongkorn University, Bangkok 10330, Thailand
Kallaya Suntornvongsagul
Center of Excellence for Environmental and Hazardous Waste Management, Chulalongkorn University, Bangkok 10330, Thailand
Journal of Applied Sciences
Year: 2013 | Volume: 13 | Issue: 5 | Page No.: 743-748







ABSTRACT
The association of Lindenbergia philippensis (Cham.) Benth. and Arbuscular Mycorrhizal Fungi (AMF) enhanced plant tolerance to high concentration of Cu by reducing the accumulation of Cu in plant root. The aim of the study was to study Cu accumulation in the zinc smelting sediment that occurred in the rhizosphere sediment and root associated with AMF. The plant was grown in the sediment containing low level of nutrients (0.86±0.03% of Total Organic Carbon (TOC), 1.15±0.40 mg kg-1 dried sediment of nitrate (NO3-N) and 5.67±5.51 mg kg-1 dried sediment of phosphorus (P)) and high concentration of heavy metals under weak basic condition (pH 7.79-8.28). High level of Cu accumulation was detected in the rhizosphere suggesting that the biological function of the plant and AMF may limit the excess Cu from reaching into plant cells. Solid particle sheath containing Cu showed rectangular and amorphous particles was observed around rhizosphere zone by SEM-EDS. Relatively lower Cu concentration was detected in plant root with active AMF (T1) comparing to plant root with suppressed AMF (T2). Interestingly, higher Cu concentration was observed in rhizosphere of plant with active AMF whereas the Cu was lower in rhizosphere of the sample without active AMF. These results indicated that Cu in the rhizosphere is influenced by the function of AMF. The results of this study provide useful insight on Cu accumulation with the presence of association between AMF and dominant plant and may be considered as a valuable tool for phytostabilization.
PDF Abstract XML References Citation
Received: February 01, 2013;
Accepted: March 18, 2013;
Published: June 28, 2013
How to cite this article
Thanchanok Kangwankraiphaisan and Kallaya Suntornvongsagul, 2013. Cu Accumulation in the Rhizosphere of Lindenbergia philippensis (Cham.) Benth. Growing in the Contaminated Sediment. Journal of Applied Sciences, 13: 743-748.
DOI: 10.3923/jas.2013.743.748
URL: https://scialert.net/abstract/?doi=jas.2013.743.748
DOI: 10.3923/jas.2013.743.748
URL: https://scialert.net/abstract/?doi=jas.2013.743.748
INTRODUCTION
Heavy metals resulting from mining, smelting, agriculture and other heavy industries are important contaminants to the environment and living organisms. Copper (Cu) one of the heavy metals involves with several plant metabolic processes since it is an indispensible component of many oxidative enzymes involving photosynthesis and other cellular structures such as cell wall (Maksymiec, 1998). Although, Cu is one of the essential trace elements required for the survival of plants, excessive amount of Cu can cause damages to cells (Lou et al., 2004; Wei et al., 2008) and affect the growth and development of plant (Madejon et al., 2009).
However, the adverse effects of Cu are varied depending on the type of plants as well as other physicochemical factors including Cu speciation, bioavailability of Cu and soil pH (Tao et al., 2003; Chaignon et al., 2009). Plant tolerance toward elevated level of Cu was reported in some studies (Lou et al., 2004; Santibanez et al., 2008; Wei et al., 2008). The differences in plant physiological characteristics including metabolic processes, gene expression and excretion of certain substances such as exudates may contribute to the various plant responses to Cu (Hall, 2002; Jonak et al., 2004). In addition to plant physiology, the mutualistic relationship between plant and the microorganisms in root-soil interface or rhizosphere was purposed as an important factor for the observed resistance to high concentration of Cu.
Arbuscular Mycorrhizal Fungi (AMF) are normally found in rhizosphere and are crucial for plant survival under extreme environments. The AMF and other rhizosphere microorganisms receive the necessary soluble carbon based nutrients from plant and in return, they facilitate other mineral nutrient uptakes from the surrounding environment of the plants (Dandan and Zhiwei, 2007). Moreover, the rhizospheric microorganisms also enhance plant resistance toward pathogenic microorganisms and plant survivability under stressful conditions. The importance of AMF on the plant susceptibility to copper was revealed in many studies (Chen et al., 2007; Zhang et al., 2009). Evidences showed that microorganism community in the rhizosphere contributes to the changes in soil pH and speciation of Cu and therefore, creates fractionation of copper and reduce the amount of accumulated Cu in plant cells (Cloutier-Hurteau et al., 2008; Zhang et al., 2009). Lower level of Cu was observed in plant shoot with the presence of AMF comparing to plant without AMF indicating an important protective role of AMP under high Cu concentration (Chen et al., 2007). In this study, the accumulation of Cu from the zinc smelter sediment in Lindenbergia philippensis (Cham.) Benth. with and without the active AMF was investigated.
MATERIALS AND METHODS
Study area: The sampling site was located in the settling pond at the zinc smelter, Tak province (Northern part of Thailand). The settling pond received sediment containing heavy metals from smelting processes. Subsequently, the leachate and the solid fractions of the sediment were separated and sent to wastewater treatment facility and secure landfill, respectively. Five plots in the settling pond (1x1 m2 each) covered with L. philippensis (Cham.) Benth. was collected sediment, rhizosphere sediment and plant roots. The samples were preserved under 4°C until analysis of the characteristics of sediment, root colonization by AMF, solid sheath on the surface root and the Cu concentration in the root and the rhizosphere sediment.
Experimental setup: Sediment was collected from the settling pond. Approximately 4 kg (wet weight) of sediment was homogenized and filled into plastic pots. The 45 days old seedling of L. philippensis (Cham.) Benth. which were grown on the peat moss (Pindstrup) was transferred into the pots containing the obtained sediment. The experimental setup consisted of two treatments with five replicates for each treatment. The two treatments were treatment with active AMF (T1) and treatment without active AMF by applying fungicide (captan: Orthocide, Arysta LifeScience) 100 mg kg-1 (wet weight sediment) every week for suppressing the active AMF (T2). Addition of this captan can reduce the root colonization by AMF about 50% in the preliminary study. (O'Neill and Mitchell, 2000) demonstrated that the root colonization by AMF could be reduced to similar level as without inoculated of AMF by applying captan. The sediment in the pots was added with tap water regularly to maintain the moisture at 60-70%. The pots were rearranged randomly once a week. After 6 months of experiment, the rhizosphere sediment and plant root were collected and analyzed for the amount of Cu.
Sample analysis: The sediment, rhizosphere sediment and plant roots were dried at 80°C until weight was constant. As for the preparation for Cu concentration analysis, the dried samples were grounded in a mortar and sieved (74 μm, 200 mesh) and subjected to measure properties of the sediment and heavy metal analysis. The fresh roots were separated to measure the root colonization by AMF and solid sheath by scanning electron microscope and dispersive X-ray spectrometer (SEM-EDS).
The properties of the sediment collected from the settling pond were determined. The pH of liquid solution was measured by using the HACH® SensION™ Waterproof pH/mV/ISE Meter (HACH Company, Loveland, CO, USA). The texture from the density of the suspension was determined by the hydrometer technique (Kettler et al., 2001) and the Cation Exchange Capacity (CEC) was measured in according to the U.S. EPA method, 9080 by using ammonium acetate. Total N and NO3-N were analyzed by adding 2 N of KCl into 10 g of sediment and titrated with 0.005 N H2SO4 (Ryan et al., 2001). The P was measured by using the sodium bicarbonate (Olsen) method (pH>7) and the ammonium heptamolybdate-ammonium vanadate was applied for determining available P (Ryan et al., 2001). TOC and total N were analyzed by using a total organic carbon analyzer (TOC-SSM 5000A, Shimadzu-Module) following U.S. EPA method 9060. Total microorganisms including bacteria, actinomycetes and fungi in the sediment were determined by standard spread plate technique with serial dilutions (Maier et al., 2000; Pepper and Gerba, 2005). Total Cu of the samples was determined by U.S. EPA method 3050B using 65%HNO3 and 30%H2O2 and Atomic Absorption Spectrometry (AAS).
The root colonization of AMF was determined by the gridline intersection method following Brundrett et al. (1996). Approximately 10 g of fresh roots were suspended in 0.1% sodium pyrophosphate and sonicated to remove adherent rhizosphere sediment particles. The freshly cleaned roots were prepared and the root surface was cleared with 10% KOH and stained in trypan blue solution in lactoglycerol over night. The root colonization was observed by light microscope.
The selected small pieces of fresh roots with rhizosphere samples were completely dried in desiccators until without moisture. Then the dried samples were coated with carbon before subjected to scanning electron microscopy with an energy dispersive, X-ray spectrometer (SEM-EDS) JEOL, model JSM-5410LV and 5800LV (Japan) to study the solid particle sheath (Wullstein and Pratt, 1981; Kodama et al., 1994).
Statistic analysis: All data are reported as means±standard deviation. The significant differences were accepted when the values of alpha level are less than 0.05. The statistical model of SPSS 16.0 (LEAD Technologies Inc., Chicago, IL) was used for all statistical tests.
RESULTS
Sampling site: The analysis of the obtained settling pond sediment revealed that the sediment was weak basic, (pH 7.79-8.28) with small amount of nutrients and microorganisms and contained high heavy metal concentrations (Table 1). L. philippensis (Cham.) Benth. was the dominant species and two species of AMF (Suntornvongsagul et at., 2011) was found associated with its root (Fig. 1a, b).
Cu accumulation in the rhizosphere and root
Concentration of Cu in the rhizosphere and root of the settling pond: The concentration of Cu accumulated in the rhizosphere sediment and plant root at the settling pond was measured. The detectable Cu concentration was 403.70±158.24 mg kg-1 of dried sample and 104.05±37.37 mg kg-1 of dried sample for rhizosphere sediment and plant root tissue, respectively. Statistical analysis using paired t-tests of Cu concentrations suggested that the Cu concentration in rhizosphere sediment was significantly higher than the level observed in plant root (p<0.05).
Solid particle sheath in the rhizosphere: Solid particle sheath was found cover the root surface at the rhizosphere (Fig. 2).
| Table 1: | Physical and chemical characteristics of the contaminated sediment |
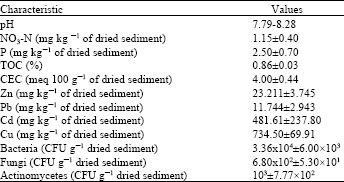 | |
The dense solid particles occurred in the rhizosphere might result from root exudates which contain the complex polysaccharide forming a layer of mucilage, enzymes, ethylene, sugars, organic acids, amino acids, root cap (Chaboud and Rougier, 1990; Xu et al., 2007) and dead cell lysates (Marschner, 2002).
With the presence of the exudates, Cu might be trapped into the substances and integrated in the solid particle sheath.
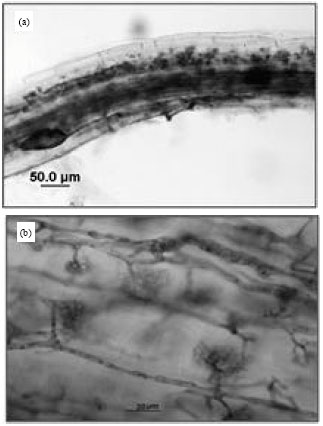 | |
| Fig. 1(a-b): | Image of (a) Vesicle (20x) and (b) Arbuscue (40x) of AMF in the L. philippensis (Cham.) Benth. root observed by light microscope |
 | |
| Fig. 2: | Plant root of L. philippensis (Cham.) Benth. with rhizosphere sediment observed with light stereo microscope (20x) |
In order to elucidate this, samples possibility were analyzed by SEM-EDS and the results indicated Cu contributed to 1.89% of element in the solid sheath (Fig. 3a, b).
Role of AMF on Cu accumulation: Higher concentration of Cu in plant root was detected in T2 in which the AMF were relatively inactive comparing to T1 (Fig. 4).
 | |
| Fig. 3(a-b): | (a) SEM photograph (magnified 500x) and (b) elemental analysis by EDS of the rhizosphere sediment |
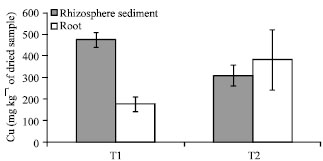 | |
| Fig. 4: | Comparison of Cu concentrations (mean value) of the control (T1) and control with captan (T2) of rhizosphere sediment and plant root. Bar and error bars represent mean (n = 5) and standard deviation values |
In T1 and T2, the detectable Cu concentration was 177.69±49.11 and 381.39±140.70 mg kg-1 of dried sample, respectively. On the contrary, higher accumulation of Cu was found in the rhizosphere sediment of T1 (437.75±36.63 mg kg-1 of dried sample) comparing to T2 (309.74±33.85 mg kg -1 of dried sample). In summary, samples with active AMF accumulated less Cu in root tissue, whereas more Cu was detected in their rhizosphere sediment comparing to the samples without active AMF. These results suggested that the lower Cu concentration observed in T1 may due to the boundary effect created by the active AMF in the rhizosphere sediment.
DISCUSSION
In this study area, L. philippensis (Cham.) Benth. was found to be the dominant species suggesting its ability to tolerate and grow under high Cu concentration. These results can be explained by Cu being accumulate in the rhizosphere zone which is the biological function may protect the excess Cu into the plant cell.
The solid particle sheath covering the plant root may contribute to plant tolerance toward Cu since part of the Cu in the soil environment was immobilized into the solid sheath as indicated by Cu content in the solid sheath. In addition to the effect of exudates, the association between plant and microorganisms may be a crucial factor as well (Sung et al., 2006; Cardon and Whitbeck, 2007). Several study showed that certain microorganisms which obtain the nutrients from root exudates (Chaboud and Rougier, 1990; Xu et al., 2007), may affect root metabolic processes increasing its resistance to extreme stresses (Bais et al., 2006).
The role of AMF on plant resist to heavy metals was previously described in several studies. Andrade et al. (2009) and Wu et al. (2010) demonstrated that AMF could protect and increase plant tolerance against heavy metals. Nevertheless, Zhang et al. (2009) and Wei et al. (2008) observed that AMF may increase Cu accumulation in root cell wall in some plant species.
In this study, samples with active AMF revealed less Cu accumulation in root tissue together with elevated level of Cu rhizosphere sediment comparing to the samples without active AMF. This suggested that, in this case, active AMF increase plant tolerance to Cu by reducing the amount of accumulated Cu in root cell by increasing the binding of Cu to the rhizosphere sediment.
CONCLUSION
In this study, the association of L. philippensis (Cham.) Benth. and AMF revealed increase in plant tolerance to high concentration of Cu by reducing the accumulation of Cu in plant root. This suggested the protective role of AMF on this plant against Cu and may be considered as a valuable tool for the study of phytostabilization of heavy metals. In order to understand the mechanisms in which these AMF contribute to the plant protection, more insight on the interaction between microorganisms and the rhizosphere environment should be studied in future work.
ACKNOWLEDGMENTS
This research was supported by the 90th Anniversary of Chulalongkorn University Fund (Ratchadaphiseksomphot Endowment Fund) and the Graduate School and supported as part of the Thai Research Fund (MRG5180311). It is also supported by instrumental and laboratory services of the Center of Excellence for Environmental and Hazardous Waste Management, the Environmental Research Institute, the Department of Microbiology and the Department of Botany, Chulalongkorn University, the Scientific Technological Research Equipment Center, Thailand.
REFERENCES
- Andrade, S.A.L., P.L. Gratao, M.A. Schiavinato, A.P.D. Silveira, R.A. Azevedo and P. Mazzafera, 2009. Zn uptake, physiological response and stress attenuation in mycorrhizal jack bean growing in soil with increasing Zn concentrations. Chemosphere, 75: 1363-1370.
CrossRef - Bais, H.P., T.L. Weir, L.G. Perry, S. Gilroy and J.M. Vivanco, 2006. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol., 57: 233-266.
CrossRefPubMedDirect Link - Chaboud, A. and M. Rougier, 1990. Comparison of maize root mucilages isolated from root exudates and root surface extracts by complementary cytological and biochemical investigations. Protoplasma, 156: 163-173.
CrossRef - Chaignon, V., M. Quesnoit and P. Hinsinger, 2009. Copper availability and bioavailability are controlled by rhizosphere pH in rape grown in an acidic Cu-contaminated soil. Environ. Pollut., 157: 3363-3369.
CrossRef - Chen, B.D., Y.G. Zhu, J. Duan, X.Y. Xiao and S.E. Smith, 2007. Effects of the arbuscular mycorrhizal fungus Glomus mosseae on growth and metal uptake by four plant species in copper mine tailings. Environ. Pollut., 147: 374-380.
CrossRefDirect Link - Cloutier-Hurteau, B., S. Sauve and F. Courchesne, 2008. Influence of microorganisms on Cu speciation in the rhizosphere of forest soils. Soil Biol. Biochem., 40: 2441-2451.
CrossRef - Dandan, Z. and Z. Zhiwei, 2007. Biodiversity of arbuscular mycorrhizal fungi in the hot-dry valley of the Jinsha river, Southwest China. Applied Soil Ecol., 37: 118-128.
CrossRef - Hall, J.L., 2002. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot., 53: 1-11.
CrossRefDirect Link - Jonak, C., H. Nakagami and H. Hirt, 2004. Heavy metal stress. Activation of distinct mitogen-activated protein kinase pathways by copper and cadmium. Plant Physiol., 36: 3276-3283.
CrossRefDirect Link - Kettler, T.A., J.W. Doran and T.L. Gilbert, 2001. Simplified method for soil particle-size determination to accompany soil-quality analyses. Soil Sci. Soc. Am. J., 65: 848-852.
CrossRefDirect Link - Kodama, H., S. Nelson, A.F. Yang and N. Kohyama, 1994. Mineralogy of rhizospheric and non-rhizospheric soils in corn fields. Clay Clay Miner., 42: 755-763.
CrossRefDirect Link - Lou, L.Q., Z.G. Shen and X.D. Li, 2004. The copper tolerance mechanisms of Elsholtzia haichowensis, a plant from copper-enriched soils. Environ. Exp. Bot., 51: 111-120.
CrossRef - Madejon, P., J.E. Ramirez-Benitez, I. Corrales, J. Barcelo and C. Poschenrieder, 2009. Copper-induced oxidative damage and enhanced antioxidant defenses in the root apex of maize cultivars differing in Cu tolerance. Environ. Exp. Bot., 67: 415-420.
CrossRef - Maksymiec, W., 1998. Effect of copper on cellular processes in higher plants. Photosynthetica, 34: 321-342.
CrossRefDirect Link - O'Neill, J.J.M. and D.T. Mitchell, 2000. Effects of benomyl and captan on growth and mycorrhizal colonization of Sitka-spruce (Picea sitchensis) and ash (Fraxinus excelsior) in Irish nursery soil. For. Pathol., 30: 165-174.
CrossRef - Ryan, J., G. Estefan and Abdul Rashid, 2001. Soil and Plant Analysis Laboratory Manual. 2nd Edn., ICARDA, Syria, Pages: 172.
Direct Link - Santibanez, C., C. Verdugo and R. Ginocchio, 2008. Phytostabilization of copper mine tailings with biosolids: Implications for metal uptake and productivity of Lolium perenne. Sci. Total Environ., 395: 1-10.
CrossRef - Sung, K., J. Kim, C.L. Munster, M.Y. Corapcioglu, S. Park, M.C. Drew and Y.Y. Chang, 2006. A simple approach to modeling microbial biomass in the rhizosphere. Ecol. Mod., 190: 277-286.
CrossRef - Suntornvongsagul, K., T. Kangwankraiphaisan, J. Piapukiew, P. Sihanonth and G.M. Gadd, 2011. Heavy metal crystallization in the rhizosphere of Lindenbergia philippensis (Cham.) Benth. on zinc smelter sediment. Proceedings of the 17th Thailand-Korea Conference on Environmental Engineering, January 20-21, 2011, Bangkok, Thailand.
- Tao, S., Y.J. Chen, F.L. Xu, J. Cao, B.G. Li, 2003. Changes of copper speciation in maize rhizosphere soil. Environ. Pollut., 122: 447-454.
CrossRef - Wei, L., C. Luo, X. Li and Z. Shen, 2008. Copper accumulation and tolerance in Chrysanthemum coronarium L. and Sorghum sudanense L. Arch. Environ. Contam. Toxicol., 55: 238-246.
Direct Link - Wu, F.Y., Y.L. Bi, H.M. Leung, Z.H. Ye, X.G. Lin and M.H. Wong, 2010. Accumulation of As, Pb, Zn, Cd and Cu and arbuscular mycorrhizal status in populations of Cynodon dactylon grown on metal-contaminated soils. Applied Soil Ecol., 44: 213-218.
CrossRefDirect Link - Wullstein, L.H. and S.A. Pratt, 1981. Scanning electron microscopy of rhizosheaths of Oryzopsis hymenoides. Am. J. Bot., 68: 408-419.
Direct Link - Xu, W.H., H. Liu, Q.F. Ma and Z.T. Xiong, 2007. Root exudates, rhizosphere Zn fractions and Zn accumulation of ryegrass at different soil Zn levels. Pedosphere, 17: 389-396.
CrossRef - Zhang, X.H., A.J. Lin, Y.L. Gao, R.J. Reid, M.H. Wong and Y.G. Zhu, 2009. Arbuscular mycorrhizal colonisation increases copper binding capacity of root cell walls of Oryza sativa L. and reduces copper uptake. Soil Biol. Biochem., 41: 930-935.
CrossRef