
Charles O. Nwuche
Department of Microbiology, University of Nigeria, Nsukka, Nigeria
Hideki Aoyagi
Department of Bioscience and Bioengineering, Graduate School of Life and Environmental Sciences, University of Tsukuba, Ibaraki, Japan
James C. Ogbonna
Department of Microbiology, University of Nigeria, Nsukka, Nigeria
Journal of Applied Sciences
Year: 2013 | Volume: 13 | Issue: 24 | Page No.: 5661-5671







ABSTRACT
An integrated treatment and valorization of Palm Oil Mill Effluent (POME) by Aspergillus terreus IMI 282743 immobilized on Luffa sponge was investigated. Effects of POME concentrations and nitrogen supplementation on Chemical Oxygen Demand (COD) reduction, microbial lipase and biomass production were evaluated in batch cultures. A 50% POME promoted the highest lipolytic activities in both immobilized and free cell cultures. In the former, the maximum lipase activity was 5.14 U mL-1 but in the non-immobilized batch, it was only 2.10 U mL-1. Lipase activities were low in the 25 and 100% POME due to overdilution and presence of inhibitory compounds, respectively. The pH was unchanged in the 100% POME but in other cultures, there were significant increase in the pH values. The pH of the 75% POME increased after a 48 h lag but in the 25 and 50% POME, the pH rose from 4-6.43 within a period of 96 h. COD did not change in the 100% POME but in the 75% POME, a 60.7% reduction was achieved. The COD of both the 50 and 25% POME decreased by 45% respectively. The immobilized biomass concentration was highest in the 75% POME (0.83 g L-1) but in the 25 and 100% POME, it was 0.27 and 0.63 g L-1, respectively. Supplementation of the 50% POME with a mixture of ammonium sulphate and yeast extract increased lipase production to10.6 U mL-1, biomass concentration to 3.7 g L-1 while the COD decreased by 80%. Lipase production from POME could be economically competitive to present industrial processes and provides additional incentive of treatment that is cheap and sustainable.
PDF Abstract XML References Citation
Received: January 30, 2013;
Accepted: October 29, 2013;
Published: January 25, 2014
How to cite this article
Charles O. Nwuche, Hideki Aoyagi and James C. Ogbonna, 2013. Lipase Production from Palm Oil Mill Effluent by Aspergillus terreus Immobilized
on Luffa Sponge. Journal of Applied Sciences, 13: 5661-5671.
DOI: 10.3923/jas.2013.5661.5671
URL: https://scialert.net/abstract/?doi=jas.2013.5661.5671
DOI: 10.3923/jas.2013.5661.5671
URL: https://scialert.net/abstract/?doi=jas.2013.5661.5671
INTRODUCTION
Microbial lipases play important roles in the hydrolysis of long chain triglycerides to intermediate and short chain di and monoglycerides, free fatty acids and glycerol (Prazeres et al., 2006; Babu and Rao, 2007). Apart from hydrolysis, lipases are also involved in a wide range of reversible conversion reactions. This makes lipases the enzyme of choice for potential application in the food, detergent, pharmaceutical, leather, textile, cosmetic and paper industries (Houde et al., 2004; Gupta et al., 2004). Lipases occur widely in nature and have been found in many species of animals (Shan et al., 2009), plants (Paques et al., 2008), bacteria, yeasts and fungi (Melo et al., 2005). However, microbial lipases are preferred because they are stable, safe and more useful than those derived from plant and animals because of the great variety of catalytic activities available, ease of genetic manipulation and regular supply due to absence of seasonal fluctuations (Hasan et al., 2006). Although several bacterial and fungal lipase producers have been isolated and studied, filamentous fungi are recognized as the best lipase producers and currently the preferred sources due to their ability to produce copious amounts of extracellular lipases, the low cost of extraction, thermal and pH stability as well as activity in organic solvents (Heravi et al., 2008). In fact, several published reports indicate that the major fungal producers of commercial lipases belong to the genera Aspergillus, Mucor and Rhizopus (Carvalho et al., 2005; Koblitz and Pastore, 2006).
Although, currently, most of the industrial enzymes are produced by submerged fermentations which allow greater control of the culture conditions than solid state fermentation (Cheetham, 1995), the cost of production has remained high because more than 40% of the production cost is estimated to be due to the culture media alone (Wu et al., 2006). Presently, interests are growing in the use of cheap, sustainable and easily available substrates as media for microbial cultivation because enzyme production using inexpensive raw materials could make industrial enzymatic processes competitive with the chemical ones (Miranda et al., 1999). Palm oil mill effluent (POME) has potential for use as substrate for microbial lipase production because of its high mineral and oil/grease contents (Ma, 2000). It is the final liquid discharge after extracting the oil from the mashed fresh fruit bunch (Bek-Nielsen et al., 1999). POME is generated in large amounts throughout the year particularly in palm oil producing countries. It is estimated that for each ton of crude palm oil that is produced, 5-7.5 tons of water are required and more than 50% of this water ends up as POME (Ahmad et al., 2003). POME is a highly polluting waste water with capacity for considerable ecological damage when discharged untreated (Ma, 2000). Thus its conversion to value added products could save money as well as solve environmental problems.
Several studies indicate that POME is an excellent medium for the cultivation of fungi for the production of useful metabolites. Conversion of POME to several products by many filamentous fungi has been documented (Wu et al., 2006, 2009; Salihu et al., 2011). However, the use of free cultures poses some difficulty due to limitations of substrate utilization, especially, in fermentations involving lipid-rich wastes. This often affects the productivity of the fermentation negatively. Also, in freely suspended cultures, the requirement of an elaborate biomass recovery or separation step is time-consuming and expensive in scaled-up processes. It is thought that immobilization would promote close substrate-microbe contact thereby increasing product yield as well as easy recovery of biosolids. Although the use of immobilized cultures has been applied in the production of several metabolites, the choice of a cheap and efficient immobilization support is imperative because the use of expensive carriers will significantly increase the cost of production. Luffa sponge is an efficient immobilization carrier that has been successfully used in numerous bioconversion studies (Roble et al., 2003). It is cheap, abundant and available almost at no cost. Unfortunately, its application in the immobilization of cultures for the production of lipases from POME has not been reported. The present study, therefore, examines a method for overcoming the challenge of low substrate utilization in submerged cultures by immobilization of A. terreus on Luffa sponge for simultaneous treatment and production of microbial extracellular lipase from POME.
MATERIALS AND METHODS
Palm oil mill effluent (POME): POME used in the present study was obtained from a local palm oil processing mill at Nsukka, Enugu State, Nigeria and maintained at 4°C.
Microorganism and spore inoculum production: A strain of Aspergillus terreus IMI 282743 was obtained from the Department of Microbiology, University of Nigeria, Nsukka and screened for lipase production. The fungus was maintained at 4°C on Potato Dextose Agar (PDA) slants and sub cultured monthly. Mature spores were harvested by gently washing the surface of 5-7 day old subculture with 5 mL of sterile distilled water containing 0.1% Triton X-100. The spore suspension was subsequently centrifuged (Model H-201FR, Kokusan-Denki Co., Ltd., Shizuoka, Japan) at 4000 rpm for 10 min, washed and re-suspended in 5 mL distilled water. The resulting spore concentration was then adjusted to1.5×107 spores mL-1 using a Neubeur haemacytometer.
Immobilization of A. terreus: A. terreus IMI 282743 was immobilized according to the method described by Roble et al. (2003). Horizontally sliced portions of Luffa sponge (Luffa cylindrica) measuring 0.5-0.7 cm high and 3.5-5.0 cm in diameter were used. The dried outer covering of mature Luffa sponge was separated by hand before the sponge was cut. The slices were properly washed to remove adhering seeds, tissue fragments and dirt. They were then air dried before further drying in hot air oven (Eyela NDO-400, Rikakikai Co. Ltd., Tokyo, Japan) at 105°C for 2-3 h. The dried Luffa slices were finally pre-weighed in a digital balance (Model AX 200, Shimadzu Corp., Tokyo, Japan) before each portion was introduced into a 500 mL Erlenmeyer flask containing 90 mL of the diluted or undiluted POME followed by sterilization. After cooling, the different POME media were each inoculated with 10 mL of 1.5×107 spores mL-1 of spore suspension.
Effect of POME concentration on lipase activity: The effect of POME concentration on lipase activity was investigated by diluting the POME with distilled water to 25, 50, 75 or 100%. A 10 mL of the standardized inoculum suspension was inoculated into a 500 mL Erlenmeyer flask containing 90 mL of the diluted or undiluted POME. Incubation was carried out at 30°C for 7 days on a rotary shaker maintained at 210 rpm. Samples taken periodically were passed through Whatman No. 1 filter paper to remove mycelial fragments and other suspended solids. The cell free filtrates were evaluated for pH, lipase activity and COD.
Effect of nutrient enrichment on biomass growth, lipase activity and COD: The effect of nitrogen supplementation on biomass concentration, lipase activity and COD was studied by enriching 50% POME with a regime composed of different concentrations and combinations of (NH4)2SO4 and yeast extract according to a protocol developed from preliminary studies. Specifically, the nutrient regime was composed of (NH4)2SO4 (0.5%), Yeast Extract (0.1%), (NH4)2SO4 (0.5%)+Yeast Extract (0.1%) and (NH4)2SO4 (1.25%). The A. terreus was immobilized and cultivated as described by Ogbonna et al. (2001).
ANALYTICAL METHODS
Lipase assay: Lipase activity was determined spectrophotometrically according to a modification of the method reported by Pera et al. (2006). The substrate for the reaction was composed of solutions A and B. Solution A contained 40 mg of para-nitrophenylpalmitate (pNPP) dissolved in 12 mL of 96% isopropanol while solution B contained 0.1 g of gum Arabic and 0.4 mL of triton X-100 dissolved in 90 mL of water. The substrate solution was prepared by adding 1 mL of solution A to 19 mL of solution B drop wise with constant stirring to obtain an emulsion that remained stable for 2 h. The assay mixture contained 1 mL of the substrate, 0.8 mL of buffer (KH2PO4-NaOH, pH 7, 0.2 M) and 0.2 mL of the crude enzyme (the filtrate). The volume was made up to 3 mL with distilled water and incubated at 40°C for 30 min in a water bath (Personal-II, Taitec Corp., Saitama-ken, Japan). Enzyme activity was stopped by adding 0.2 mL of isopropanol. The absorbance was measured at 410 nm against substrate free blank using a spectrophotometer (Model UV- 1200, Shimadzu Corp., Tokyo, Japan). The standard graph was prepared by using para-nitrophenol at concentrations of 0-10 μM. One lipase unit (U) was defined as the amount of enzyme that liberated 1 μmol p-nitrophenol per minute under the assay conditions.
Analysis of POME: pH, Oil and Grease (O and G), Total Solids (TS) and Total Suspended Solids (TSS) were determined according to Standard Methods (APHA, 2005). Chemical Oxygen Demand (COD), Phosphorus (P), Ammonia-Nitrogen (NH3-N), Total Nitrogen (N) and Nitrates (-NO3) were measured by the Hach’s Spectrophotometic method (DR/4000, Hach Co., Ltd., Tokyo). Carbohydrate content was analyzed by the Phenol-Sulphuric acid method (DuBois et al., 1956) while Soluble Protein was measured by Bradford’s method (Bradford, 1976). Reducing sugar (Glucose) was measured by the Mutarotase/Glucose Oxidase assay using Glucose C2 Kit (Wako Pure Chemical Ind., Osaka) while Phenol was estimated by the Phenol test Kit (Wako Pure Chemical Ind., Osaka). Moisture content was determined by weight loss in 25 mL POME portions after drying in a hot air oven at 105°C for 24 h.
Determination of biomass concentration: Biomass concentration was measured at the end of each phase of fermentation. The Luffa sponge containing the immobilized mycelia was removed by a pair of sterile forceps, washed gently with distilled water and placed in the oven to dry at 105°C for 24 h. They were cooled in a desiccator and re-weighed. The differences in weight between the dried and pre-weighed Luffa and the weight after fermentation represent the biomass concentration in g/culture volume (100 mL).
Statistical analysis: The results presented are averages of triplicate determinations and were statistically analyzed by the standard error of means (SEM) and the two-way analysis of variance (ANOVA).
RESULTS
Effect of POME concentration on lipase production by immobilized A. terreus: Lipase activity was undetected in the 100% POME for nearly 48 h before it gradually rose to a peak concentration of 1.25 U mL-1 after 96 h (Fig. 1). Thereafter, lipolytic activity stabilized till the end of fermentation. In the 75% POME, low lipase concentration was detected after 24 h but an activity peak of 2.66 U mL-1 was reached after 72 h of fermentation. The 50% POME cultures promoted the highest lipase activity which rose steadily to a peak concentration of 5.14 U mL-1 after 7 days.
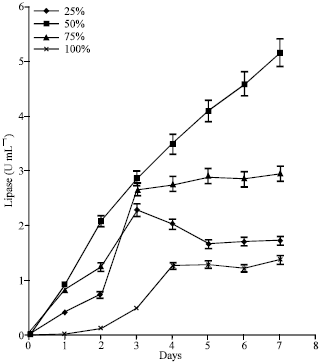 | |
| Fig. 1: | Effect of POME concentrations on lipase production by immobilized cultures of A. terreus |
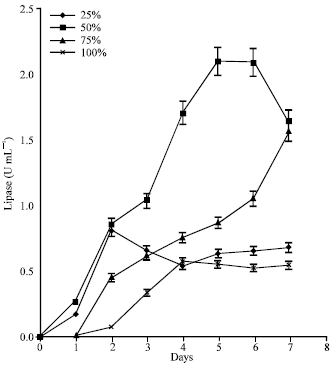 | |
| Fig. 2: | Effect of POME concentrations on lipase production by free cultures of A. terreus |
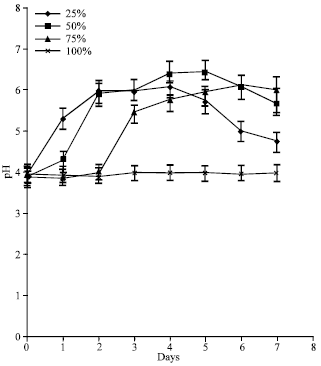 | |
| Fig. 3: | pH profiles of different POME concentrations during lipase production by A. terreus |
Lipolytic activity in the 25% POME increased almost linearly to 2.28 U mL-1 within the first 72 h. Thereafter, activity declined to 1.65 U mL-1 at the end of the experiment.
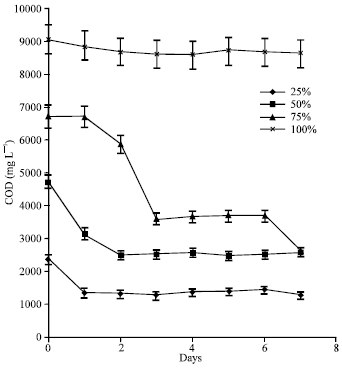 | |
| Fig. 4: | COD kinetics of the different POME concentrations during lipase production by immobilized cultures of A. terreus |
In the POME batches containing free cultures (i.e., without immobilization) lipase production started after 48 h of fermentation in the 100% POME but rose to an activity peak of 0.58 U mL-1 after 4 days (Fig. 2). In the 75% POME, activity was observed after 24 h lag although lipase concentration rose steadily to 1.57 U mL-1 after the 7th day of fermentation. Cultures from the 50% POME expressed the highest lipolytic concentration of 2.10 U mL-1 after 5 days. The lipase activity was stable from the 5-6th day before it finally declined. In the 25% POME cultures, activity rose sharply to a peak of 0.82 U mL-1 after 48 h but slowly dropped to a constant value after 96 h.
Effect of POME concentration on pH changes: The pH of the 100% POME was constant all through the course of fermentation (Fig. 3). In the 75% POME however, following a 48 h lag period, pH of the medium increased from 4-6.01 before subsequently declining as fermentation progressed. The pH of the 50% POME gradually increased to 6.5 within 120 h of cultivation before it declined to 5.7. In the 25% POME, pH rose sharply to peak values after 48 h and remained stable for 2 days before it gradually dropped.
Effect of POME concentration on COD kinetics: COD of the 100% POME did not change significantly during the cultivation (Fig. 4).
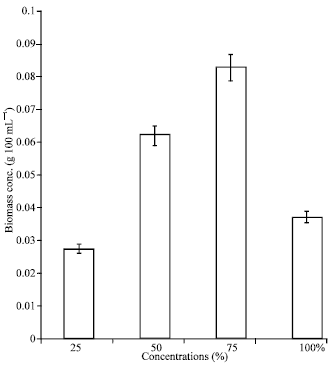 | |
| Fig. 5: | Effect of POME concentrations on biomass of immobilized cultures of A. terreus during lipase production |
However, following 24 h initial lag period, a 60.7% reduction in COD was achieved in the 75% POME while in the 50 and 25% POME, COD decreased by 45.3 and 44.9%, respectively.
Effect of POME concentrations on biomass production: The biomass concentration in the 100% POME was 0.037 g 100 mL-1 culture volume at the end of fermentation (Fig. 5). However, cell growth was most favored in the 75% POME, resulting in a final concentration of 0.083 g 100 mL-1. Biomass from the 50 and 25% POME were 0.062 and 0.027 g 100 mL-1, respectively. Biomass obtained in the 75% POME was two times higher than the concentration from the undiluted POME. The present finding indicates that POME dilution was favorable to biomass production.
Effect of nutrient enrichment on cell growth, lipase production and COD reduction: The biomass concentration increased as a result of the addition of nutrients to the 50% POME (Fig. 6). Cell growth was higher (p<0.05) in the enriched cultures than in the non-enriched media. A biomass concentration of 0.37 g 100 mL-1 was obtained from the 50% POME enriched with yeast extract (0.1%). This represents a 6-fold increase over the concentration (0.062 g L-1) obtained from the non-enriched 50% POME.
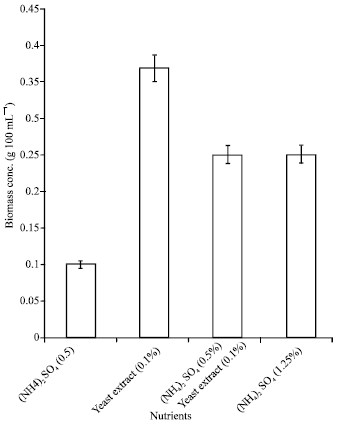 | |
| Fig. 6: | Effect of nutrient enrichment on the biomass of immobilized cultures of A. terreus cultivated in 50% POME concentration during lipase production |
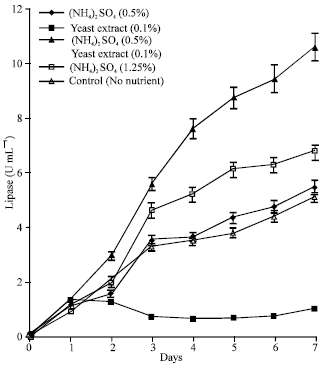 | |
| Fig. 7: | Effect of nutrient enrichment on lipase production by immobilized cultures of A. terreus |
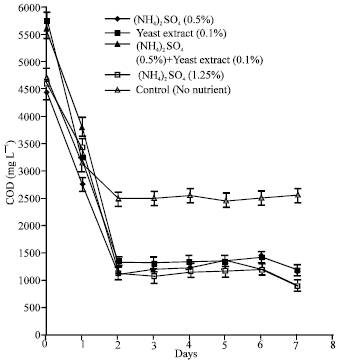 | |
| Fig. 8: | Effect of nutrient enrichment on the COD of 50% POME concentration during lipase production by immobilized cultures of A. terreus |
The biomass concentration was lowest (0.1 g 100 mL-1) in the medium supplemented with (NH4)2SO4 (0.5%) but in the media containing (NH4)2SO4 (0.5%)+Yeast Extract (0.1%) and (NH4)2SO4 (1.25%), comparable biomass concentration of 0.25g 100 mL-1 was obtained.
Addition of a nutrient regime composed of different concentrations and combinations of ammonium sulphate and yeast extract to 50% POME had variable effects on lipase production (Fig. 7). Lipolytic activity was highest in the medium supplemented with (NH4)2SO4 (0.5%)+yeast extract (0.1%). In this case the lipase activity rose steadily to a peak value of 10.6 U mL-1 by the 7th day while in the medium containing (NH4)2SO4 (1.25%), the activity increased to 6. 8 U mL-1 within the same period. In the batch enriched with (NH4)2SO4 (0.5%), the lipase activity was comparable (5.46 U mL-1) to the data obtained from the non-enriched 50% POME cultures (5.14 U mL-1). However, the lipolytic activity was negatively affected by Yeast Extract (0.1%) supplementation. The maximum lipase activity of 1.30 U mL-1 was reached after 24 h and dropped shortly afterwards to 0.71 U mL-1.
The results also showed that the nutrient enrichment significantly (p<0.05) promoted COD reduction in all the 50% POME batches tested (Fig. 8). In the medium containing (NH4)2SO4 (0.5%), 80.0% COD loss was obtained while in the batch enriched with Yeast Extract (0.1%), COD depreciated by 79.2%. COD in the (NH4)2SO4 (0.5%)+Yeast Extract (0.1%) and (NH4)2SO4 (1.25%) enrichments dropped by 78.8 and 80.5%, respectively after 7 days of fermentation. However, in the control 50% POME (without enrichment), the reduction in COD was 45.3%.
DISCUSSION
Effect of POME concentration on lipase production: Aspergillus terreus grew in all the different POME concentrations tested but lipolytic activity was highest in the 50% POME. In the POME batches containing immobilized cultures (Fig. 1), lipase activity was significantly (p<0.05) higher than values obtained from corresponding batches containing free cultures (Fig. 2). The highest lipase activity of 5.14 U mL-1 was achieved but in the non-immobilized cultures, peak lipase activity of 2.10 U mL-1 was obtained after 120 h. Comparable lipase activity was reported in Candida cylindraceae (ATCC 14830) suspension cultures using POME as the medium (Salihu et al., 2011). The low lipase activity reported in such cultures might be due to poor contact between the organism and the oil components of the medium. Materials intended for use as substrates for microbial fermentation should not only be present in sufficient concentrations but must also be available in useable forms. The physical nature of oils imposes severe restrictions on the ease with which they are available for microbial utilization. They are usually difficult to partition into the aqueous phase and when agitations are conducted at low temperatures, they often aggregate into clumps and separate out of the medium. Immobilization was found to be effective in improving the efficiency of substrate utilization and productivity of the fermentation process. In the present study, lipolytic activity increased 2.5 fold (5.12 U mL-1) in the 50% POME cultures following immobilization. Luffa sponge has been reported to be an efficient carrier for both filamentous fungi and flocculating yeasts (Ogbonna et al., 2001). It has been successfully used as immobilization carrier for the bioconversion of sugar beet juice and raw cassava starch to ethanol (Ogbonna et al., 2001; Roble et al., 2003). They are cheap and abundant in tropical countries where they grow as weeds in cultivated lawns. The matured pods are usually harvested and used as sponge for washing bodies, items and utensils after the dry epicarpic coverings have been removed. In this study, the Luffa was found to entrap oil by the help of the lattice structure and the oil-bed formed as a result provided an attachment support for the mould (Fig. 9).
 | |
| Fig. 9: | A 250 mL Erlenmeyer flask containing A. terreus immobilized on Luffa sponge after fermentation in POME for lipase production |
This promoted substrate contact, enhanced cell growth and COD reduction. In particular, the lipase activity was found to increase up to a maximum after which it decreased. This agrees with report by other authors (Lopes et al., 2008) and might be caused by the depletion of nutrients, accumulation of toxic end products of metabolism or other changes in medium composition. POME dilutions also affected lipase activity. Results in Fig. 1 indicate the need for dilution. Lipase activity was low in the undiluted (100%) POME but dilution led to increase in lipase and biomass production as well as reduction in COD. In the 100% POME, the major medium components such as oil, carbohydrate and protein did not occur in highly concentrated amounts (Table 1), thus the observed inhibition of microbial activity might have been due to other components such as antioxidants, phenolic acids and flavonoids. The presence of these compounds in POME has been confirmed (Lin et al., 2005; Uzel et al., 2005). The 25% POME concentration was found to be a poor cultivation medium because very low lipase and biomass concentration were obtained after fermentation perhaps due to over dilution which resulted in the depletion of key nutrients and minerals needed by the organism.
| Table 1: | Characteristics of POME used in the study |
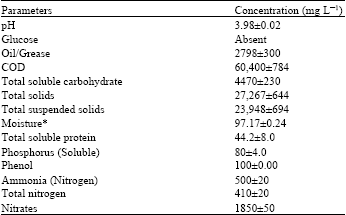 | |
| *(%) | |
Effect of POME concentration on pH kinetics during lipase production: The pH of the 100% POME was unchanged throughout the study maybe due to very low microbial activity in the cultures (Fig. 3). Preliminary studies indicated that pH 4 was optimal for lipase production by this organism. Since the POME had comparable pH value, no effort was made to adjust pH during experiment. Changes in pH profiles are known to reflect changes in metabolism during fermentation (Alonso et al., 2005). The absence of such change in the 100% POME might be due to the presence of certain inhibitory compounds in the medium. In the 75% POME, the organism recovered after a 48 h lag during which time, it adapted successfully to conditions in the medium. In the other lower POME concentrations, pH rose up to 6.4 due to utilization of organic acids and production of alkaline compounds (Abd Rahman et al., 2005) by the culture. Although pH 4 promoted the highest lipase activity, it was not the pH best suited for biomass production. Several studies have indicated that conditions which favor biomass growth are often different from those at which the highest lipolytic activities are produced (Contesini et al., 2010).
Effect of POME concentration on COD : The COD of the undiluted POME was unchanged (Fig. 3) but substantial reduction in organic load occurred at the other POME concentrations. In the 75% POME, COD reduction was 60.7% while in the 50 and 25% POME, COD reduction of 45.3 and 44.9% were achieved. The rate of COD reduction was higher in the 75% POME than in the other concentrations probably due to over dilution of the 50 and 25% POME with consequent limitation of essential nutrients. As microorganisms utilize the oil/grease present in the POME, biomass is boosted as a result of uptake of mineral elements and lipid hydrolysis products. The eventual reduction of oil/grease from the medium results in the lowering of COD because oil is converted to soluble products like fatty acids and glycerol which are eliminated from the system by microbial nutrition. Thus, as biomass increased, the rate of nutrient uptake increased bringing about rapid reduction in organic load of the effluent (Asses et al., 2009).
Effect of POME concentrations on biomass production: Despite sufficient nutrient content, the low biomass concentration (0.037 g 100 mL-1) recorded in the 100% POME after fermentation might have resulted from the presence of inhibitory components. However, biomass from the 25% POME was lower (0.027 g 100 mL-1) than the value obtained with the undiluted POME due to overdilution which depleted key nutrients and mineral elements in the medium. The 75% POME had the highest (0.083 g 100 mL-1) cell concentration and this was followed closely by the 50% POME with 0.062 g 100 mL-1.
Effect of nutrient enrichment on biomass production, lipase activity and COD reduction: Biomass production increased as a result of the addition of nutrients to the 50% POME (Fig. 6) batches. Thus biomass concentration was significantly (p<0.05) higher in the enriched cultures than in the non-enriched media. The result showed that the medium supplemented with yeast extract (0.1%) resulted in the highest biomass concentration of 0.37 g 100 mL-1 but negatively affected lipase production (Fig. 7). Lipase production in fungi is known to be boosted in the presence of different inorganic and organic nitrogen compounds or their combinations. Such beneficial effect of nitrogen supplementation has been reported by several authors. Lipase activity was significantly favored by (NH4)2SO4 and NH4Cl in cultures of Candida cylindraceae (D’Annibale et al., 2006). For fungi of the genus Penicillium, higher lipase activity have been obtained with organic nitrogen sources or a combination of organic and inorganic sources such as peptone or yeast extract with ammonium sulphate (Lima et al., 2003) than with ammonium sulphate as sole nitrogen source. However, conditions which favor biomass production are often different from those at which the highest lipolytic activities are produced (Contesini et al., 2010). In the present study (NH4)2SO4 (0.5%) promoted higher lipase activity than yeast extract 0.1% but a combination of both compounds i.e., NH4)2SO4 (0.5%)+Yeast Extract (0.1%) resulted in lipase activity (10.6 U mL-1) which is higher than the sum of the lipase activities in media supplemented with either of the nitrogen source. It is suspected that some form of synergism may have been involved in the observed lipolytic activities. Despite the high lipolytic activity obtained in the (NH4)2SO4 (0.5%)+Yeast Extract (0.1%) enriched media, the recovered biomass (0.25 g 100 mL-1) was significantly (p<0.05) lower than the concentration obtained from Yeast Extract (0.1%) supplemented medium (0.37 g 100 mL-1) and only comparable to cell growth obtained from the (NH4)2SO4 (1.25%) amended medium. Peak lipolytic activity in the medium containing (NH4)2SO4 (1.25%) was 6.8 U mL-1. Also, the high lipase activity recorded in the medium enriched with (NH4)2SO4 (0.5%) (5.46 U mL-1) did not correspond to the very low biomass (0.1 g mL-1) obtained from the system. The concentration of yeast extract added to the media was chosen after preliminary studies and thoughtful consideration of its possible impact on the organic load of the waste water. It is deducible from Fig. 7 that lipase activity from the medium enriched with (NH4)2SO4 (0.5%) was not statistically different (p>0.05) from the activity obtained from the non-enriched control treatment (5.12 U mL-1). However, while the COD in the former decreased by 80% (Fig. 8), COD in the control batch decreased by only 45.3%. Therefore, although (NH4)2 SO4 (0.5%) did not significantly improve lipase production; there was sufficient increase in biomass (Fig. 6) which conversely resulted in substantial reduction in the COD (Asses et al., 2009). Several workers such as Jin et al. (1999) had reported that nitrogen and phosphorus supplementation had beneficial effect on biomass production in cultures grown on agro-industrial effluents. The effect of the different nutrient enrichments show that biomass (Fig. 6) was promoted to different concentrations after fermentation. Incidentally, the observed variations in biomass did not show a graded activity on COD reduction (Fig. 8). For instance, although both concentrations of (NH4)2SO4 (i.e., 0.5 and 1.25%) had significantly different biomass concentrations, the level of COD reduction attained by both were basically the same. One reason could be that COD reduction did not depend on the final biomass concentration but on the progressive activity of an actively metabolizing microbiota. This is obvious from the result (Fig. 8) which showed that the most significant reduction in COD occurred within 48 h of fermentation. Finally, the COD of the different POME enrichments were significantly (p<0.05) lower than data obtained from the control treatment and meets the Malaysian permissible discharge characteristic of 1000 mg L-1 for effluent waste waters.
Analysis of POME used in the study: Principal biophysical properties of POME used in the present study are summarized in Table 1. The factors were found to vary with climate, palm variety as well as the efficiency of oil extraction. The mean oil/grease concentration and COD were 2798 and 60,400 mg L-1, respectively. These values agreed with Bhatia et al. (2007) and Lam and Lee ( 2011) but contrasts with Oswal et al. (2002). The concentration of total soluble carbohydrate (4470 mg L-1), suspended (23,948 mg L-1) and total solids (27,267 mg L-1) are related to previous reports by Ho et al. (1984) and Bhatia et al. (2007). The total soluble protein content of the waste water was found to be abysmally low (44.2 mg L-1). Nitrogen is originally present in POME in organic (protein) form but as time progresses, the organic compound gradually degrades to ammoniacal nitrogen (Chow, 1991) thereby diminishing the initial protein concentration. Glucose was not detected, contrary to a previous study by Wu et al. (2009) but the presence of phenol in POME agreed with Lin et al, (2005) and Uzel et al. (2005).
CONCLUSION
The use of immobilized cultures in the present study resulted in higher lipase activity and low COD content and showed potential for significant treatment cost savings than the free cultures. When free cells are used for lipase production or waste water treatment, biomass must first be separated by filtration or centrifugation before discharge otherwise cell lysis could occur resulting in the release of organic materials back into the environment. Also, the application of either or both separation methods might be extremely expensive on a large scale but the use of Luffa sponge as an immobilization carrier offers a cheap, efficient and sustainable method of achieving biosolid removal and promotes COD reduction and lipase production by improving microbe-substrate contact. Supplementation of POME with a combination of inorganic and organic nitrogen [i.e., (NH4)2SO4 (0.5%)+Yeast Extract (0.1%)] was needed for higher lipase activity and reduction in effluent COD to environmentally acceptable levels. A technically feasible integrated approach for simultaneous lipase production and POME treatment could be developed around this method.
ACKNOWLDGMENTS
Funding for this work was provided by the Tertiary Education Trust Fund (TETFUND) of the Federal Republic of Nigeria to Charles O. Nwuche. CON is grateful to Mr. Takashi Ishihara for his assistance during the author’s tenure as visiting foreign research fellow to the University of Tsukuba, Japan. The Microbiology Department, University of Nigeria is appreciated for the gift of the typed culture used in the study.
REFERENCES
- Abd Rahman, R.N.Z.R., L.P. Geok, M. Basri and A.B. Salleh, 2005. An organic solvent-tolerant protease from Pseudomonas aeruginosa strain K: Nutritional factors affecting protease production. Enzyme Microbial Technol., 36: 749-757.
CrossRefDirect Link - Alonso, F.O.M., E.B.L. Oliveira, G.M. Dellamora-Otiz and F.V. Perira-Meirelles, 2005. Improvement of lipase production at different stirring speeds and oxygen levels. Braz. J. Chem. Eng., 22: 9-18.
Direct Link - Asses, N., L. Ayed, H. Bouallagui, I. Ben-Rejeb, M. Gargouri and M. Hamdi, 2009. Use of Geotrichum candidum for olive mill wastewater treatment in submerged and static culture. Bioresour. Technol., 100: 2182-2188.
CrossRef - Babu, I.S. and G.H. Rao, 2007. Optimization of process parameters for the production of lipase in submerged fermentation by Yarrowia lipolytica NCIM 3589. Res. J. Microbiol., 2: 88-93.
CrossRefDirect Link - Bhatia, S., Z. Othman and A.L. Ahmad, 2007. Pretreatment of Palm Oil Mill Effluent (POME) using Moringa oleifera seeds as natural coagulant. J. Hazard. Mater., 145: 120-126.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Carvalho, P.D.O., F.J. Contesini, R. Bizaco and G.A. Macedo, 2005. Kinetic properties and enantioselectivity of the lipases produced by four Aspergillus species. Food Biotechnol., 19: 183-192.
CrossRef - Contesini, F.J., D.B. Lopes, G.A. Macedo, M.D.G. Nascimento and P.D.O. Carvalho, 2010. Aspergillus sp. lipase: Potential biocatalyst for industrial use. J. Mol. Catal. B: Enzym., 67: 163-171.
CrossRef - D'Annibale, A., G.G. Sermanni, F. Federici and M. Petruccioli, 2006. Olive-mill wastewaters: A promising substrate for microbial lipase production. Bioresour. Technol., 97: 1828-1833.
CrossRef - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Gupta, R., N. Gupta and P. Rathi, 2004. Bacterial lipases: An overview of production, purification and biochemical properties. Applied Microbiol. Biotechnol., 64: 763-781.
CrossRefDirect Link - Hasan, F., A.A. Shah and A. Hameed, 2006. Industrial applications of microbial lipases. Enzyme Microb. Technol., 39: 235-251.
CrossRefDirect Link - Heravi, K.M., F. Eftekhar, B. Yakhchali and F. Tabandeh, 2008. Isolation and identification of a lipase producing Bacillus sp. from soil. Pak. J. Biol. Sci., 11: 740-745.
CrossRefPubMedDirect Link - Ho, C.C., Y.K. Tan and C.W. Wang, 1984. The distribution of chemical constituents between the soluble and the particulate fractions of palm oil mill effluent and its significance on its utilisation/treatment. Agric. Wastes, 11: 61-71.
CrossRef - Houde, A., A. Kademi and D. Leblanc, 2004. Lipases and their industrial applications: An overview. Applied Biochem. Biotechnol., 118: 155-170.
CrossRefPubMedDirect Link - Jin, B., H.J. van Leeuwen, B. Patel, H.W. Doelle and Q. Yu, 1999. Production of fungal protein and glucoamylase by Rhizopus oligosporus from starch processing wastewater. Process Biochem., 34: 59-65.
CrossRef - Lam, M.K. and K.T. Lee, 2011. Renewable and sustainable bioenergies production from Palm Oil Mill Effluent (POME): Win-win strategies toward better environmental protection. Biotechnol. Adv., 29: 124-141.
CrossRefPubMedDirect Link - Lima, V.M.G., N. Krieger, M.I.M. Sarquis, D.A. Mitchell, L.P. Ramos and J.D. Fontana, 2003. Effect of nitrogen and carbon sources on lipase production by Penicillium aurantiogriseum. Food Technol. Biotechnol., 41: 105-110.
Direct Link - Lin, Y.T., R.G. Labbe and K. Shetty, 2005. Inhibition of Vibrio parahaemolyticus in seafood systems using oregano and cranberry phytochemical synergies and lactic acid. Innov. Food Sci. Emerg. Tecnol., 6: 453-458.
CrossRef - Lopes, M., N. Gomes, C. Goncalves, M.A.Z. Coelho, M. Mota and I. Belo, 2008. Yarrowia lipolytica lipase production enhanced by increased air pressure. Lett. Appl. Microbiol., 46: 255-260.
CrossRef - Melo, L.L.M.M., G.M. Pastore and G.A. Macedo, 2005. Optimized synthesis of citronellyl flavour esters using free and immobilized lipase from Rhizopus sp. Biochemistry, 40: 3181-3185.
Direct Link - Miranda, O.A., A.A. Salgueiro, M.C.B. Pimentel, L.J.L. Filho, E.H.M. Melo and N. Duran, 1999. Lipase production by Brazilian strain of Penicillium citrinu using an industrial residue. Bioresour. Technol., 69: 145-147.
Direct Link - Ogbonna, J.C., H. Mashima and H. Tanaka, 2001. Scale up of fuel ethanol production from sugar beet juice using loofa sponge immobilized bioreactor. Bioresour. Technol., 76: 1-8.
CrossRef - Oswal, N., P.M. Sarma, S.S. Zinjarde and A. Pant, 2002. Palm oil mill effluent treatment by a tropical marine yeast. Bioresour. Technol., 85: 35-37.
CrossRefPubMedDirect Link - Pera, L.M., C.M. Romero, M.D. Baigori and G.R. Castro, 2006. Catalytic properties of lipase extracts from Aspergillus niger. Food Technol. Biotechnol., 44: 247-252.
Direct Link - Prazeres, J.N., J.A.B. Cruz and G.M. Pastore, 2006. Characterization of alkaline lipase from Fusarium oxysporum and the effect of different surfactants and detergents on the enzyme activity. Braz. J. Microbiol., 37: 505-509.
CrossRef - Roble, N., J. Ogbonna and H. Tanaka, 2003. A novel circulating loop bioreactor with cells immobilized in Loofa (Luffa cylindrica) sponge for the bioconversion of raw cassava starch to ethanol. Applied Microbiol. Biotechnol., 60: 671-678.
CrossRef - Salihu, A., M.Z. Alam, I.M. AbdulKarim and H.M. Salleh, 2011. Optimization of lipase production by Candida cylindracea in palm oil mill effluent based medium using statistical experimental design. J. Mol. Catal. B: Enzymatic, 69: 66-73.
CrossRefDirect Link - Shan, T., T. Wu, Y. Reng and Y. Wang, 2009. Breed difference and regulation of the porcine adipose triglyceride lipase and hormone sensitive lipase by TNFalpha. Anim. Genet., 40: 863-870.
PubMedDirect Link - Uzel, A., K. Sorkun, O. Oncag, D. Cogulu, O. Gencay and B. Salih, 2005. Chemical compositions and antimicrobial activities of four different Anatolian propolis samples. Microbiol. Res., 160: 189-195.
CrossRefDirect Link - Wu, T.Y., A.W. Mohammad, J.M. Jahim and N. Anuar, 2009. Optimized reuse and bioconversion from retentate of pre-filtered palm oil mill effluent (POME) into microbial protease by Aspergillus terreus using response surface methodology. J. Chem. Technol. Biotechnol., 84: 1390-1396.
Direct Link - Wu, T.Y., A.W. Mohammad, J.M. Jahim and N. Anuar, 2006. Investigations on protease production by a wild-type Aspergillus terreus strain using diluted retentate of pre-filtered palm oil mill effluent (POME) as substrate. Enzyme Microb. Technol., 39: 1223-1229.
Direct Link - Ahmad, A.L., S. Ismail, N. Ibrahim and S. Bhatia, 2003. Removal of suspended solids and residual oil from palm oil mill effluent. J. Chem. Technol. Biotechnol., 78: 971-978.
CrossRef