
Z.C. Cob
Marine Ecosystem Research Center, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
A. Arshad
Department of Aquaculture, Faculty of Agriculture
J.S. Bujang
Department of Biology, Faculty of Science, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
W.L.W. Muda
Marine Ecosystem Research Center, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
M.A. Ghaffar
Marine Ecosystem Research Center, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Journal of Applied Sciences
Year: 2010 | Volume: 10 | Issue: 8 | Page No.: 628-635







ABSTRACT
Strombus canarium is a commercially important gastropod that has great potential for advancement into aquaculture. In this study, the metamorphosis response of Strombus canarium larvae to various metamorphosis cues associated with conch nursery habitat and to KCl and GABA, were tested. Bioassays were run as static, no choice experiment and adopting a continuous exposure approach. Strombus canarium larvae showed strong metamorphosis responses when sediment (i.e., conch nursery habitat sediment/SD-NU) and detrital substrata (i.e., Thalassia detritus leachate/T-LC) from their nursery habitat were used (p<0.05). There was no metamorphosis in treatments using sterilized conch nursery habitat sediment (SD-ST) and sediment taken from outside conch nursery habitat (SD-OT). Experiments using fresh macrophyte blades of Enhalus acoroides (EA), Thalassia hemprichii (TH), Halophila ovalis (HA) and Ulva (UL) and adult conditioned seawater (SD-SW) also showed negative respond. Conch larvae demonstrate active habitat selection during metamorphosis and no spontaneous metamorphosis was observed. Settlement in S. canarium is associative in nature where epibionts associated with conch nursery habitat could be the cue for the metamorphosis. However, the specific epibionts/inducers and mechanisms underlining the process were not studied and therefore are subjected to more detailed investigation. The use of KCl was comparable with treatments using natural inducers (SD-NU and T-LC), thus was suggested for application in hatchery spat production of the species.
PDF Abstract XML References Citation
How to cite this article
Z.C. Cob, A. Arshad, J.S. Bujang, W.L.W. Muda and M.A. Ghaffar, 2010. Metamorphosis Induction of the Dog Conch Strombus canarium
(Gastropoda: Strombidae) Using Cues Associated with Conch Nursery Habitat. Journal of Applied Sciences, 10: 628-635.
DOI: 10.3923/jas.2010.628.635
URL: https://scialert.net/abstract/?doi=jas.2010.628.635
DOI: 10.3923/jas.2010.628.635
URL: https://scialert.net/abstract/?doi=jas.2010.628.635
INTRODUCTION
Metamorphosis in marine invertebrates involved significant morphological and physiological changes that facilitate the transformation of the larvae into young juvenile. Major ecological changes occurred, which include a shift from pelagic, encapsulated or brooded existence to that of an independent benthic individual (Gosselin, 1997; Bryan and Qian, 1998). The transformation process involved lots of energy due to significant increased in metabolic demand during the metamorphosis (Shilling et al., 1996; Marsh et al., 1999; Bryan, 2004). Successful metamorphosis is therefore, one of the most critical parts in supplying new recruits for the natural populations. It is also important for advancement of commercially important invertebrate species into aquaculture. The percentage of successful metamorphosis is considered to be of primary importance and critical for aquaculture (Garcia-Lavandeira et al., 2005) in order to maintain a steady supply and reliable source of seeds.
Metamorphosis in marine invertebrate is an active process and rarely occurred spontaneously. Upon reaching the metamorphosis competence stage invertebrate larvae will actively in search for these cues. In most cases the cues are very specific and generally reflects suitable habitat for the post-metamorphic life phase (Stoner et al., 1996; Williams and Degnan, 2009). Abundant literature has shown that settling marine invertebrate larvae are influenced by specific physical, chemical and biological cues derived from a variety of sources in the marine environment (Manriquez et al., 2004; Sawatpeera et al., 2004; Holmes et al., 2005; Xing et al., 2008; Hayakawa et al., 2009; Williams et al., 2009).
Strombus canarium Linnaeus, 1758 is a widely distributed conch species within the tropical waters and is highly associated with muddy bottom and seagrass bed areas (Purchon and Purchon, 1981; Poutiers, 1998; Cob et al., 2009a). It is a commercially important conch species within the Southeast Asian region (Erlambang and Siregar, 1995; Cob et al., 2008, 2009a, b). It has traditionally been fished and contributes to the economic of the local fishermen (Purchon and Purchon, 1981; Erlambang and Siregar, 1995).
Strombus canarium is a gonochoristic gastropod with separate sexes. Fertilization of egg is internal and large egg masses are laid by attaching them to seagrasses during spawning season from November to March. The egg mass is creamy-white in color and can easily be collected from the seagrass bed during low tides throughout the spawning season (Cob et al., 2009d). The species showed high fecundity, which ranged from 48,000 to 71,000 eggs in a single egg mass (Cob et al., 2009c). The larval stages of this conch have recently been described (Cob et al., 2009c, 2009d) and it shared great similarity with other Strombus species (e.g., Davis et al., 1993; De Navarrete et al., 2007). The species certainly has great aquaculture potential. However, more information is needed, particularly regarding the settlement and metamorphosis of the species.
Recent study showed that there is specific nursery habitat for S. canarium population at Merambong Shoal, in the Straits of Johor, Malaysia (Cob et al., 2009b). The purpose of the investigation was to determine parameters within the conch nursery habitat that might contribute as natural cues for conch larvae settlement and metamorphosis. Therefore, we tested and analyzed the effect of various natural cues associated with the nursery habitat. Apart from that the effect of K+ ion elevation and neurotransmitter GABA on S. canarium larvae were also conducted. The result is practically very important for management and conservation of the resources and hopefully lead to advancements in aquaculture of the species.
MATERIALS AND METHODS
Culture of Strombus canarium larvae: Strombus canarium egg masses were collected from Merambong Shoal, Johor Straits, Malaysia (01°19.778’N, 103°35.798’E). The egg masses were gently washed with 0.22 μm filtered seawater (FSW) to remove sand and debris and incubated in individual containers filled with FSW, at salinity 30±1 PSU and temperature 29±1°C until hatch. Larvae were held at concentrations of 1-2 larvae mL-1 in 2 L round bottom glass container of 30±1 PSU and 29±1°C and fed daily with the algae Isochrysis galbana at 1000 cells mL-1 (Cob et al., 2009c). The culture medium was totally replaced every 2 days with freshly filtered FSW. After 16 days, larvae that exhibit exploration and swim-crawl behavior as well as other physical characteristics that indicates metamorphic competence such as the presence of dark-green pigmentation on shell, foot and mantle margin, development of proboscis etc. (Cob et al., 2009c, d), were utilized for the settlement experiments.
Design of settlement experiments: Metamorphosis assays were run as static, no-choice experiments, with three replicates per treatment. Ten competent larvae were used in each replicate, which were taken from the same egg mass to reduce variability and all experiments were tested simultaneously. Larvae were introduced into a well of a 6-well plate (sterile Nunclon® 6-well polystyrene, 35 mm diameter, 15 mL volume), containing the respective settlement cues in 10 mL of FSW. The well plates (with lid on) were kept in an incubator at 29±1°C and salinity 30±1 PSU, under a 12L:12D light regimen. Larvae were subjected to continuous exposure to the metamorphosis inducers. A treatment with only FSW was used as a negative control, which also act as a test of spontaneous metamorphosis.
Larvae behavior was observed under a dissecting microscope. The larvae were considered have successfully metamorphosed when they lost their velar lobes and began to crawl using their foot (Cob et al., 2009c). The conch juvenile showed active feeding activities immediately after metamorphosed (Cob et al., 2009c). Percentage of metamorphosis was determined at 24, 48, 72 and 96 h after the initiation of the experiment. Abnormal behavior of the newly settled juvenile due to incomplete metamorphosis e.g., slow moving, inactive and/or not using foot to crawl (Boettcher and Targett, 1998; Cob et al., 2009c) were also noted.
Metamorphosis assays Settlement in response to fresh macrophyte blades: The larval settlement response to fresh macrophyte blades i.e., Thalasia hemprichii (TH), Halophila ovalis (HA), Enhalus acoroides (EA) and Ulva sp. (UL) were tested. The macrophytes were taken from conch nursery habitat during extreme low tide when the seagrass bed was exposed. The macrophytes were gently rinsed with FSW and their surfaces were carefully scraped to remove detritus, sediment and other epibionts. A small fragment (ca 1 cm2) of the macrophyte blades were introduced into the respective experimental well.
Settlement in response to seagrass detritus: Senescent and partially decomposed seagrass blades (i.e., macro-detritus) of Thalassia hemprichii (T-DET) and Halophila ovalis (H-DET) were collected from the conch nursery habitat during extreme low tide. The macro detritus was gently rinsed with FSW to remove sediment, but the epibiota were kept intact (not scraped). Fragments of about 1 cm2 of T-DET and H-DET were introduced into the respective experimental well.
Settlement in response to aqueous extract of seagrass detritus: Extract from partially decomposed leaf fragments (macro-detritus) of Thalassia hemprichii (T-LC) and Halophila ovalis (H-LC) were tested. The macro-detritus was gently rinsed in FSW to remove sand and unwanted debris and was further shredded and ground using pestle and mortar. FSW was added at a ratio of ~0.5 g detritus to 1 mL FSW. The solution was kept frozen for 1 week to lyse the cells. Prior the experiment the solution was allowed to thaw overnight and filtered using a qualitative whatman-1 filter paper. Assay concentrations of 10 mL aqueous extracts per 1 L FSW were used in the settlement experiment.
Settlement in response to surface sediment: The uppermost layer (5 mm) of top sediment was used for this experiment. The treatment for this experiment includes: sediment taken from conch nursery habitat (SD-NU), sediment taken from conch nursery habitat but further autoclaved (SD-ST) and sediment taken from outside conch nursery habitat (SD-OT) (i.e., from dense Enhalus bed of more than 80% coverage). The macro-detritus was manually removed from the sediment using forceps and smaller detritus was removed by decanting method using FSW. The sediments were transferred into the experimental wells to about 3-4 mm in thickness. Ten milliliter of FSW was added and the assays were conditioned for 24 h with mild aeration prior to the introduction of competent larvae.
Surface sediment conditioned seawater (SD-SW): Uppermost (5 mm) layer of sediment were taken from conch nursery habitat during extreme low tide. The macro-detritus was manually removed and FSW was added at a ratio of 0.5 g sediment per 2 mL FSW, with mild aeration. After 24 h the conditioned seawater was filtered using a 0.45 μm MF-millipore membrane filter and transferred to the experimental well.
Settlement in response to Adult Conditioned Seawater (AD-SW): Five adult specimens of Strombus canarium were taken from the field. The conchs were rinsed with FSW and their shells were scraped to remove epibionts and fouling and were kept in 2 L FSW with moderate aeration. After 24 h, the seawater was filtered using a 0.45 μm MF-Millipore membrane filter and introduced to the experimental well.
Settlement in response to elevated ion concentrations (K+) and neuroactive compound (GABA): There are many ions and neuroactive compounds that have been found to affect larva settlement in marine mollusks, however, only Potassium chloride (KCl) and γ-aminobutyric acid (GABA) were tested. KCl was selected as it is cheap and economical for commercial application (Yu et al., 2008), whilst GABA has increasing been used by commercial hatcheries because of its higher efficiency compared with other settlement methods (Bernal and Beltran, 1998). The KCl stock solution of 1 M was prepared by dissolving the chemicals in distilled water. Serial dilutions with FSW were made immediately before commencing the experiments. Test concentrations were 5, 10 and 15 mM K+. Stock solution of GABA was prepared by dissolving the chemicals in FSW immediately prior to the experiments. The stock solutions of 10-2 M were diluted to prepare solutions of 10-7, 10-6 and 10-5 M.
Data analysis: There were three replicates of each treatment. The percentage of metamorphosis in each experimental treatment was transformed (arcsine) before the statistical analysis was conducted. To improve the arcsine-transformation, those replicates with zero were given the value of 1/(4n), where n is the number of larvae in a single replicate (Yu et al., 2008). Treatments in which percent metamorphosis was equal to zero for all replicates were not included in the statistical analyses. Normality and homogeneity of variance were tested using Anderson-Darling and Bartlett’s test respectively. Data complied with the requirement for parametrical test were analyzed with a one-way ANOVA utilizing a post-hoc Tukey’s test to compare difference between treatments and control (Yu et al., 2008). Otherwise, the Kruskal-Wallis test followed by Dunn’s method was applied (Gebauer et al., 1998). All statistical analyses were conducted using the MINITAB® Statistical Software (Minitab Inc., State College, USA).
RESULTS
The cumulative percentage of metamorphosis after 24, 48, 72 and 96 h are presented in Table 1. Analysis of variance showed a highly significant different between treatments (oneway-ANOVA, F = 40.39, df = 35, p<0.05). Post-hoc analysis (Table 2) showed three different groups that significantly differed from one another. Higher percentages of metamorphosis were observed in treatments using sediment taken from nursery habitat (SD-NU), Thalassia detritus leachate (T-LC) and 5 mM KCl (p<0.05), followed by a slightly lower metamorphosis in treatments using sediment conditioned seawater (SD-SW), Thalassia detritus (T-DET), Halophila detritus (H-DET), 10 and 15 mM KCl (p<0.05).
| Table 1: | Metamorphosis response of Strombus canarium larvae exposed to different inducers |
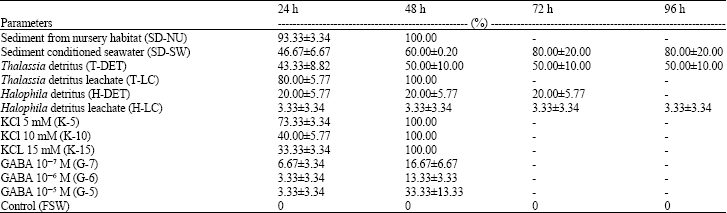 | |
| Values are cumulative percentage of metamorphosis (Mean±SE, N = 3). FSW = 0.22 μm filtered seawater. Assays with negative metamorphosis response were not presented | |
| Table 2: | Tukey’s pairwise comparisons between all treatments after 24 h exposure to the inducers |
 | |
| *Denotes significantly different and zero (o) denotes not significantly different, at 0.05 probability levels. SD-NU: Sediment from nursery habitat, SD-SW: Sediment conditioned seawater, T-DET : Thalassia detritus, T-LC: Thalassia detritus leachate, H-DET: Halophila detritus, H-LC: Halophila detritus leachate , K-5: KCl 5 mM, K-10: KCl 10 mM, G-7: GABA 10-7 M, G-6: GABA 10-6 M, G-5: GABA 10-5 M | |
Lowest metamorphic responses were observed in treatments using GABA (10-7, 10-6 and 10-5 M) and Halophila detritus leachate (H-LC) (p<0.05). There was no metamorphosis occurred in treatments using fresh macrophyte blades (HA, TH, EA and UL), adult conditioned seawater (SD-SW), sterile nursery habitat sediment (SD-ST) and sediment taken from outside conch nursery habitat (SD-OT). In addition, there was no spontaneous metamorphosis as none of the larvae in the control treatments has metamorphosed.
Treatments using sediment from nursery habitat (SD-NU), KCl (5, 10 and 15 mM) and Thalassia detritus leachate (T-LC) showed the best result where all larvae in these treatments successfully metamorphosed after 48 h. Treatment using sediment conditioned seawater (SD-SW) also showed good metamorphic response where, 80% metamorphoses recorded after 96 h exposure. In treatment using Thalassia detritus (T-DET), up to 50% metamorphoses were recorded after 48 h, but no further metamorphosis recorded thereafter. For the treatments using Halophila detritus (H-DET) and Halophila detritus leachate (H-LC), metamorphoses were only observed during the first 24 h (at 20 and 3.33%, respectively).
The neurotransmitter GABA was less efficient as inducers for the species as only 3 to 7% of metamorphoses were recorded. The juveniles from these treatments were relatively in poor shape compared to the other treatments. They were inactive and showed very slow respond when disturbed and the animal remained partially retracted into the shell with their columella opening facing upwards. All juveniles exposed to GABA were subsequently died by day 3 (72 h).
DISCUSSION
Recruitment is an important process that plays a critical role in the population dynamics and community structures of benthic marine organisms, especially those that develop from an initial planktonic larval phase. Successful recruitment can be influenced by both settlement success and early post-settlement mortality and how well they adapt to their new environment after metamorphosis (Stoner et al., 1996; Pinedo et al., 2000; Roberts, 2001). Thus, to some extend there might be some relationship between settlement and benthic invertebrate’s habitat preferences. Previous studies showed that S. canarium is highly associated with seagrass bed ecosystem and there is specific nursery habitat present where small juveniles can be found in large numbers (Cob et al., 2008, 2009a). However, there was no metamorphosis response when the S. canarium larvae were exposed to fresh macrophyte blades (fragments of Halophila ovalis, Thalassia hemprichii and Enhalus acoroides), indicating that S. canarium settlement in seagrass beds may be related to features other than the seagrass itself. The seagrass ecosystem formed an important habitat and refuge for the species, but there might be other factors more important for survivorship of the newly settled juvenile.
Strombus canarium larvae on the other hand showed strong metamorphosis responses when sediment and detrital substrata from their nursery habitat were used, which is in agreement with previous studies on other Strombus species (Davis and Stoner, 1994; Boettcher and Targett, 1996; Stoner et al., 1996). Many other invertebrate larvae also showed the same characteristic where positive responses to associative cues from their favorable feeding habitat were reported by Williams et al. (2007), Xing et al. (2008) and Hayakawa et al. (2009). This behavior might be important to ensure settlement in an environment most appropriate for their survival, as juvenile survivorship differed greatly among habitats with different physical and biological environments (Naylor and McShane, 1997; Herbert and Hawkins, 2006; Walker, 2007).
Highest percentage of metamorphosis was observed when, larvae were exposed to sediment taken from their natural nursery habitat. However, no metamorphosis was observed in treatment using the same sediment but sterilized, indicating that factors other than the physical characteristics of the substrate itself are more important for S. canarium larvae. Benthic sediment has in many instances been reported to emanate important metamorphic inducer for gastropods (Pederson and Page, 2000; Smith et al., 2005; Walker, 2007). The upper layer of seagrass bed sediment is very rich in detritus and other associated epibionts (Kharlamenko et al., 2001) that highly preferred by juvenile Strombus. The present study indicated that these associated epibionts within the nursery habitat might have emanated the most important cues for S. canarium larvae. However, the types of epibionts and their chemical properties however, need further investigation. According to Boettcher and Targett (1998), the types of chemical cues involved in Strombus metamorphosis and the mechanisms that controlling it share general features with chemoreception in adult aquatic invertebrates. They normally consists of chemicals with low molecular weight and soluble in water. Autoclaved sterilization might have destroyed the epibionts as well as other important metabolites (cues) thus, resulted in negative metamorphic response by the larvae.
There was no metamorphosis response when cues associated with conspecific adult conch were used. The result of this experiment suggest that the important cues for S. canarium larvae settlement are not aggregative (cues associated with conspecifics), but rather associative (cues associated with non-conspecific-hosts, prey, or biofilm) in nature. Associative cues produced by marine algae, particularly the rhodophytes, were important in the settlement and metamorphosis of many invertebrate larvae (Morse, 1992; Huggett et al., 2005; Hayakawa et al., 2009). Although, juvenile Strombus lives in colony gregarious settlement has never been reported within the genus. This behavior might be an evolutionary selection to avoid being accidentally eaten up by the adults, which also engulf sediment and other particulate matter during feeding.
In KCl exposure trial, the recovery after 24 h exposure was relatively high and comparable with treatments using natural inducers (i.e., sediment from nursery habitat and Thalassia detritus leachate). A similar study on S. gigas reported between 52%-95% recoveries within 16 h exposure at 15 mM (Davis et al., 1990). Another study by Boettcher and Targett (1998) reported KCl concentration between 20 and 22 mM induced levels of metamorphosis equivalent to those induced by the algal extract. Low concentrations of KCl (between 5 and 15 mM) are therefore suggested for large scale metamorphosis of the species studied, but test on commercial scale still yet to be conducted. Elevated K+ concentration induced metamorphosis in marine invertebrates presumably by depolarizing sensory cells (Boettcher and Targett, 1998). Potassium chloride has considerable potential for S. canarium aquaculture (spat production) mainly because it is readily available, inexpensive, effective and non-toxic (Yu et al., 2008). It has been applied as metamorphosis inducer in other mollusks hatcheries with consistent dosage (Pechenik and Heyman, 1987; Davis et al., 1990; Yu et al., 2008).
The GABA is one of neurotransmitters that had the ability to inhibit velar cilia movement thus, induces the larvae metamorphosis behavior (Kaspar and Mountfort, 1995). GABA has been used by an increasing number of commercial hatcheries, because of it higher efficiency over other settlement methods (Bernal and Beltran, 1998). It has been reported to effectively induced settlement and metamorphosis in many mollusks larvae (Doroudi and Southgate, 2002; Zhao et al., 2003; Garcia-Lavandeira et al., 2005). In contrast, GABA was less effective as metamorphic inducer for S. canarium larvae studied. The percentage of metamorphosis was very low in this treatment. Boettcher and Targett (1996) reported no significant effect on larval metamorphosis when exposed to 1 and 100 μM GABA. The current study also showed evidence of toxic effects, which has been reported in other settlement studies using GABA (e.g., Bryan and Qian, 1998; Li et al., 2006; Yu et al., 2008). The toxicity might be due to long exposure to the chemical or through bacterial infestation as. GABA provide a suitable substrate for bacterial growth.
CONCLUSION
The present study showed that S. canarium larvae demonstrate active habitat selection during metamorphosis. The metamorphosis responses were strongly influenced by the sediment and detrital substrata taken from their natural nursery habitat. Observations suggested that associated epibiont within the conch nursery habitat elicited the important cues for S. canarium larvae metamorphosis. As biological and chemical cues play great roles in the species metamorphosis, further investigations are therefore needed for extraction and characterization of the active compounds and the mechanism of the cues. The experiment also demonstrates that potassium chloride (KCl) has considerable potential in S. canarium aquaculture. It may provide a potentially effective and inexpensive method for enhancing large populations of synchronized spat production in S. canarium hatchery. As difficulties in the commercial culturing of mollusks mainly associated with larval metamorphosis, the establishment of metamorphosis cues for this species provide strong basis for advancement of the species into aquaculture.
ACKNOWLEDGMENTS
The authors would like to thank the deanery and staffs of the Biology Department, Universiti Putra Malaysia and School of Environmental and Natural Resource Sciences, Universiti Kebangsaan Malaysia for technical supports, equipments and laboratory facilities provided. This study was funded by the Malaysian government through Universiti Kebangsaan Malaysia research grant No. UKM-ST-08-FRGS0001-2008.
REFERENCES
- Boettcher, A.A. and N.M. Targett, 1996. Induction of metamorphosis in queen conch, Strombus gigas Linnaeus, larvae by cues associated with red algae from their nursery grounds. J. Exp. Mar. Biol. Ecol., 196: 29-52.
CrossRef - Boettcher, A.A. and N.M. Targett, 1998. Role of chemical inducers in larval metamorphosis of queen conch, Strombus gigas Linnaeus: relationship to other marine invertebrate systems. Biol. Bull., 194: 132-142.
Direct Link - Bryan, P.J., 2004. Energetic cost of development through metamorphosis for the seastar Mediaster aequalis (Stimpson). Mar. Biol., 145: 293-302.
CrossRef - Bryan, P.J. and P.Y. Qian, 1998. Induction of larval settlement and metamorphosis in the abalone, Haliotis diversicolor (Reeve). J. Exp. Mar. Biol. Ecol., 223: 39-51.
CrossRef - Cob, Z.C., A. Arshad, B.J. Sidik, S.M.N. Amin and A.G. Mazlan, 2008. Growth, mortality, recruitment and yield-per-recruit of Strombus canarium Linnaeus, 1758 (Mesogastropoda: Strombidae) from the west Johor Straits, Malaysia. Res. J. Fish. Hydrobiol., 3: 71-77.
Direct Link - Cob, Z.C., A. Arshad, J.S. Bujang and M.A. Ghaffar, 2009. Species description and distribution of Strombus (Mollusca, Strombidae) in Johor straits and its surrounding areas. Sains Malaysiana, 38: 39-46.
Direct Link - Cob, Z.C., A. Arshad, J.S. Bujang and M.A. Ghaffar, 2009. Age, growth, mortality and population structure of Strombus canarium (Gastropoda: Strombidae): Variations in male and female sub-populations. J. Applied Sci., 9: 3287-3297.
CrossRefDirect Link - Cob, Z.C., A. Arshad, M.A. Ghaffar, J.S. Bujang and W.L.W. Muda, 2009. Development and growth of larvae of the dog conch, Strombus canarium (Mollusca: Gastropoda), in the laboratory. Zool. Stud., 48: 1-11.
Direct Link - Cob, Z.C., A. Arshad, J.S. Bujang and M.A. Ghaffar, 2009. Seasonal variation in growth and survival of Strombus canarium (Linnaeus, 1758) larvae. Pak. J. Biol. Sci., 12: 676-682.
CrossRefPubMedDirect Link - Davis, M. and A.W. Stoner, 1994. Trophic cues induce metamorphosis of queen conch larvae (Strombus gigas Linnaeus). J. Exp. Mar. Biol. Ecol., 180: 83-102.
CrossRef - Doroudi, M.S. and P.C. Southgate, 2002. The effects of chemical cues on settlement behavior of blacklip pearl oyster (Pinctada margaritifera) larvae. Aquaculture, 209:: 117-124.
CrossRef - Garcia-Lavandeira, M., A. Silva, M. Abad, A.J. Pazos, J.L. Sanchez and M.L. Perez-Paralle, 2005. Effects of GABA and epinephrine on the settlement and metamorphosis of the larvae of four species of bivalve mollusks. J. Exp. Mar. Biol. Ecol., 316: 149-156.
CrossRef - Gebauer, P., I. Walter and K. Anger, 1998. Effects of substratum and conspecific adults on the metamorphosis of Chasmagnathus granulata (Dana) (Decapoda: Grapsidae) megalopa. J. Exp. Mar. Biol. Ecol., 223: 185-198.
CrossRef - Gosselin, L.A., 1997. An ecological transition during juvenile life in a marine snail. Mar. Ecol. Prog. Ser., 157: 185-194.
CrossRef - Hayakawa, J., T. Kawamura, S. Ohashi, T. Horii and Y. Watanabe, 2009. The Settlement Cues of an Articulated Coralline Alga Marginisporum crassissima for the Japanese Top Shell Turbo cornutus. J. Shellfish Res., 28: 569-575.
CrossRef - Herbert, R.J.H. and S.J. Hawkins, 2006. Effect of rock type on the recruitment and early mortality of the barnacle Chthamalus montagui. J. Exp. Mar. Biol. Ecol., 334: 96-108.
CrossRef - Holmes, S.P., G. Walker and J. van der Meer, 2005. Barnacles, limpets and periwinkles: The effects of direct and indirect interactions on cyprid settlement and success. J. Sea Res., 53: 181-204.
CrossRef - Huggett, M.J., R. de Nys, J.E. Williamson, M. Heasman and P.D. Steinberg, 2005. Settlement of larval blacklip abalone, Haliotis rubra, in response to green and red macroalgae. Mar. Biol., 147: 1155-1163.
CrossRef - Kaspar, H.F. and D.O. Mountfort, 1995. Microbial production and degradation of γ-aminobutyric acid (GABA) in the abalone larval settlement habitat. FEMS Microbiol. Ecol., 17: 205-212.
CrossRef - Kharlamenko, V.I., S.I. Kiyashko, A.B. Imbs and D.I. Vyshkvartzev, 2001. Identification of food sources of invertebrates from the seagrass Zostera marina community using carbon and sulfur stable isotope ratio and fatty acid analyses. Mar. Ecol. Prog. Ser., 220: 103-117.
CrossRef - Smith, P.R.K., C.A. Richardson and R. Seed, 2005. Growth and development of the veliger and juveniles of Polinices pulchellus (Gastropoda: Naticidae). J. Mar. Biol. Assoc. UK., 85: 171-174.
CrossRef - Li, H., W. Lin, G. Zhang, Z. Cai, G. Cai, Y. Chang and K. Xing, 2006. Enhancement of larval settlement and metamorphosis through biological and chemical cues in the abalone Haliotis diversicolor supertexta. Aquaculture, 258: 416-423.
CrossRef - Manríquez, P.H., S.A. Navarrete, A. Rosson and J.C. Castilla, 2004. Settlement of the gastropod Concholepas concholepas on shells of conspecific adults. J. Mar. Biol. Assoc. UK., 84: 651-658.
CrossRef - Marsh, A.G., P.K.K. Leong and D.T. Manahan, 1999. Energy metabolism during embryonic development and larval growth of an antarctic sea urchin. J. Exp. Biol., 202: 2041-2050.
Direct Link - Naylor, J.R. and P.E. McShane, 1997. Predation by polychaete worms on larval and postsettlement abalone Haliotis iris (Mollusca: Gastropoda). J. Exp. Mar. Biol. Ecol., 214: 283-290.
CrossRef - Pechenik, J.A. and W.D. Heyman, 1987. Using KCl to determine size at competence for larvae of the marine gastropod Crepidula fornicata (L.). J. Exp. Mar. Biol. Ecol. 112: 27-38.
CrossRef - Pinedo, S., R. Sarda, C. Rey and M. Bhaud, 2000. Effect of sediment particle size on recruitment of Owenia fusiformis in the Bay of Blanes (NW Mediterranean Sea): an experimental approach to explain field distribution. Mar. Ecol. Prog. Ser., 203: 205-213.
CrossRef - Purchon, R.D and D.E.A. Purchon, 1981. The Marine Shelled Mollusca of West Malaysia and Singapore. Part 1. General Introduction and an account of the collecting Stations J. Moll. Stud., 47: 290-312.
Direct Link - Williams, R.R., S.M. Shjegstad and V.J. Paul, 2007. Larval metamorphic competence in four species of Phestilla (Gastropoda; Opisthobranchia). J. Exp. Mar. Biol. Ecol., 351: 160-167.
CrossRef - Williams, R.R., S.M. Shjegstad and V.J. Paul, 2009. Larval metamorphosis of Phestilla spp. in response to waterborne cues from corals. J. Exp. Mar. Biol. Ecol., 375: 84-88.
CrossRef - Bernal, R.S. and C.A. Beltran, 1998. Optimizing the concentration of gamma-aminobutyric acid (GABA) for inducing larval metamorphosis induction in the red abalone Haliotis rufescens (Mollusca: Gastropod). J. World Aquac. Soc., 29: 463-470.
CrossRef - Shilling, F.M., O.H. Guldberg and D.T. Manahan, 1996. Sources of energy for increased metabolic demand during metamorphosis of the abalone Haliotis rufescens (Mollusca). Biol. Bull., 191: 402-412.
Direct Link - Stoner, A.W., 2003. What constitutes essential nursery habitat for a marine species? A case study of habitat form and function for queen conch. Mar. Ecol. Prog. Ser., 257: 275-289.
Direct Link - Stoner, A.W., M. Ray, R.A. Glazer and K.J. McCarthy, 1996. Metamorphic responses to natural substrata in a gastropod larva: decisions related to postlarval growth and habitat preference. J. Exp. Mar. Biol. Ecol., 205: 229-243.
CrossRef - Walker, J.W., 2007. Effects of fine sediments on settlement and survival of the sea urchin Evechinus chloroticus in northern New Zealand. Mar. Ecol. Prog. Ser., 331: 109-118.
CrossRef - Williams, E.A. and S.M. Degnan, 2009. Carry-over effect of larval settlement cue on postlarval gene expression in the marine gastropod Haliotis asinina. Mol. Ecol., 18: 4434-4449.
CrossRef - Xing, R.L., C.H. Wang, X.B. Cao and Y.Q. Chang, 2008. Settlement, growth and survival of abalone, Haliotis discus Hannai in response to eight monospecific benthic diatoms. J. Applied Phycol., 20: 47-53.
CrossRef - Yu, X., W. He, J.D. Gu, M.X. He and Y. Yan, 2008. The effect of chemical cues on settlement of pearl oyster Pinctada fucata martensii (Dunker) larvae. Aquaculture, 277: 83-91.
CrossRef - Zhao, B., S. Zhang and P.Y. Qian, 2003. Larval settlement of the silver or gold-lip pearl oyster Pinctada maxima (Jameson) in response to natural biofilms and chemical cues. Aquaculture, 220: 883-901.
CrossRef