
A. Togola
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
F.E. Nwilene
Africa Rice Center (AfricaRice), P.M.B. 5320, Ibadan, Oyo State, Nigeria
E.A. Kotoklo
Laboratory of Applied Entomology, Faculty of Science, University of Lome, B.P. 1515, Lome, Togo, Benin
K. Amevoin
Laboratory of Applied Entomology, Faculty of Science, University of Lome, B.P. 1515, Lome, Togo, Benin
I.A. Glitho
Laboratory of Applied Entomology, Faculty of Science, University of Lome, B.P. 1515, Lome, Togo, Benin
O.E. Oyetunji
Africa Rice Center (AfricaRice), P.M.B. 5320, Ibadan, Oyo State, Nigeria
A. Niang
Africa Rice Center (AfricaRice), 01 B.P. 2031, Cotonou, Benin
Journal of Applied Sciences
Year: 2012 | Volume: 12 | Issue: 7 | Page No.: 675-680







ABSTRACT
In tropical Africa, termite damage to rice crops leads to important losses of farmers’ income. The present study aimed to evaluate the effect of rice varieties and cultural practices on termite populations and damage in the field. For this purpose, 10 rice varieties (seven NERICA, one Oryza glaberrima and two O. sativa) were cultivated in rainfed upland at Niaouli (Benin) under four cultivation practices. Termite populations and damage were evaluated during tillering, heading and maturation stages. NERICA 6, CG14 (the African O. glaberrima, parent of the NERICA), NERICA 10, NERICA 2 and NERICA 3 were the least infested and suffered least damage, while NERICA 4 and LA23 were the most susceptible. Among the cultural practices, use of Metarhizium anisopliae and regular weeding resulted in lower termite populations and less damage than other practices. Rice was most susceptible to termite attack in the period from heading to maturation. Results of this study should help rice farmers in their choice of tolerant varieties and adapted cultural practices as effective alternative measures to control termites and save rice production in the rainfed upland ecology.
PDF Abstract XML References Citation
Received: October 30, 2011;
Accepted: March 28, 2012;
Published: June 19, 2012
How to cite this article
A. Togola, F.E. Nwilene, E.A. Kotoklo, K. Amevoin, I.A. Glitho, O.E. Oyetunji and A. Niang, 2012. Effect of Upland Rice Varieties and Cultural Practices on Termite Populations and Damage in the Field. Journal of Applied Sciences, 12: 675-680.
DOI: 10.3923/jas.2012.675.680
URL: https://scialert.net/abstract/?doi=jas.2012.675.680
DOI: 10.3923/jas.2012.675.680
URL: https://scialert.net/abstract/?doi=jas.2012.675.680
INTRODUCTION
Rice (Oryza spp.) is a staple crop in Africa, where it has been cultivated for more than 3000 years (ARC, 2005). On this continent, rice production is increasing because of its role in both human nutrition and national economies. However, the local production is insufficient to achieve food security and the demand for rice imports continues to rise.
Indeed, many constraints are responsible of low rice production in Africa. Apart from the irregular climatic condition and low soil fertility, insects have a considerable impact on rice productivity, causing yield losses of about 30% in upland rice and up to 100% in lowland rice (Umeh et al., 2001; Togola et al., 2010). They are regularly present in rice fields damaging rice plants and compromising farmers’ efforts.
Termites are one group of soil-living arthropods that are particularly harmful to rainfed upland rice and to most dryland crops (Wood and Cowie, 1988). Their role in the reduction of rice yields is very significant. In the field, rice plants are attacked mainly on their root systems. Later, the stems are reached through galleries built from the roots that are then filled with soil (Nwilene et al., 2008). Infested seedlings dry out or become predisposed to attack from rodents, ants, fungi or bacteria (Wood and Cowie, 1988; Nwilene et al., 2008). Termite damage in the field reduces seedling density and leads to substantial loss of rice productivity (Heinrichs and Barrion, 2004; Nwilene et al., 2008). Losses of 50-100% have been reported on many tropical crops (Wood and Pearce, 1991; Sekamatte et al., 2001; Kiwuso et al., 2004; Nwilene et al., 2008; Agunbiade et al., 2009). For instance, severe damage has been reported on yam, cassava (Atu, 1993), sugarcane (Sands, 1977), groundnut (Johnson and Gumel, 1981), sorghum (Logan et al., 1990) and maize (Wood et al., 1980). Termites also attack wild plants in natural savannah. They can feed on all material that contains cellulose.
Control of termites is based mainly on the use of chemicals which have negative effects on other living resources in the environment. It is, therefore, important to explore alternative solutions that are effective against termites and less polluting of the environment.
The present study aimed to test the effectiveness of some improved rice varieties and cultural practices as integrated crop management options against termites in rice fields. The study intends to contribute to the management of soil-borne pests with emphasis on termites which are a key issue for rice producers, especially those working in upland ecologies.
MATERIAL AND METHODS
The study was conducted during the 2009 cropping season (from July to October) at Niaouli, 80 km North of the district of Cotonou, Benin. The site is known to be a hot spot for termite infestation. It records an average annual rainfall of 1200 mm and average daily evapotranspiration of 3.5-3.7 mm. The monthly mean temperature is 21.5°C during the rainy season and 32°C during the dry season. The rainfall pattern is bimodal and comprises a main rainy season from March to July and a short rainy season from September to November. The location has an upland ecology with a ferralitic upland soil. The upper horizon (0-25 cm) is sandy whereas, the lower horizon (25-50 cm) is a clay-sandy soil. The vegetation is humid guinea savannah covered with monocotyledonous and dicotyledonous herbage with some trees.
Experimental design: The experimental design was a split plot with four cultural practices in the main plots: (1) weed-free (weeding at 21 and 42 days after sowing); (2) organic farming (using sawdust spread during the two weeding periods at the rate of 200 g m-2); (3) use of Metarhizium anisopliae (entomopathogenic fungus; 2 g mixed with 600 g of sawdust and spread on the whole plot during the two weeding periods) and (4) left weedy (the control treatment). Sawdust was collected from a local carpenter. Metarhizium fungus was obtained from laboratory culture. The main plots were 25x3 m with 3 m alley between plots. The sub-plots were the 10 rice varieties: seven NERICA varieties (1, 2, 3, 4, 6, 7 and 10), one Oryza glaberrima (CG14), two Oryza sativa (WAB56-104, the Asian parent of the NERICA and LAC23 which is resistant to stem borers. Varieties were sown in sub-plots of 1x3 m with 2 m alley between sub-plots (sub-plots were randomized within main plots). All treatments (cultural practices and rice varieties) were replicated three times.
Planting techniques: The test lines were sown at a hill spacing of 20x20 cm. Gaps were filled once at 7 days after emergence. Three seedlings were maintained on each hill. A basal dose of NPK (15-15-15) at 150 kg ha-1 was applied before sowing. Also a top-dressing of urea (50 kg ha-1) was applied twice during the weeding periods at 21 and 42 days after sowing (DAS).
Sampling of termite population and damage: The population sampling was carried out during three phenological stages of rice: (1) tillering (40 DAS), (2) heading (60 DAS) and (3) maturation (90 DAS). Sampling comprised locating five sampling points 0.5 m distant from each other along plot diagonals. Holes of 0.25x0.25x0.1 m (Benito et al., 2004) were made between hills. For each new sampling period a new hole was dug a few centimeters away from the previous sampling point. In addition, nine hills-three hills each of fresh plants, wilted plants and dry plants-were extracted at maturity to collect and count termites.
The collected soils were investigated in an entomology laboratory and termites species were identified using several identification guides (Sands, 1965; Harris, 1968; Pearce et al., 1992). Termite damage was also recorded during the three phenological stages. It consisted of counting dead and total plants in each rice plot in order to calculate the percentage of dead plants.
Data collection and statistical analysis: The main data collected concerned the numbers of termites per species in each plot and infested plants in each plot during the three observation periods.
The average termite population per plot and the average infested plant percentage were calculated. The data were analyzed by analysis of variance (ANOVA) with SAS 8.2 (SAS Institute, 2002). Significant differences between termite populations and between damage to rice varieties or cultural practices were compared by the Student-Newman-Keuls (SNK) test at 5% probability level.
RESULTS
Effect of rice varieties on termite populations and damage: The population of termites showed large variation among rice varieties and according to their phenological stages.
At tillering stage, the termite population was low on all rice varieties with no significant differences between them. At heading stage, although the mean populations were higher than those at tillering stage, there were again no significant differences among varieties. At maturity stage, the termite populations were high and significantly different among the rice varieties (p<0.001). NERICA 4 (31.1 individuals 0.06 m-2) was the most heavily infested. It was followed by NERICA 1, LAC23 and NERICA 7 that recorded 26.3, 25.4 and 22.4 individuals 0.06 m-2, respectively. During the same period, NERICA 6, NERICA 10 and CG 14 were least infested scoring 0.7, 4.3 and 4.8 individuals 0.06 m-2, respectively (Table 1).
| Table 1: | Termite population and damage on rice varieties through the phenological stages |
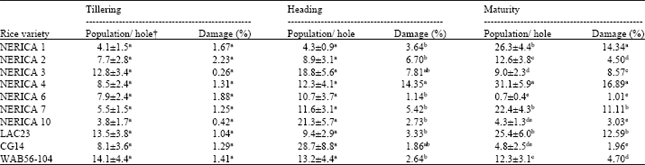 | |
| † Sampling holes were 0.25x0.25x0.1 m, Means followed by the same letter are not significantly different at the 5% level according to SNK test | |
The damage was very low during tillering stage but subsequent stages showed high damage levels with peaks of 14.35% at heading and 16.89% at maturity. NERICA 4, NERICA 1, LAC23 and NERICA 7 were most damaged varieties with 16.89, 14.34, 12.59 and 11.11% damage, respectively, at maturity. In contrast, NERICA 6 (1.01%), CG 14 (1.96%), NERICA 10 (3.03%) and NERICA 2 (4.5%) were highly tolerant to termite attack. The O. sativa parent of the NERICA (WAB 56-104), also showed high tolerance, with 4.7% of infested tillers. NERICA 1 was resistant to termite attack during tillering and heading but very susceptible at maturity when 14.34% tillers were infested. NERICA 3 showed a tolerance to termite attack at all stages (Table 1). There were highly significant differences (p<0.001) among the varieties in terms of damage at heading and maturity stages.
Effect of cultural practices on termite populations and damage: Termite populations and damage were both very low during the tillering stage in all plots. However, the plots with sawdust were significantly more infested (14.9 termites/hole) than the others. Conversely, the clean (weed-free) plots (3.7 individuals) drew a smaller termite population than the other treatments but the difference was not significant (Fig. 1). By heading stage, termite density had increased above that of tillering stage under all cultural practices. At this stage, the sawdust plots were highly infested with 21.4 termites/hole, while the three other treatments were not significantly different from one another. At maturity, the sawdust treatment remained most infested (20.3 individuals) whereas termite densities were moderate in the other treatments (Fig. 1).
Termite damage varied not only according to crop growing stages but also according to cultural practices (Fig. 2). Indeed, the damage level was low during tillering and high at heading under all cultural practices. At maturity, damage reached its highest level under all cultural practices except for the Metarhizium treatment. At that last stage, the various treatments showed significant differences: Metarhizium and weed-free treatments were the best practices scoring 4.47% and 7.27% infested tillers, respectively.
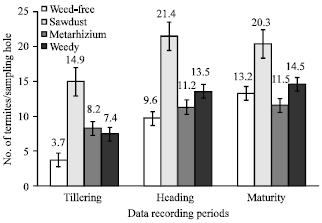 | |
| Fig. 1: | Population of termites according to cultural practice |
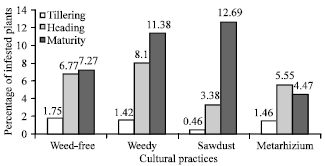 | |
| Fig. 2: | Percentage of infested plants according to cultural practice |
Effect of physical state of rice plant on termite abundance: At maturity stage, the physical state of the plant had a highly significant influence on the infestation of termites. Indeed, termite populations were double on dry plants (4.5 individuals/hill) compared to wilted plants (2.6 individuals) and five times those on fresh plants (0.9 individuals) (Fig. 3).
Termite population dynamics and damage during the cropping cycle: The regression lines of termite population and damage showed linear trends from tillering to maturation (Fig. 4).
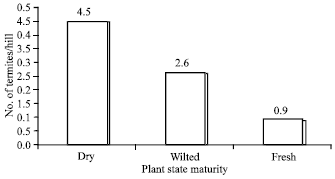 | |
| Fig. 3: | Effect of plant state on population of termites at maturity |
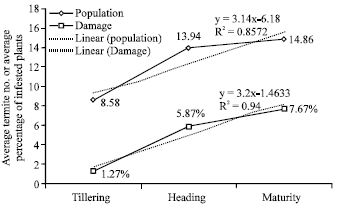 | |
| Fig. 4: | Evolution of termite population and damage from tillering to maturity stages |
The trend lines of the two parameters were parallel and showed positive coefficients of regression: R2 = 0.85 for the population and R2 = 0.94 for the damages.
DISCUSSION
This study showed that NERICA 6, CG14 and NERICA 3 recorded very low termite populations even in weedy and sawdust-treated plots and equally low termite damage. NERICA 6, CG14, NERICA 10 and NERICA 2 showed the lowest rates of infested tillers during rice crop phenological stages. NERICA 3 also showed a tolerance to termite attack. The tolerance of these NERICA varieties to termite attack could have been inherited from their O. glaberrima parent, CG14-the tolerance of this variety to African biotic constraints in general has been reported by WARDA (1992) as by Jones et al. (1997). Oryza glaberrima has been cultivated in Africa for more than 3000 years (Bidaux, 1978; Carpenter, 1978), where it has probably developed some mechanisms of resistance against various stresses (Ukwungwu et al., 1998). Thus, the NERICA varieties would have received their resistance genes from this parent. This research confirms the good adaptation of O. glaberrima and its descendants (NERICA) to African environmental conditions (ARC, 2005). In the same way, Heinrichs and Barrion (2004) demonstrated that termite attack was generally less severe on local varieties than imported ones.
NERICA 1 had very low termite infestation during the early vegetative stages (tillering and heading) but was heavily infested at maturity. This variety is aromatic and the aroma produced especially during the vegetative stages would repel termites during this period. But at maturity this variety was heavily infested probably because of the dryness of the stems and the diminution of the aroma.
The results demonstrated the tolerance of five NERICA varieties to termites-an element of the performance of NERICA against field insects in general (Bidaux, 1978; Arraudeau, 1992; WARDA, 1992; Nwilene et al., 2008). NERICA 4 and LAC23 were rather susceptible to termite attack.
Among cultural practices, the weed-free and Metarhizium plots were least infested and with moderate damage compared to weedy and sawdust plots which drew high termite populations and suffered high levels of damage. By preventing weed competition, the weed-free method would have reinforced plant vigor and increased the plants’ resistance to termite attack. The effectiveness of Metarhizium would be due to its entomopathogenic property that kills termites and reduces the populations (Kramm and West, 1982; Sun et al., 2002; Wright et al., 2005). Under weedy conditions the seedlings are stressed and less vigorous creating a favorable environment for termite infestation. Umeh et al. (2001) report on the role of crop residues in increasing termite populations in the field. Wood et al. (1980), Mora et al. (1996) and Sileshi et al. (2005) also found that wild grasses or crop residues left on the ground constitute important cellulose sources for termites. Our study revealed the presence of six species of termites in rice fields: Microtermes sp., Pseudacanthotermes militaris, Macrotermes bellicosus, Microcerotermes parvus, Amitermes evuncifer and Trinervitermes oeconomus.
Concerning the physical state of the plants during the maturation stage, most of the termite population was recorded on dry plants, followed by the wild plants probably because of the availability of cellulose in those plants. Termites were present in the rice field during the whole crop cycle but their populations and damage were economically important at maturity stage.
CONCLUSION
This study showed the influence of varieties, phenological stages, plant physical state and cultural practices on both termite populations and damage on rice in an upland ecology. It revealed the efficacy of various methods for controlling termites infestation, such as the use of NERICA 6, CG14, NERICA 10, NERICA 2 and NERICA 3 as tolerant varieties and the use of weeding and Metarhizium as cultural practices. The results of this study should help smallholder farmers to access information about effective technologies against termites that remain serious constraints especially for dryland crops.
REFERENCES
- Agunbiade, T.A., F.E. Nwilene, A. Onasanya, M. Semon, A. Togola, M. Tamo and O.O. Falola, 2009. Resistance status of upland NERICA rice varieties to termite damage in Northcentral Nigeria. J. Applied Sci., 9: 3864-3869.
CrossRefDirect Link - Atu, U.G., 1993. Cultural practices for the control of termite (Isoptera) damage to yams and cassava in south-eastern Nigeria. Int. J. Pest Manage., 39: 462-466.
CrossRef - Benito, N.P., M. Brossard, A. Pasini, M. de Fatima Guimaraes and B. Bobillier, 2004. Transformations of soil macroinvertebrate populations after native vegetation conversion to pasture cultivation (Brazilian Cerrado). Eur. J. Soil Biol., 40: 147-154.
CrossRef - Johnson, R.A. and M.H. Gumel, 1981. Termite damage and crop loss studies in Nigeria: The incidence of termite-scarified groundnut pods and resulting kernel contamination in field and market samples. Trop. Pest Manage., 27: 343-350.
CrossRef - Jones, M.P., M. Dingkuhn, G.K. Aluko and M. Semon, 1997. Interspecific Oryza sativa L. X O. Glaberrima steud. Progenies in upland rice improvement. Euphytica, 94: 237-246.
CrossRefDirect Link - Kramm, K.R. and D.F. West, 1982. Termite pathogens: Effects of ingested Metarhizium, beauveria and Gliocladium conidia on worker termites (Reticulitermes sp.). J. Inverteb. Pathol., 40: 7-11.
CrossRef - Logan, J.W.M., R.H. Cowie and T.G. Wood, 1990. Termite (Isoptera) control in agriculture and forestry by non-chemical methods: A review. Bull. Entomol. Res., 80: 309-330.
CrossRefDirect Link - Mora, P., C. Rouland and J. Renoux, 1996. Foraging, nesting and damage caused by Microtermes subhyalinus (Isoptera: Termitidae) in a sugarcane plantation in the Central African Republic. Bull. Entomol. Res., 86: 387-395.
CrossRefDirect Link - Nwilene, F.E., T.A. Agunbiade, M.A. Togola, O. Youm and O. Ajayi et al., 2008. Efficacy of traditional practices and botanicals for the control of termites on rice at Ikenne, Southwest Nigeria. Int. J. Trop. Insect Sci., 28: 37-44.
CrossRefDirect Link - Sekamatte, M.B., M.W.O. Latigo and A.R. Smith, 2001. The effect of maize stover used as mulch on termite damage to maize and activity of predatory ants. Afr. Crop Sci. J., 9: 411-419.
Direct Link - Sileshi, G., P.L. Mafongoya, F. Kwesiga and P. Nkunika, 2005. Termite damage to maize grown in agroforestry systems traditional fallows and monoculture on nitrogen-limited soils in eastern Zambia. Agric. Forest Entomol., 7: 61-69.
CrossRefDirect Link - Umeh, V.C., F. Waliyar, S. Traore, I.M. Chaibou, B. Omar and J. Detognon, 2001. Farmers opinions and influence of cultural practices on soil pest damage to groundnut in West Africa. Insect Sci. Applied, 21: 257-265.
Direct Link - Wood, T.G., R.A. Johnson and C.E. Ohiagu, 1980. Termite damage and crop loss studies in Nigeria: A review of termite (Isoptera) damage, loss in yield and termite Microtermes) abundance at Mokwa. Trop. Pest Manage., 26: 241-253.
CrossRefDirect Link - Wright, M.S., A.K. Raina and A.R. Lax, 2005. A strain of the fungus Metarhizium anisopliae for controlling subterranean termites. J. Econ. Entomol., 98: 1451-1458.
PubMedDirect Link