
Tooba Mirzapour
Institute of Bioscience, Universiti Putra Malaysia, Selangor, Malaysia
Tengku Azmi Bin Tengku Ibrahim
Institute of Bioscience, Universiti Putra Malaysia, Selangor, Malaysia
Mansoureh Movahedin
Department of Anatomical Sciences, Tarbiat Modares University, Tehran, Iran
Abd Wahid Haron
Department of Veterinary Clinical Studies, Universiti Putra Malaysia, Malaysia
Siti Shapor Siraj
Department of Biology, Universiti Putra Malaysia, Selangor, Malaysia
Morteza Koruji
Department of Anatomical Sciences, Faculty of Medicine, Iran University of Medical Science, Tehran, Iran
Mohamadreza Nowroozi
Department of Urology, Faculty of Medicine, Tehran University of Medical Science, Tehran, Iran
Shahram Rafieian
Department of Urology, Faculty of Medicine, Tehran University of Medical Science, Tehran, Iran
Journal of Applied Sciences
Year: 2011 | Volume: 11 | Issue: 1 | Page No.: 163-173







ABSTRACT
Research in developmental biology has led to the discovery of different types of stem cells (totipotent, pluripotent and multipotent stem cells) that can give rise to multiple tissue types. This review summarizes a description about the stem cell concept, different type of stem cells and their potential applications. The face of extraordinary advances in the prevention, diagnosis and treatment of human diseases, devastating illnesses such as heart disease, diabetes, cancer and diseases of the nervous system, continue to deprive people of health, independence and well-being has been reviewed in this study. Stem cell research leading to prospective therapies in reparative medicine has the potential to affect the lives of millions of people around the world and there is a good reason to be optimistic. The road towards the development of an effective cell-based therapy for widespread use is long and involves overcoming numerous technical, legislative, ethical and safety issues.
PDF Abstract XML References Citation
Received: June 19, 2010;
Accepted: August 08, 2010;
Published: November 10, 2010
How to cite this article
Tooba Mirzapour, Tengku Azmi Bin Tengku Ibrahim, Mansoureh Movahedin, Abd Wahid Haron, Siti Shapor Siraj, Morteza Koruji, Mohamadreza Nowroozi and Shahram Rafieian, 2011. Stem Cells Research and its Applications: A Review. Journal of Applied Sciences, 11: 163-173.
DOI: 10.3923/jas.2011.163.173
URL: https://scialert.net/abstract/?doi=jas.2011.163.173
DOI: 10.3923/jas.2011.163.173
URL: https://scialert.net/abstract/?doi=jas.2011.163.173
INTRODUCTION
Regeneration is a remarkable physiological process in which remaining tissues organize to reform a missing body part. Several invertebrates, such as planarian, flatworms and Hydra, regenerate tissues with speed and precision. From vertebrates, the salamanders, repair lost body parts through the dedifferentiation of specialized cells into new precursor cells. Stem cells can also be identified in plants in the root and shoot meristems (Bangso and Richard, 2004). However, the majority of higher vertebrates are incapable of any form of whole-organ regeneration, even though they had all the necessary instructions and machinery to generate the tissue during embryonic development (Andrews, 2002).
Most tissue regenerate events in mammals are dedifferentiation-independent events resulting from the activation of pre-existing stem cells or progenitor cells. These cells are the common denominator for nearly all types of regeneration (Bangso and Richard, 2004).
The canonical definition of a stem cell is a cell with the ability to divide indefinitely in culture and with the potential to give rise to mature specialized cell types (Alison et al., 2002). This style of cell division characteristic of stem cells is asymmetric. In fact, when a stem cell divides, the daughter cells can either enter a path leading to the formation of a differentiated specialized cell or self-renew to remain a stem cell, thereby ensuring that a pool of stem cells is constantly replenished in the adult organ. This mechanism is a necessary physiological mechanism for the maintenance of the cellular composition of tissues and organs in the body (Andrews, 2002; Bangso and Richard, 2004; Kanatsu-Shinohara et al., 2004).
The field of stem cells began with the study of teratocarcinomas in the 1960s. Teratocarcinomas are malignant germ cell tumors that form an undifferentiated EC (Embryonal Carcinoma cells) component and a differentiated component that can include all three germ layers. Mouse EC cell lines that could be stably propagated in vitro were established in the early 1970s. They could be cultured in sufficient quantities to perform experiments that would have been impossible with intact mammalian embryos (Kahan and Ephrussi, 1970).
Isolation of a Pluripotent cell line from early mouse embryos cultured in a medium conditioned by teratocarcinomas stem cells was done by Evans and Kaufman (1981) and Martin (1981). After that derivation of human embryonic stem cells (ESCs) in 1998 ignited an explosion of public interest in stem cells (Thomson et al., 1998). In turn, the recent derivation of mouse and human induced pluripotent stem cells depended on the prior studies on mouse and human ESCs. Both human ESCs and induced pluripotent stem cells can self-renew indefinitely in vitro while maintaining the ability to differentiate into advanced derivatives of all three germ layers, features very useful for understanding the differentiation and function of human tissues, for drug screen and toxicity testing and for cellular transplantation therapies (Yu et al., 2008). Clearly, stem cell research leading to prospective therapies in reparative medicine has the potential to affect the lives of millions of people around the world for the better and there is good reason to be optimistic.
TYPES OF STEM CELLS AND ITS APPLICATION
Stem cells can be classified as Totipotent, Pluripotent and Multipotent cells. Totipotency is the ability to form all cell types of the conceptus, including the entire fetus and placenta. These cells have unlimited capability; they can basically form the whole organism. Early mammalian embryos are clusters of totipotent cells.
Pluripotency is the ability to form several cell types of all three germ layers (ectoderm, mesoderm and endoderm) but not the whole organism. In theory, pluripotent stem cells have the ability to form all the 200 or so cell types in the body.
Multipotency is the ability of giving rise to a limited range of cells and tissues appropriate to their location.
There are four classes of pluripotent stem cells: (1) Embryonic stem cells, (2) Embryonic germ cells, (3) Multipotent germline stem cells or mGSCs, (4) Embryonic carcinoma cells and (5) The Multipotent adult progenitor cell from bone marrow.
Mammalian development starts from a single cell that can give rise to all cells required for a new life, but through subsequent differentiation events, developmental potential becomes increasingly restricted. As the one-cell embryo divides, it forms a morula, a mulberry-like cluster of undifferentiated cells. The first differentiation event occurs when the outer layer of cells of the morula differentiates to the trophectoderm, forming the blastocyst stage embryo. The cells inside the blastocyst (inner cell mass, or ICM) give rise to all cells of the adult body and some extraembryonic tissues, while the trophectoderm gives rise to the outer layer of the placenta (Andrews, 2002; Bangso and Richard, 2004; Yu et al., 2008).
Embryonic Stem (ES) cells, however, are derived from the isolated Inner Cell Masses (ICM) of mammalian. The continuous in vitro subculture and expansion of an isolated ICM on an embryonic fibroblast feeder layer (human or murine) leads to the development of an embryonic stem cell line. In nature, however, embryonic stem cells are ephemeral and present only in the ICM of blastocysts. These cells are destined to differentiate into tissues of the three primordial germ layers (ectoderm, mesoderm and endoderm) and finally form the adult organism (Bangso and Richard, 2004).
Totally, if early mouse embryos are transferred to extrauterine sites, such as the kidney or testis capsules of adult mice, they can develop into teratocarcinomas (Solter and Knowles, 1978). These embryo transplantation experiments demonstrated that the intact embryo has a cell population that can give rise to pluripotent stem cell lines and this key discovery led to the search for culture conditions that would allow the direct in vitro derivation of pluripotent stem cells from the embryo, without the intermediate need for teratocarcinoma formation in vivo.
Embryonic Germ (EG) cells are isolated from Primordial Germ Cells (PGCs) during the developing gonadal ridge of 5 to 9 week-old fetuses of elective abortions. These cells are diploid germ cell precursors that transiently exist in the embryo before they enter into close association with the somatic cells of the gonad and become irreversibly committed as germ cells (Anway et al., 2003).
Primordial germ cells are pluripotent and are capable of forming all three primordial germ layers. The derivation of EG cells was reported in 1998, but in spite of efforts by several groups, their long-term proliferative potential appears to be limited (Bangso and Richard, 2004; Shamblott et al., 1998). Unlike ES cells, however, EG cells retain some features of the original PGCs, including genome-wide demethylation, erasure of genomic imprints and reactivation of X-chromosomes, the degree of which likely reflects the developmental stages of the PGCs from which they are derived (Yu et al., 2008; Maser and Depinho, 2002).
Other Pluripotent stem cells are multipotent germline stem cells (mGSCs) that more recently have been derived from both neonatal and adult mouse testis. These cells share a similar morphology with mouse ES cells, express typical mouse ES cell specific markers, differentiate into multiple lineages in vitro, form teratomas and contribute extensively to chimeras including germline cells upon injection into blastocysts. However, mGSCs have an epigenetic status distinct from both ES cells and germline stem cells. The mouse testis contains different subpopulations of germline stem cells (Koruji et al., 2007, 2009; Mansoureh, 2009). The origin of mGSCs is still somewhat unclear, though it might be possible that in vitro culture of germline stem cells reprograms a minority of these cells to resume an ES cell-like state (Guan et al., 2006; Izadyar et al., 2008; Kanatsu-Shinohara et al., 2004).
Another type of Pluripoten stem cells is Embryonal Carcinoma cell (ECs). Kleinsmith and Pierce demonstrated that a single EC cell is capable of both unlimited self-renewal and multi lineage differentiation, establishing the existence of a pluripotent stem cell and also providing the intellectual framework for both mouse and human Embryonic Stem (ES) cells. This was also the first experimental demonstration of a cancer stem cell, predating the current intense interest in cancer stem cells by several decades (Yu et al., 2008; Kleinsmith and Pierce, 1964). EC cells express antigens and proteins that are similar to cells present in the ICM, which led to the concept that EC cells are an in vitro counterpart of the pluripotent cells present in the ICM. Indeed, some EC cell lines are able to contribute to various somatic cell types upon injection into mouse blastocysts. However, most EC cell lines have limited developmental potential and contribute poorly to chimeric mice, likely due to the accumulation of genetic changes during teratocarcinoma formation and growth (Gachelin et al., 1977; Solter and Knowles, 1978).
Human embryonal carcinoma (hEC) cell lines have identified from tumours of germ cell origin at the first time. These cells have long served as the human counterpart of murine EC cells for studying human development and differentiation in vitro (Andrews, 2002). Cell lines of hEC are capable of multi lineage differentiation in vitro but, being of tumour origin, are unfortunately mostly aneuploid, which makes them unsuitable for cell-replacement therapeutics. in contrast to mouse EC cells, human EC cells are highly aneuploid, which likely accounts for their inability to differentiate into a wide range of somatic cell types and which limits their utility as an in vitro model of human development (Yu et al., 2008).
A highly plastic, adult-derived bone marrow cell, with features very similar to Mesenchymal Stem Cells (MSC) has also been described as a Multipotent Adult Progenitor Cell (MAPC) (Jiang et al., 2002). These cells are initially isolated together with MSC, but subsequently grow indefinitely in nutrient-poor medium. In specific conditioning media, MAPC can differentiate into cells which express markers of endodermal, mesodermal and ectodermal origin. The same pluripotent ability can be observed in vivo when MAPC are injected into murine blastocysts or when MAPC are injected intravenously into sublethally-irradiated immunodeficient mice. The relationship of MAPC to MSC is unknown. MAPC could be MSC progenitors or may even represent a cell population generated in vitro as an artifact which has no counterpart in vivo (Dazzi et al., 2006).
Multipotency is seen in adult stem cells for example blood stem cells give rise to red blood cells, white blood cells and platelets, whereas skin stem cells give rise to the various types of skin cells. Adult stem cells-also known as somatic stem cells-can be found in diverse tissues and organs. The best-studied adult stem cell is the Hematopoietic Stem Cell (HSC). These cells have been used widely in clinical settings for over 40 years and form the basis of bone marrow transplantation successfully. Unfortunately, HSCs-like many other adult stem cells-are rare and difficult to isolate in large numbers from their in vivo niche (Igura et al., 2004).
Adult stem cells have also been isolated from several other organs such as the brain (neuronal stem cells), skin (epidermal stem cells), eye (retinal stem cells) and gut (intestinal crypt stem cells). Mesenchymal stem cells (MSCs) are another well characterized population of adult stem cells.
Some recent reports suggest that adult stem cells, such as haemopoietic stem cells, neuronal stem cells and mesenchymal stem cells, could cross boundaries and differentiate into cells of a different tissue. This phenomenon of unprecedented adult stem cell plasticity has been termed ‘transdifferentiation’ and appears to defy canonical embryological rules of strict lineage commitment during embryonic development (Bjornson et al., 1999; Krause et al., 2001).
APPLICATIONS OF EMBRYONIC STEM CELLS
Therapeutic applications: Cell therapy in diverse organs has bloomed for degenerative diseases over the past decade, following a previous period of development in which haematopoietic stem cells grafts in oncology were its most prominent clinical application (Mitjavila-Garcia et al., 2005). On the other hand, biological therapies have emerged clinically more than 20 years ago with haematopoietic stem cells and have since, particularly over the past 10 years, rocketed up in research laboratories as well as, already, in the clinic for some indications.
It is anticipated that research with embryonic stem cells will help produce cells and tissue for replacement therapies for treating disorders such as Parkinson's disease, heart attacks, blood disorders and diabetes (Rajagopal et al., 2003).
Two groups can be identified in those biological therapies on the basis of their goal, which may be to substitute for a lost cell population (substitutive cell therapy) or for the delivery of proteins endowed with therapeutic properties (protective cell therapy, usually by gene transfer).
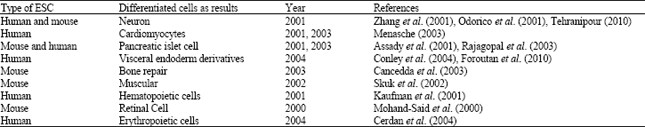
| Table 1: | Some researches about differentiation of embryonic stem cells (human or mouse) to different somatic cells |
 | |
Up to now, most clinical applications have been in substitutive therapy.
Embryonic stem cells could be of value for creating in-vitro culture systems and animal models that may be used to study human genetic disease, gene function, drug discovery and developmental biology (Fathi et al., 2006).
Human ES cell (hESC) lines have been studied for much less time but have already given rise to neurons (Zhang et al., 2001), cardiomyocytes (Odorico et al., 2001), pancreatic islet cells (Assady et al., 2001), although, this is controversial, haematopoietic cells (Odorico et al., 2001), red blood cells (Cerdan et al., 2004), or visceral endoderm derivatives (Conley et al., 2004). The first protocol for a guided differentiation of human ES cells has just been published namely towards a dopaminergic neuronal fate (Mitjavila-Garcia et al., 2005). Type 1 diabetes and Parkinsonism emerge as prime candidates for hESC-based cell-replacement therapy. A similar approach utilizing hESC-derived dopaminergic neurons could hypothetically be used to treat Parkinson’s disease (Table 1).
ES cells would be candidates for cell replacement therapies for degenerative diseases, but direct transplantation of ES cells may result in teratoma, while neurons and glial cells are post-mitotic cells and lost the ability of proliferation when grafted to the brain and neurons cannot differentiate according to the environmental cues or migrate to the injury areas of the brain. Thus, the Neural Precursor Cells (NPCs) with the properties of neural stem cells differentiated from ES cells would be suitable candidates for cell replacement (Xu, 2005). So, precursor cells should be used in cell replacement therapies for degenerative diseases except of ES cells.
Some methods for derivation of human ES cells: For the successful therapeutic application of human ES cells, the appropriate derivation, expansion, manipulation and differentiation of the cells are crucial. There are three methods that can be used in the derivation of human ES cells, depending on the quality of the blastocysts. These methods are the immunosurgical method, the whole embryo culture method and the partial embryo culture method (Odorico et al., 2001; Perrier et al., 2004).
In general, the immunosurgical isolation of ICM, a procedure that was developed in 1975 by Solter and Knowles, has become a common practice in the majority of laboratories in which ES cell derivation is conducted. The underlying principle of immunosurgery is the lysis of the trophectoderm using the antibody/ complement reaction. First, the expanded blastocyst is treated with pronase to dissolve the Zona Pellucida (ZP). The ZP-free blastocysts are then treated with anti-human whole serum antibody and with guinea pig complement. The blastocyst, with its lysed trophectoderm, is rinsed in fresh culture medium and the ICM is isolated from the lysed trophectoderm. The isolated ICM is then cultured on a layer of mitotically inactivated feeder cells in a gelatin-coated tissue culture dish, thereby generating the human ES cell line (Oh et al., 2005). This method, however, is associated with a high probability of ICM loss in cases in which the human ES cells are derived from blastocysts with smaller or indistinct ICMs. The immunosurgical method is also significantly associated with a risk of contamination of the human ES cells with animal pathogens (Kim et al., 2005).
The whole embryo culture method can be employed to establish human ES cell lines via the seeding of the entire blastocyst, without its zona pellucida, directly onto a feeder layer (Kim, 2004). This method can be used regardless of the embryo quality, but is most effective in blastocysts that lack any visible ICM. However, trophectodermal overgrowth tends to hinder the expansion of the ICM during the initial steps of human ES cell derivation. Therefore, the partial embryo culture method was developed and can be used in the derivation of human ES cells from blastocysts with smaller ICMs (Oh et al., 2005).
Expansion of embryonic stem cells: Mitotically inactivated cell feeder layers were first used to support epithelial cells and were later successfully adapted for the culture of mouse EC cells (Mitjavila-Garcia et al., 2005) and mouse ES cells (Aladjem et al., 1998).
Medium that is conditioned by co-culture with various cells was found to be able to sustain ES cells in the absence of natural feeders (Yu et al., 2008). These cells serve as feeder cells in embryonic stem cell (ESCs). For many years Murine Embryonic Fibroblast (MEF) has used as feeder cells for culture of human ESCs. The specific role of MEF in prolonging ESCs culture is less clear. It is generally known that MEF provides a befitting environment for the interplay of signaling networks that regulate the fate of ESCs. The feeders cells of xenogeneic or allogeneic origin make it possible to introduce inter-and intra-species transfer of pathogens (Richards et al., 2004). In addition, the use of MEF as feeders has other disadvantages, including a limited performance window that is optimal only between the 4th and 6th passages, so, it is necessary to repeatedly sacrifice mouse fetuses (Shi et al., 2006; Stojkovic et al., 2004). There are some unanswered questions for systems retaining some use of feeder cells and those hESC lines with a history of culture on murine feeders. Firstly, is it possible that there could be a risk of persistence of murine molecules on hESCs? Secondly, could viruses (and possibly prions) from the feeder cells persist and be transmitted to humans by methodologies that retain the use of conditioned medium or animal derived extracellular matrix? Thirdly, could intimate contact with feeder cells, irrespective of the species of origin, lead to cell fusion and generate cells with tumorigenic potential? Hantaan virus, lymphocytic choriomeningitis virus (LCMV) and reovirus-3 have all been detected in mouse colonies in the past and these viruses have notably caused serious infection and even fatalities, in laboratory workers. In addition to the viral issues, there are also concerns regarding the persistence of mouse molecules in hESC cultures and these concerns have been highlighted by the discovery that certain mouse glycoprotein molecules have been identified on the surface of hES cells cultured on mouse embryonic fibroblasts. Such glycoproteins if sustained on the surface of hES cells could result in severe immune reactions against any transplanted cell products from hESCs (Stacey et al., 2006).
The general conclusion that cells grown on mouse feeder cells would not be appropriate for clinical applications has lead to significant efforts to grow hESC lines on human feeder cells including, foetal fibroblasts, skin fibroblasts, placental fibroblasts and a number of other human cell types. Immortalised cells have also been used as feeder cells and feeder cell lines have also been derived from the original hESCs themselves.
There are two methods used. One is an autogenetic feeder system, in which hESCs are cultured on a feeder-free system, after one week, they differentiate spontaneously into fibroblast-like cells. These fibroblast-like cells can then be passaged and serve as feeder cells. The second method is an EB-derived feeder system in which the hESCs are induced to form EB and then these EB were attached onto a substrate for further differentiation. After 10-14 days fibroblast-like cell are collected, passaged and all have the ability to serve as feeder cells (Shi et al., 2006).
Immune rejection for embryonic stem cells and Induced Ploripotent Stem (IPS) cells technique: Contrary to adult stem cells, the embryonic stem can be propagated and even amplified for a long time in culture, without losing its stem cell character. The major problem facing widespread use of embryonic stem cells in cell therapies and organ replacement however is their anticipated rejection by the patient's immune system, which will recognize them as foreign and destroy them (Drukker et al., 2006). Cell therapy is mostly based to date upon the transfer to the recipient of cells that are homologous to those altered by the disease. Whatever the donor and even when the cell therapy product is autologous, this approach requires a very heavy logistics which, in the present conditions, restricts the patients treated to a very limited number. At worst, the logistic burden is so heavy that one cell therapy protocol, although promising in pilot studies cannot be extended.
Whether hES cell therapy will request the same type of immunosuppressive treatment as that currently needed for organ transplant is an important issue because this will be an essential parameter in the eventual extension of the technique to a large number of patients. The transplantation of tissues and organs between two unrelated individuals results almost invariably in graft rejection, unless immunosuppressive therapy is given to control the alloimmune response (Mitjavila-Garcia et al., 2005).
The only way to overcome an immune reaction, or rather to avoid it, is to perfectly match the donor and the recipient. A perfect match, however, exists only between identical twins. For all other patients, donor and recipients are matched as well as possible and the patients must remain on drugs for the rest of their lives, to prevent the occurrence of an immune reaction with deadly outcome. For the vast majority of patients in need of transplants or tissue repair there are however no appropriate donors.
To overcome the problem of immunological incompatibility of allograft of ES-derived cells, the generation of autologous ES cells for every patient by therapeutic cloning has been suggested (Lanza et al., 1999).
Wilmut et al. (1997) demonstrated Trans-acting material in mammalian oocytes is sufficient to change the epigenetic status of a differentiated nucleus to a totipotent state. This discovery completely changed the mindset of developmental biologists who previously thought this was impossible in mammals (Solter and Knowles, 1978). Immediately following the derivation of human ES cells, it was suggested that Somatic Cell Nuclear Transfer (SCNT) might be used to create patient-specific stem cells as Induced Pluripotent Stem (IPS) cells (Yu et al., 2007). Yet applying SCNT to human material has proven challenging and has only very recently been achieved in nonhuman primates (12). This approach, would lead to the production of cells and tissue matching the patient and that would not cause an immune reaction when the cells are transplanted into the patient (Yu et al., 2008).
For nuclear transfer, the DNA from any one cell in the body of a patient (usually a skin or muscle cell) could be removed and transferred through a microscopic glass tube into an unfertilized egg that previously had its own DNA removed (Campbell et al., 1927).
In a culture dish, the egg is then coaxed into developing as if it had been fertilized. The one egg cell divides rapidly and generates the blastocyst. The inner cell mass, a part of the blastocyst, is then removed and embryonic stem cells grown out of it. These embryonic stem cells, containing the patient's DNA, now match the patient's profile and will not be rejected by the patient's immune system. These embryonic stem cells can now be used to generate cells and tissues for the patient (Mitjavila-Garcia et al., 2005). The success of SCNT demonstrated that reprogramming could be mediated by transacting factors. Through cell-cell fusion, EC cells, ES cells and EG cells were also capable of reverting the differentiated state of somatic cells to that of pluripotent stem cells. Induced Pluripotent Stem (iPS) Cells are obtained by reprogramming an ordinary somatic (body) cell, such as a skin cell, back to a pluripotent state (Baguisi et al., 1999).
While this procedure sounds straightforward and is being performed successfully in animal models, it was only very recently performed successfully with human eggs and human DNA. However, many technological, legal and ethical hurdles need to be overcome before this procedure can be used for human therapy.
APPLICATIONS OF ADULT STEM CELLS
Thrapeutic applications: Adult stem cells are stem cells that come from different parts of the body and, depending on where they are from, have different properties. They exist in several different tissues including bone marrow, blood and the brain. Adult stem cells can be obtained in quantities appropriate for clinical applications, so the researchers can used them as good candidates for tissue repair (Tehranipour, 2010). A type of Adult stem cells is Mesenchymal stem cells (MSC).These cells were first identified by Friedenstein who described undifferentiated cells in adult bone marrow which were able to differentiate in vitro along a number of mesenchymal lineages, including bone and cartilage (Dazzi et al., 2006). MSCs are prevalent in bone marrow at low quantities (1 out of 10 000-100 000 mononuclear cells). It is thought that they respond to local injury by dividing to produce daughter cells that differentiate into multiple mesodermal tissue types, including bone, cartilage, muscle, marrow stroma, tendon, ligament, fat and a variety of other connective tissues. Several reports hint that MSCs can form a variety of cell types in the laboratory, including fat cells, cartilage, bone, tendon and ligaments, muscles cells, skin cells and even nerve cells (Bangso and Richard, 2004; Foroutan et al., 2010).
Compared to adult stem cells from other tissues, hematopoietic stem cells are easy to obtain and have been studied by scientists for many years. They were the first stem cells to be used successfully in blood disorder therapies. More recently, their use in treatment of breast cancer and coronary artery diseases is also being explored.
After the first applications of adult stem cells in the haematopoietic system and the skin (epidermal) grafts, one now sees the development of that approach for neurodegenerative diseases like Parkinson’s or Huntington’s, diabetes mellitus, bone repair Duchenne muscular dystrophy or heart failure. Many other potential applications are under study in research laboratories like, quoting only mature examples that are getting into the clinic, retinal pathologies, cartilage, liver necrosis or other neurological diseases (Mitjavila-Garcia et al., 2005).
Origin of adult stem cells: Cord blood has recently emerged as an alternative source of hematopoietic stem cells for the treatment of leukemia and other blood disorders (Gluckman, 2000). The advantages of using cord blood as a source of stem cells are that umbilical cord blood is easily obtained and is abundant. The thousands of babies are born each day and, until recently, umbilical cord blood was discarded after birth. Now, cord blood is collected and stored by public or private cord blood banks. However, there are a limited number of stem cells in any given cord which prevents its generalized use for the treatment of blood disorders in adults. The use of umbilical cord blood stem cells for other uses, such as organ and tissue repair, is under also investigation.
Unlike most other human adult stem cells, mesenchymal stem cells can be obtained from a small bone marrow sample (Caplan, 1991) from a given patient, expanded in culture and given back to the patient (Le Blanc et al., 2004). This would avoid the problems associated with immune rejection of foreign transplanted cells or tissues.
Another type of adult stem cells obtain from tissues that stay there. In that case a small pieces of that tissue is dissected and taken for enzymatic digestion. After that stem cell will be isolated and enriched by specific growth factors and co-culture with specific cells in vitro culture.
Therapeutic applications of spermatogonial stem cells as a type of adult stem cells: Stem cells derived from mammalian adult tissues have much greater differentiation potential and the study of their behavior is important, both biologically and clinically. These cells provide replacements for cells that are lost during physiological turnover, or damaged by injury, aging and disease (Tadokoro et al., 2002). Several studies suggested that spermatogonial stem cells diminish or lose their activity as male ages. The enhancement and proliferation of reminded stem cells, not only is need for success in transplantation but also it might be helpful for treatment infertility (Koruji et al., 2007; Mansoureh, 2009). The pioneering work for the establishment of germ cell transplantation was performed by Brinster and Zimmermann (1994) in rodent models. Intratubular microinjection of germ cell suspensions via the rete testis under guidance by ultrasonography into the seminiferous tubules restimulated spermatogenesis in spermatogonial stem cells (Absalan et al., 2008). It has done in primates, rodents, mice, macaque, goat, cattle (Brinster and Nagano, 1998; Dirami et al., 1999; Radford et al., 1999; Russell et al., 1990; Schlatt et al., 1999, 2002).
Transplantation technique can be used in future:
| • | For treatment the men with abstractive azoospermia or cryptorchidism disease (Absalan et al., 2008) |
| • | To prevent infertility in male patients due to chemotherapy or radiotherapy (oncological patients) (Brinster, 2007; Koruji et al., 2008). Currently, many children and young adults are successfully treated for several types of cancer. However, these treatments cause permanent or long-lasting infertility |
| • | Study on the maintenance and proliferation of SSC in culture are enormously valuable in understanding their biology and in identifying crucial growth factors for self-renewal (Koruji et al., 2009) |
| • | To find markers for identification of spermatogonial subtypes |
The hope that someday many debilitating human diseases will be treated with stem cell therapy is inspired by remarkable examples of whole-organ and limb regeneration in animals, as well as the historical success of bone marrow transplants, which have improved the lives of many patients suffering from leukaemia and immunological and other blood disorders (Kubota and Brinster, 2006).
For successful therapeutic application most of the adult stem cells isolated from the body will have to be amplified in culture (Makoolati et al., 2009; Sadri-Ardekani et al., 2009). While, the embryonic stem cell can be coaxed to amplify in culture without losing its capabilities, adult stem cells (with exception of the mesenchymal stem cell) can not be amplified in culture to meaningful cell numbers.
The progress and results obtained in the field of Adult Stem Cells (ASC) show not only their great plasticity but also their many possible uses, in all likelihood no different from those of embryonic stem cells, since plasticity depends in large part upon genetic information, which can be reprogrammed. However, many technological, legal and ethical hurdles need to be overcome before this procedure can be used for human therapy.
CONCLUSIONS
Stem cells-whether cord blood, adult or embryonic- have numerous applications in the areas of scientific research and cell therapy. For researchers, stem cells are the key to understanding how humans develop the way they do. Hopefully, the study of stem cells will unravel the mystery of how an undifferentiated cell is able to differentiate and will also determine what is the signal that triggers the sequence. The greater understanding and possibly even control, of cell division and differentiation is a significant strategy in the battle against dreaded illnesses such as cancer, which is basically the continuous multiplication of abnormal cells. The use of stem cells for the testing of new medicines is another highly-anticipated application. Although certain cells are already utilized for this purpose-cancer cells, for example, are used to tests anti-tumor drugs-testing on pluripotent cells would open up this field to a much broader number of cell types.
The third and possibly most important, application is cell therapy, which is the use of stem cells to produce the cells and tissues required for the renewal or repair of body organs that have been damaged by debilitating and mortal diseases such as cancer, spinal cord injuries, glaucoma, Parkinson’s. Since, the first demonstration that the EC cell component of teratocarcinomas is a pluripotent stem cell, the family of pluripotent stem cell lines that can form teratomas has grown to include ES, EG, adult stem cells. Yet, even with more than 40 years of research on pluripotent stem cells, it is still not possible to write a simple paragraph describing why these special cells can differentiate to all other cell types but others cannot (Yu et al., 2008).
Several hurdles in the manipulation and differentiation of hESCs must also be overcome before the technology can be successfully transferred to the bedside (Izadyar et al., 2000). Cell-replacement therapies require the growth of massive numbers of hESCs, so large-scale hESC culture strategies utilizing bioreactors and perfusion systems must be developed to generate sufficient numbers of cells. High efficiency directed differentiation strategies, safer and purer populations of hESCs and their differentiated progeny and clinically compliant xeno-free hESC cell lines must be produced. Lastly, the tissue rejection concerns with cell-replacement therapy using ES cells have to be overcome (Bangso and Richard, 2004).
A recurrent statement in stem cell biology today is the importance of standardizing culture conditions. Culture conditions have a profound effect on stem cell self-renewal, differentiation and possibly on stem cell plasticity (Kubota et al., 2004). A re-acquisition of plasticity in somatic stem cell transdifferentiation might arise because of in vitro culture conditions that actively promote reactivation and dedifferentiation. It is imperative that hESC lines are created for research as well as for clinical application. Furthermore, the existing pool of hESCs might not be truly representative of the general normal human population because all current hESC lines have been derived from infertile couples.
The study of hESC lines harbouring genetic and other chromosomal defects will also give new insight into the early developmental events and cellular pathology of human diseases (Maitra et al., 2005). Elucidation of the mechanisms by which ESCs maintain pluripotency during prolonged proliferation may give insight to the mechanisms by which cancer cells self-renew, as it follows that both mechanisms would be similar, if not identical. Cellular transplantation based on iPS cells has tremendous clinical potential, but a truly regenerative medicine would direct endogenous cells to participate in the repair of damaged tissues that cannot normally regenerate it and such regeneration would require nonphysiological changes in differentiation status (Hanna et al., 2007). Now that such a dramatic change as the conversion of a somatic cell to a pluripotent stem cell has been achieved, clearly similar screens will be carried out to test whether other nonphysiological transitions can be induced between other cell types by a limited number of factors (Yu et al., 2008). Obviously, it is not yet possible to compare the therapeutic results obtained and obtainable using embryonic stem cells and adult stem cells. For the latter, various pharmaceutical firms are already conducting clinical experiments which are showing success and raising genuine hopes for the not too distant future. On the other hand, the road towards the development of an effective cell-based therapy for widespread use is long and involves overcoming numerous technical, legislative, ethical and safety issues.
REFERENCES
- Aladjem, M., B.T. Spike, L.W. Rodewald, T.J. Hope, M. Klemm, R. Jaenisch and G.M. Wahl, 1998. ES cells do not activate p53-dependent stress responses and undergo p53-independent apoptosis in response to DNA damage. Curr. Biol., 8: 145-155.
CrossRef - Alison, M.R., R. Poulsom, S. Forbes and N.A. Wright, 2002. An introduction to stem cells. J. Pathol., 197: 419-423.
CrossRefPubMedDirect Link - Andrews, P., 2002. From teratocarcinomas to embryonic stem cells. Philos. Trans. R. Soc. Lond. B Biol. Sci., 357: 405-417.
PubMed - Anway, M.D., J. Folmer, W.W. Wright and B.R. Zirkin, 2003. Isolation of Sertoli cells from adult rat testes: An approach to ex vivo studies of Sertoli cell function. Biol. Reprod., 68: 996-1002.
PubMed - Assady, S., G. Maor, M. Amit, J. Itskovitz-Eldor, K.L. Skorecki and M. Tzukerman, 2001. Insulin production by human embryonic stem cells. Diabetes, 50: 1691-1697.
CrossRefDirect Link - Baguisi, A., E. Behboodi, D.T. Melican, J.S. Pollock and M.M. Destrempes et al., 1999. Production of goats by somatic cell nuclear transfer. Nat. Biotechnol., 17: 456-461.
CrossRefDirect Link - Bangso, A. and M. Richard, 2004. History and perspective of stem cell research. J. Best Practice Res., 18: 824-842.
PubMed - Bjornson, C., R.L. Rietzle, B.A. Roynoids, M.C. Magli and A.L. Vescovi, 1999. Turning brain into blood: Hematopoietic fate adapted by adult neural stem cells in vivo. Science, 283: 534-537.
PubMed - Brinster, R.L. and M. Nagano, 1998. Spermatogonial stem cell transplantation, cryopreservation and culture. Semin. Cell Dev. Biol., 9: 401-409.
CrossRef - Brinster, R.L. and J.W. Zimmermann, 1994. Spermatogenesis following male germ-cell transplantation. Proc. Natl. Acad. Sci. USA., 91: 11298-11302.
Direct Link - Cancedda, R., B. Dozin, P. Giannoni and R. Quarto, 2003. Tissue engineering and cell therapy of cartilage and bone. Matrix. Biol., 22: 81-91.
PubMed - Cerdan, C., A. Rouleau and M. Bhatia, 2004. VEGF-A165 augments erythropoietic development from human embryonic stem cells. Blood, 103: 2504-2512.
CrossRef - Conley, B.J., A.O. Trounson and R. Mollard, 2004. Human embryonic stem cells form embryoid bodies containing visceral endoderm like derivatives. Fetal Diagn. Ther., 19: 218-223.
CrossRef - Dirami, G., N. Ravindranath, V. Pursel and M. Dym, 1999. Effects of stem cell factor and granulocyte macrophage-colony stimulating factor on survival of porcine type A spermatogonia cultured in KSOM. Biol. Reprod., 61: 225-230.
Direct Link - Drukker, M., H. Katchman, G. Katz, S.E.T. Friedman and E. Shezen et al., 2006. Human embryonic stem cells and their differentiated derivatives are less susceptible to immune rejection than adult cells. Stem Cells, 24: 221-229.
PubMed - Evans, M.J. and M.H. Kaufman, 1981. Establishment in culture of pluripotential cells from mouse embryos. Nature, 292: 154-156.
CrossRef - Foroutan, T., A. Hosseini, A.A. Pourfatholah, M. Soleimani, K. Alimoghadam and N. Mosaffa, 2010. Peritoneal mesothelial progenitor or stem cell. J. Biol. Sci., 10: 460-464.
CrossRefDirect Link - Gachelin, G., R. Kemler, F. Kelly and F. Jacob, 1977. PCC4, a new cell surface antigen common to multipotential embryonal carcinoma cells, spermatozoa and mouse early embryos. Dev. Biol., 57: 199-209.
CrossRef - Gluckman, E., 2000. Current status of umbilical cord blood hematopoietic stem cell transplantation. Exp. Hematol., 28: 1197-1205.
Direct Link - Guan, K., K. Nayernia, L.S. Maier, S. Wagner and R. Dressel et al., 2006. Pluripotency of spermatogonial stem cells from adult mouse testis. Nature, 440: 1199-1203.
CrossRefDirect Link - Hanna, J., M. Wernig, S. Markoulaki, C.W. Sun and A. Meissner et al., 2007. Treatment of sickle cell anemia mouse model with iPS cells generated from autologous skin. Science, 318: 1920-1923.
CrossRef - Igura, K., X. Zhang, K. Takahashi, A. Mitsuru, S. Yamaguchi and T.A. Takashi, 2004. Isolation and characterization of mesenchymal progenitor cells from chorionic villi of human placenta. Cytotherapy, 6: 543-553.
PubMed - Izadyar, F., L.B. Creemers, F.M. van Dissel-Emiliani, A.M. van Pelt and D.G. de Rooij, 2000. Spermatogonial stem cell transplantation. Mol. Cell Endocrinol., 169: 21-26.
CrossRef - Izadyar, F., F. Pau, J. Marh, N. Slepko and T. Wang et al., 2008. Generation of multipotent cell lines from a distinct population of male germ line stem cells. Reproduction, 135: 771-784.
CrossRefDirect Link - Jiang, Y., B.N. Jahagirdar, R.L. Reinhardt, R.E. Schwartz and C.D. Keene et al., 2002. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature, 418: 41-49.
CrossRefDirect Link - Kahan, B.W. and B. Ephrussi, 1970. Developmental potentialities of clonal in vitro cultures of mouse testicular teratoma. J. Natl. Cancer Inst., 44: 1015-1036.
PubMed - Kanatsu-Shinohara, M., K. Inoue, J. Lee, M. Yoshimoto and N. Ogonuki et al., 2004. Generation of pluripotent stem cells from neonatal mouse testis. Cell, 119: 1001-1012.
Direct Link - Kaufman, D.S., E.T. Hanson, R.L. Lewis, R. Auerbach and J.A. Thomson, 2001. Hematopoietic colony-forming cells derived from human embryonic stem cells. Proc. Natl. Acad. Sci. USA., 98: 10716-10721.
PubMed - Kim, H.S., S.K. Oh, Y.B. Park, H.J. Ahn and K.C. Sung et al., 2005. Methods for derivation of human embryonic stem cells. Stem Cells, 23: 1228-1233.
CrossRef - Kleinsmith, L.J. and Jr. G.B. Pierce, 1964. Multipotentiality of single embryonal carcinoma cells. Cancer Res., 24: 1544-1551.
Direct Link - Krause, D.S., N.D. Theise, M.I. Collector, O. Henegariu and S. Hwang et al., 2001. Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell, 105: 369-377.
Direct Link - Kubota, H., M. Avarbock and R.L. Brinster, 2004. Growth factors essential for self-renewal and expansion of mouse spermatogonial stem cells. Proc. Natl. Acad. Sci. USA., 101: 16489-16494.
CrossRefDirect Link - Kubota, H. and R.L. Brinster, 2006. Technology insight: In vitro culture of spermatogonial stem cells and their potential therapeutic uses. Nat. Clin. Pract. Endocrinol. Metab., 2: 99-108.
PubMed - Lanza, R.P., J.B. Cibelli and M.D. West, 1999. Prospects for the use of nuclear transfer in human transplantation. Nat. Biotechnol., 17: 1171-1174.
PubMed - Le Blanc, K., I. Rasmusson, B. Sundberg, C. Gotherstrom, M. Hassan, M. Uzunel and O. Ringden, 2004. Treatment of severe acute graft-versus-host disease with third party haploidentical mesenchymal stem cells. Lancet, 363: 1439-1441.
PubMed - Maitra, A., D.E. Arking, N. Shivapurkar, M. Ikeda and V. Stastny et al., 2005. Genomic alterations in cultured human embryonic stem cells. Nat. Genet., 37: 1099-1103.
CrossRef - Martin, G.R., 1981. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. USA., 78: 7634-7638.
Direct Link - Maser, R.S. and R.A. Depinho, 2002. Connecting chromosomes, crisis and cancer. Science, 297: 565-569.
CrossRefDirect Link - Menasche, P., 2003. Skeletal muscle satellite cell transplantation. Cardiovasc Res., 58: 351-357.
Direct Link - Mitjavila-Garcia, M.T., C. Simonin and M. Peschanski, 2005. Embryonic stem cells: Meeting the needs for cell therapy. Adv. Drug Delivery Rev., 57: 1935-1943.
Direct Link - Mohand-Said, S., D. Hicks, H. Dreyfus and J.A. Sahel, 2000. Selective transplantation of rods delays cone loss in a retinitis pigmentosa model. Arch. Ophthalmol., 118: 807-811.
PubMed - Odorico, J.S., D.S. Kaufman and J.A. Thomson, 2001. Multilineage differentiation from human embryonic stem cell lines. Stem Cells, 19: 193-204.
CrossRef - Oh, S.K., H.S. Kim, H.J. Ahn, H.W. Seol and Y.Y. Kim et al., 2005. Derivation and characterization of new human embryonic stem cell lines: SNUhES1, SNUhES2 and SNUhES3. Stem Cells, 23: 211-219.
Direct Link - Perrier, A.L., V. Tabar, T. Barberi, M.E. Rubio and J. Bruses et al., 2004. Derivation of midbrain dopamine neurons from human embryonic stem cells. Proc. Natl. Acad. Sci., 101: 12543-12548.
CrossRefDirect Link - Radford, J., S. Shalet and B. Lieberman, 1999. Fertility after treatment for cancer. Questions remain over ways of preserving ovarian and testicular tissue. BMJ., 319: 935-936.
PubMedDirect Link - Rajagopal, J., W.J. Anderson, S. Kume, O.I. Martinez and D.A. Melton, 2003. Insulin staining of ES cell progeny from insulin uptake. Science, 299: 363-363.
PubMedDirect Link - Richards, M., S.P. Tan, J.H. Tan, W.K. Chan and A. Bongso, 2004. The transcriptome profile of human embryonic stem cells as defined by SAGE. Stem Cells, 22: 51-64.
PubMedDirect Link - Sadri-Ardekani, H., S.C. Mizrak, S.K.M. van Daalen, C.M. Korver and H.L. Roepers-Gajadien et al., 2009. Propagation of human spermatogonial stem cells in vitro. JAMA., 302: 2127-2134.
Direct Link - Schlatt, S., S.S. Kim and R. Gosden, 2002. Spermatogenesis and steroidogenesis in mouse, hamster and monkey testicular tissue after cryopreservation and heterotopic grafting to castrated hosts. Reproduction, 124: 339-346.
CrossRefDirect Link - Schlatt, S., G. Rosiepen, G.F. Weinbauer, C. Rolf, P.F. Brook and E. Nieschlag, 1999. Germ cell transfer into rat, bovine, monkey and human testes. Hum. Reprod., 14: 144-150.
CrossRefDirect Link - Shamblott, M.J., J. Axelman, S. Wang, E.M. Bugg and J.W. Littlefield et al., 1998. Derivation of pluripotent stem cells from cultured human primordial germ cells. Proc. Natl. Acad. Sci., 95: 13726-13731.
Direct Link - Shi, Y.T., Y.Z. Huang, F. Tang and J.X. Chu, 2006. Mouse embryonic stem cell-derived feeder cells support the growth of their own mouse embryonic stem cells. Cell Biol. Int., 30: 1041-1047.
PubMedDirect Link - Skuk, D., J.T. Vilquin and J.P. Tremblay, 2002. Experimental and therapeutic approaches to muscular dystrophies. Curr. Opin. Neurol., 15: 563-569.
PubMedDirect Link - Solter, D. and B.B. Knowles, 1978. Monoclonal antibody defining a stage-specific mouse embryonic antigen (SSEA-1). Proc. Natl. Acad. Sci. USA., 75: 5565-5569.
Direct Link - Stojkovic, M., M. Lako, P. Stojkovic, R. Stewart and S. Przyborski et al., 2004. Derivation of human embryonic stem cells from day-8 blastocysts recovered after three-step in vitro culture. Stem Cells, 22: 790-797.
PubMedDirect Link - Tadokoro, Y., K. Yomogida, H. Ohta, A. Tohda and Y. Nishimune, 2002. Homeostatic regulation of germinal stem cell proliferation by the GDNF/FSH pathway. Mech. Dev., 113: 29-39.
PubMedDirect Link - Tehranipour, M., 2010. Bone marrow stromal cells for the treatment of spinal cord injury in rats. J. Biol. Sci., 10: 53-57.
CrossRefDirect Link - Thomson, J.A., J. Itskovitz-Eldor, S.S. Shapiro, M.A. Waknitz, J.J. Swiergiel, V.S. Marshall and J.M. Jones, 1998. Embryonic stem cell lines derived from human blastocysts. Science, 282: 1145-1147.
CrossRef - Xu, Y., 2005. A new role for p53 in maintaining genetic stability in embryonic stem cells. Cell Cycle, 4: 363-364.
PubMedDirect Link - Yu, J., M.A. Vodyanik, K. Smuga-Otto, J. Antosiewicz-Bourget and J.L. Frane et al., 2007. Induced pluripotent stem cell lines derived from human somatic cells. Science, 318: 1917-1920.
CrossRefDirect Link - Zhang, S.C., M. Wernig, I.D. Duncan, O. Brustle and J.A. Thomson, 2001. In vitro differentiation of transplantable neural precursors from human embryonic stem cells. Nat. Biotechnol., 19: 1129-1133.
PubMedDirect Link - Absalan, F., M. Movahedin and S.J. Mowla, 2008. Combination of in vivo cryptorchid testis and in vitro co-culture system to obtain high purification and proliferation of mouse spermatogonial stem cells. Int. J. Fertil. Sterility, 2: 115-120.
Direct Link - Fathi, F., T. Tiraihim, S.J. Mowla and M. Movahedin, 2006. Transfection of CCE mouse embryonic stem cells with EGFP and BDNF genes by the electroporation method. Rejuvenation Res., 9: 26-30.
PubMed - Koruji, M., M. Movahedin, S.J. Mowla and H. Gourabi, 2007. Colony formation ability of frozen thawed spermatogonial stem cell from adult mouse. Iran. J. Reproductive Med., 5: 109-115.
Direct Link - Koruji, M., M. Movahedin, S.J. Mowla, H. Gourabi and A.J. Arfaee, 2008. The morphological changes of adult mouse testes after 60Co gamma-Radiation. Iran. Biomed. J., 12: 35-42.
PubMed - Koruji, M., M. Movahedin, S.J. Mowla, H. Gourabi and A.J. Arfaee, 2009. Efficiency of adult mouse spermatogonial stem cell colony formation under several culture conditions. Cell. Dev. Biol. Anim., 45: 281-289.
CrossRefDirect Link - Makoolati, Z., M. Movahedin and M. Forouzandeh-Moghadam, 2009. Effects of different doses of bone morphogenetic protein 4 on viability and proliferation rates of mouse embryonic stem cells. Yakhteh Med. J., 11: 29-34.
Direct Link - Wilmut, I., A.E. Schnieke, J. McWhir, A.J. Kind and K.H.S. Campbell, 1997. Viable offspring derived from fetal and adult mammalian cells. Nature, 385: 810-813.
CrossRefPubMedDirect Link
Melika Reply
It was great because I understood many things about stem cells by this review. Some articles doesn't be open in our country but I could get abstract about those articles in this paper.
soha Reply
This article is a good review about stem cells. I referred it to everyone that wants to start a research about stem cells.