
O.O. Osemwegie
Department of Plant Biology and Biotechnology, Faculty of Life Sciences,University of Benin, P.M.B. 1154, Benin City, Edo State, Nigeria
J.A. Okhuoya
Department of Plant Biology and Biotechnology, Faculty of Life Sciences,University of Benin, P.M.B. 1154, Benin City, Edo State, Nigeria
A.O. Oghenekaro
Department of Plant Biology and Biotechnology, Faculty of Life Sciences,University of Benin, P.M.B. 1154, Benin City, Edo State, Nigeria
G.A. Evueh
Department of Plant Protection, Rubber Research Institute of Nigeria,P.M.B. 1049, Benin City, Edo State, Nigeria
Journal of Applied Sciences
Year: 2010 | Volume: 10 | Issue: 5 | Page No.: 391-398







ABSTRACT
Permanent plots in rubber plantations and a lowland forest, each measuring 25x25 m, were randomly laid out using coloured ribbons and studied twice a month for macrofungi for a period of 14 months. A total of 435 fruit bodies belonging to 93 different species of macrofungi were encountered, 70% of which were identified. Identified taxa were distributed into 4 Classes, 9 Orders and 28 Families with the class Hymenomycetes and family Tricholomataceae as the best represented taxa. Agaric (52%) and polypores mushrooms (31%) were also recorded as the best represented life-forms while wood-based substrates recorded 70% of the total mushroom taxa encountered during the study. The species richness and diversity estimate of 100 randomization accumulation sample order of mushroom abundance data from each of the sampled plots showed that the forest (Plot E) had the best species richness and diversity index values compared to plot A, B, C and D.
PDF Abstract XML References Citation
How to cite this article
O.O. Osemwegie, J.A. Okhuoya, A.O. Oghenekaro and G.A. Evueh, 2010. Macrofungi Community in Rubber Plantations and a Forest of Edo State, Nigeria. Journal of Applied Sciences, 10: 391-398.
DOI: 10.3923/jas.2010.391.398
URL: https://scialert.net/abstract/?doi=jas.2010.391.398
DOI: 10.3923/jas.2010.391.398
URL: https://scialert.net/abstract/?doi=jas.2010.391.398
INTRODUCTION
Defining the population of fungi globally has in recent times remained a challenge to mycologists all over the world (Wood, 1992). Many mycologists agreed that there are more fungi in the world than reported, especially in the tropics and sub-saharan Africa and this has resulted in the inconsistency associated with global fungi estimate reported by Hammond (1992), Hawksworth (1993) and Crous et al. (2006). Scanty information abounds on the diversity of African macrofungi (Mueller et al., 2007; Osemwegie et al., 2006). In Nigeria, mushrooms are often overlooked in many biodiversity studies compared to plants and animals (Idu et al., 2007). Mushrooms in Nigeria are poorly collected, sparingly studied and relatively underutilized (Osemwegie and Okhuoya, 2009; Labarere and Menini, 2000). Higher plants are preferred by most Nigerian as sources of food amidst sporadic reports on the use of some mushrooms as food, food supplements and in folk medicine practices, especially by the rural populace (Akpaja et al., 2005; Okhuoya and Akpaja, 2005; Osemwegie et al., 2006).
In some developed countries of the world, macrofungi are exploited for economic gains in the areas of food security and foreign exchange earnings via large scale mushroom cultivation, mushroom export and pharmacopoeia. They are also used to improve silviculture, agroforestry and agriculture and industries such as brewing, beverage, enzymes, dye, paper mill, organic acids, hormones and animal feeds industries (Arora, 1989; Chang and Miles, 1991; Wainwright, 1992; Wasser, 2007). Mushrooms are also applied in waste management and remediation of contaminated arable lands and waters (Wasser, 2007). Several researchers both in Nigeria and abroad have reported that many macrofungi are potential biological control agents of insects, arthropods and other microorganisms of bacteria and fungi origin (Roberts and Hajek, 1993; Boa, 2004; Jonathan and Fasidi, 2005; Gbolagade et al., 2007).
In Nigeria, mushroom researches have focused more on low-cost cultivation of many indigenous edible mushrooms, their nutriceutics and ethnomycology rather than their diversity, taxonomy, biogeography and ecology (Rammeloo and Walleyn, 1993; Osemwegie and Okhuoya, 2009). Lodge et al. (1995) remarked that the knowledge of mushroom composition and ecology are central to efforts establishing proactive conservation strategies and identifying areas in urgent need of conservation as well as species in short- and long-term danger of extinction. Studies on the diversity of wild macrofungi indigenous to Nigeria are regional and biased to agroecosystems (Osemwegie and Okhuoya, 2009).
This study aimed at identifying the diverse mushroom taxa associated with rubber agroforests and comparing their mushroom community with that of a secondary uncultivated forest within the same ecological zone.
MATERIALS AND METHODS
Study area: The study area, Rubber Research Institute of Nigeria (RRIN) Iyanomo is located in Ikpoba-Okha local Government Area of Edo State, approximately about 29 km from Benin City (Fig. 1). The geographical, ecological and edaphic characteristics are as enumerated in Table 1.
Sampled forest and rubber plots: Permanent plots, each measuring 25x25 m were laid out from randomly selected rubber plantations and a lowland secondary rainforest all connected by a common road. The plots were each approximately 5 m away from the edge of the road. Plot C and D were old (50-55 years old), no longer being tapped for latex and characterized by thick undergrowth and broad canopy cover while Plots A and B were younger (38-43 years old) populated by rubber trees being tapped for latex and weeded once every year. Plot E was a secondary forest with thick undergrowth and rich tree diversity. The plots were each surveyed twice a month for mushrooms for a period of 14 month which ran from 2006 through to 2007 across season gradients. Observed mushrooms were collected and preserved according to Lodge et al. (2004). Vouchered mushrooms were kept in the Mushroom Biology Unit of the Department of Plant Biology and Biotechnology, University of Benin, Benin City, Edo State for further molecular verification. Encountered macrofungi were photographed in situ and features such as phenology, smell, habit, colour, nature of substrate and associations recorded before transportation to the laboratory for identification.
| Table 1: | Geographic ecological and edaphic characteristics of the study area |
 | |
Identification was based on macroscopic features and nomenclature was carried out using a variety of field monograph of coloured mushrooms and books (Largent and Their, 1984; Largent, 1986; Arora, 1991; Mueller et al., 2004; Lincoff, 2005). The number of fruit bodies per sampled data was later subjected to analysis using Estimates statistical package according to Colwell (2005) and Chao et al. (2005).
RESULTS AND DISCUSSION
A total of 93 different species of macrofungi represented by 435 fruit bodies (abundance value) were encountered during the 14 months period of study from which 70% were already identified and named (Table 2). The identified species were distributed into 28 Families, 9 Orders, 4 Class and 2 Phyla (divisions). The most represented mushroom life-forms encountered in the study area was the fleshy fungi (gilled or agaric mushrooms) and polypores comprising of 52 and 31% species, respectively. The earth-stars, puffballs, tubers and cup fungi were the least represented (Table 3). The family Tricholomataceae and members of the class Hymenomycetes were the best represented taxa (Table 3). The study showed that wood and litter-based substrates supported the growth of 70 and 23% of mushrooms taxa observed during the study, respectively while the soil-based substrate recorded the least (Fig. 1). Mushrooms such as Chlorophyllum sp., Coprinopsis atramentarius (Bull.) Redhead, Vilgalys and Monclavo, Hygrocybe sp. and Pleurotus tuberregium (Fr.) Sing were observed to colonize more than one type of substrate or exhibited flexible substrate propensity (Table 2).
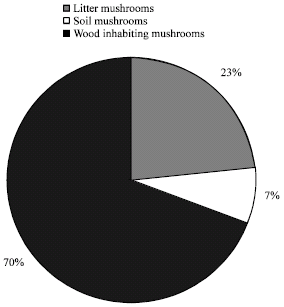 | |
| Fig. 1: | Substrate types and quantitative representation of mushrooms that inhabit the |
| Table 2: | List of macrofungi observed per sampled plot and their substrate propensity |
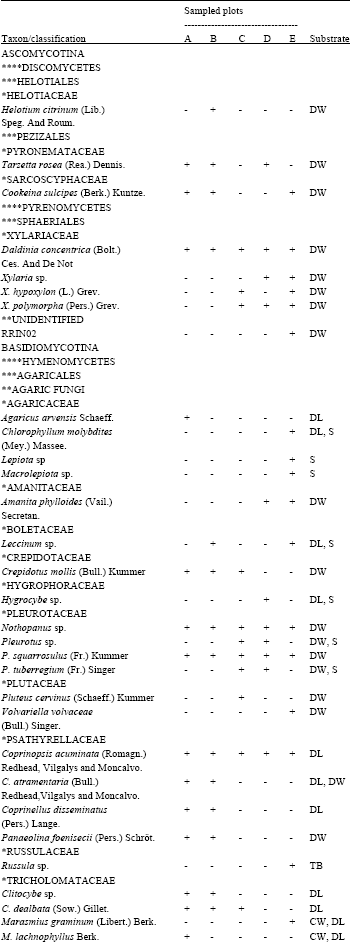 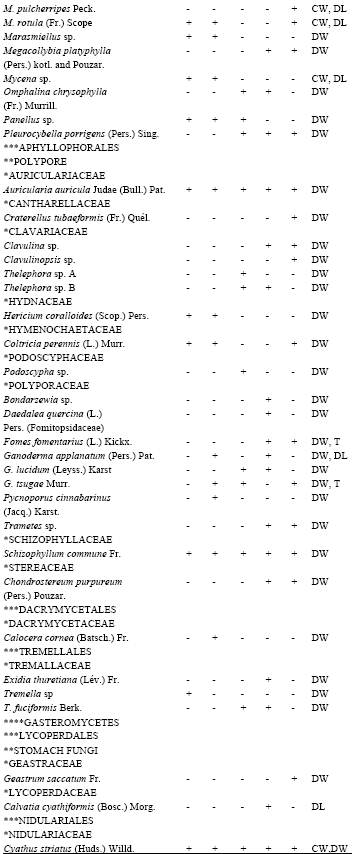 | |
| Plot A and B = 38-43 years old; Plot C and D = 50-55 years old. ****Class, ***Order, **Group, *Family, +: Present, -: Absent, AYR: All year round, BW: Burried wood, CW: Coarse wood, DL: Decomposing litters, DW: Dead decaying wood (tree stump and fallen logs), S: Soil, T: Living tree, TB: Tree branch | |
Rubber tree-dominated secondary forest (Plot E) recorded the highest number of mushroom species (41) and abundance while Plot D which is one of the older rubber plots surveyed recorded 36 different types of macrofungi (Table 4, 5). The highest number of unshared species (16) was also recorded for Plot E and the least (4) for Plot B. Plots A and B were observed to be the most similar recording similarity index values (Chao shared estimate, Jaccard, Sorensen and Morisita-Horn classic) closer to 1 compared to similarity indices recorded for other sampled plots (Table 6).
| Table 3: | Distribution and amount of mushrooms across life-forms and taxonomic hierarchy |
 | |
| Table 4: | Number of mushrooms encountered per sampled plots |
 | |
Plot E also recorded the highest biodiversity indices and the least number of shared mushroom species amounting to 19% of the total mushroom taxa. It was also most varied in terms of mushroom composition compared to Plots A, B and C (Table 5).
Wood-substrate colonizers like Auricularia auricular Judae (Bull.) Pat., Coprinopsis acuminatus (Romagn.) Redhead, Vilgalys and Monclavo, Cyathus striatus (Huds.) Willd., Daldinia concentrica (Bolt. ex Fr.) Ces., Nothopanus sp., Pleurotus squarrosulus (Fr.) Kum., Volvariella volvaceae (Bull.) Singer and Schzophyllum commune Fr were observed in all the plots surveyed. They were also observed all the year round. Auricularia auricular, S. commune, P.tuberregium, Agaricus arvensis Schaeff., P. squarrosulus and Pluteus cervinus (Schaeff. ex Fr.) Kum were some of the indigenous edible mushrooms recorded.
A survey of rubber agroforests and a secondary forest in Rubber Research Institute of Nigeria recorded 93 different mushrooms which amounted to a total of 435 fruit bodies (abundance value) belonging to 28 families within the 14 months period of study. Straatsma and Krisai-Greilhuber (2003) constitute an average of 4.9 fruit bodies per species per month over a total study area of 3125 m2. The number of mushroom taxa recorded from the study contrast with similar work done by Shigeki et al. (1994) in young forests and evergreen broad-leaved forests, Straatsma et al. (2001) in Swiss forests and Osemwegie and Okhuoya (2009) in oil palm agroforests of Edo State.
| Table 5: | Computation of biodiversity indices±SD per sampled plot using 100 randomized sample order |
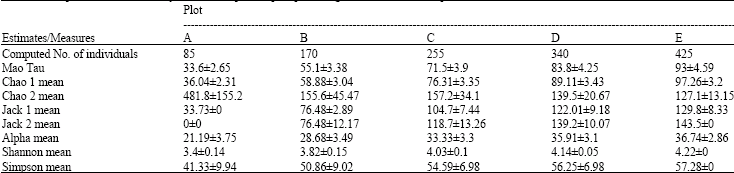 | |
| Mao Tau: No. of observed species | |
| Table 6: | Comparing the sampled plots’ mushroom composition using a similarity index programme in estimates |
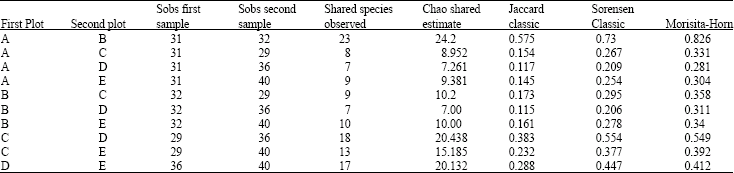 | |
| Values closer to 1 are more similar in species composition | |
The differences in the amount and assemblage of recorded mushroom taxa may be due to variations in sample frequency and time, land area covered during surveys, the nature of woodland vegetation (homo-or heteroculture; riparian or lowland or savannah etc.) studied and geographical location. This apparently proves that many mushrooms such as those that produce hypogeous and ephemeral fruit bodies may have been missed by the study and remains to be discovered with further sampling (Lynch and Thorn, 2006). Reports such as those of Nicholson (2000) and Osemwegie et al. (2006) on mushroom diversity in Nigeria portrayed agroecosystems as impoverished in mushroom diversity and overlooked as viable site for mushroom studies. Flynn et al. (2009) emphasized their ultimate ecological function of provisioning ecosystem services such as biogeochemical cycles, soil binding, decomposition, soil conditioning and regulation of ecosystem balance which supports the well-being of other biotas. These services amongst which are mycorrhization may be harnessed for the development and management of agroecosystems in Nigeria as reported in some African countries (Marx et al., 1993). Bolger (2001) and Loreau et al. (2001) observed a positive relationship between rich biodiversity and ecosystem functions while also recognizing the functional value of species in forming the ideological framework for improving the performance and productivity and decreasing the input of energy, chemical fertilizers and pesticides in agricultural systems. Further studies are required to fully understand the relevance of mushroom community or assemblage to the overall health of rubber plantations. This present study has provided the insight and bases by the sheer diversity of macrofungi recorded in the rubber plantations.
A break down of the result obtained from the study showed that the rubber agroforest plots (A, B, C and D) sampled recorded mushroom diversity whose composition varied with their relative age and level of human disturbance. Plot E however recorded the highest number of fruit bodies amounting to 40 species of mushrooms and this was reflected in the values of Mau Tao (93±4.6), Alpha (36.7±2.9), Shannon (4.2±0.0) and Simpson (57.3±0.0) diversity indices computed while Plot A recorded the least. The tree heterogeneity and according to Tsui et al. (1998) the low level of human disturbances associated with plot E may be responsible for the higher value of species diversity indices recorded. It is however, important to note that further study is required to adequately understand the qualitative and quantitative impact of human dynamics in structuring the mushroom composition of a vegetation. This result therefore supports the line of thought that human activities do impinge on mushroom diversity and stands in agreement with existing scientific mushroom biodiversity findings that relate mushroom diversity to tree diversity (Sala et al., 2000; Jumpponen et al., 2004). Lodge et al. (1995), Laitung and Chauvet (2005) and Mueller et al. (2007) observed a parallel relationship between tree diversities and mushroom richness while Hawksworth (2001) recognized the use of trees in the estimation of global mushroom diversity. Conversely, the varying degrees of rubber latex-tapping activity that characterized the other plots studied and their respective tree homogeneity may be responsible for the relatively low incidence of mushroom taxa and diversity.
A similarity index analysis of the various sampled plots according to Chao et al. (2005, 2006) showed that Plots A and B were the most similar in terms of species composition, sharing 74% (23 species) of their recorded mushroom taxa while Plot E only shared 19% (9 species) of its total taxa. The reason for this is not yet fully understood but it might be connected to variations in the overall nature (diversity of trees, other biota, climate, landscape, productivity or turnover) and intrinsic configuration (tree girth and distance from one another, physiognomy or vegetation layers, canopy spread, gaps, fragmentation) of each of the sampled plots.
The study also recorded 52% agaric and 31% polypore fungi, respectively while other mushroom life-forms such as earth stars, puffballs and tubers were scanty. Agaric and polypore fungi are mostly saprotrophic and capable of biodegrading many recalcitrant organic-based substrates (Lynch and Thorn, 2006). This inherent attribute coupled with their intrinsic enzyme spectrum and dynamics which according to Schmit (2005) consequently broadens accessible substrate-based options, may be the reason for their high representation. The high level of accessible energy resources (cellulose, hemicellulose and lignin) fixed in diverse wood-based substrates in the various sampled plots may have also accounted for the 70% wood-inhabiting mushrooms recorded during the study. Consequently, the volume of wood and its distribution within the sampled plots may have accounted for the high incidence of unshared species (16) observed in Plot E as compared with Plot B which recorded 4 unshared species. This result supports research findings illuminating wood-based substrate as a major determinant of mushroom diversity in woodland vegetations (forest and agroforests) in both temperate and tropical regions. Although, little is known about variations in the pattern of wood resource utilization by different species of macrofungi, these factors in addition to the nature of substrate chemistry and microenvironment may have impacted more on the distribution of mushrooms than species richness in both agroforest and forest systems. The high (31%) incidence of fleshy (agaric) mushroom life-forms recorded during the study correlates positively with increased representation of members of the family Tricholomataceae most of which were litter mushrooms. Chlorophyllum species, C. atramentarius, P. tuberregium and Hygrocybe species were observed to fruit on both soil and wood substrates. This wider substrate colonization propensity observed amongst some of the macrofungi may have also played a fundamental role in the higher incidences of polypore and agaric mushroom life-forms recorded during the study.
Auricularia auricular, C. acuminate, C. striatus, Daldinia concentrica (Bolt.) Ces. and DeNot., Nothopanus sp., P. squarrosulus and S. commune were observed throughout the study area, overlapping boundaries of sampled plots. This characteristic may be attributed to the availability of widely distributed rich nutrient-based substrates (wood debris). In addition, this observation is in concert with of Ozinga et al. (2009) that the dynamism rather than the mechanism of their spore dispersal in space (long-distance travel) and/or time (dormancy or rest period) can determine the biogeographic spread of mushroom taxa.
The relatively large number of unidentified species incurred by the study was due to dearth of previous studies, expert mushroom taxonomists, revised mushroom diversity data especially on Nigeria’s mycoflora and foreign technical supports (Osemwegie and Okhuoya, 2009). Researchers and mushroom scientists are challenged to inventory the nation’s mushroom heritage and explore the grey areas of mushroom taxonomy, ecology and biotechnology studies in Nigeria without prejudice to any vegetation.
Agroecosystems were hitherto perceived as poor in mushroom diversity are by this study recognized as good alternative and sustainable sources of mushroom resources with unprecedented utilitarian values. The study recorded popular edible and medicinal mushrooms such as A. arvensis, A. auricular, Macrolepiota sp., P. tuberregium, P. squarrosulus, Pluteus cervinus, S. commune and V. volvacea in rubber agroforests. It also lends credence to claims that rich tree diversity facilitated luxuriant growth of mushroom-forming fungi which ab initio provide ecosystem services and ecological energy-balance. Furthermore, the study recognized the superiority of forests over agroforests in terms of mushroom assemblage, diversity, abundance and species richness. The contribution of mushrooms to woodland systems was conceptualized by Lawton (1994) and Giller and O’Donovan (2002), who reiterated the need to conserve and preserve national indigenous mushroom flora as a tool in the whole complex process of forest and agroforest management.
ACKNOWLEDGMENTS
We thank the managements of the University of Benin for providing part of the grant for the research and the Rubber Research Institute of Nigeria (RRIN) for approving the use of her rubber plantations and forest for the research. The contributions of Dr. O.S Isikhuemhen of North Carolina State University and Dr. Cathie Aime of Louisiana State University in the identification of some of the mushroom taxa cannot also go on appreciated.
REFERENCES
- Arora, D., 1991. All that the Rain Promises and More: A Hip Pocket Guide to Western Mushrooms. Illustrated Edn., Ten Speed Press, Berkeley, USA., ISBN: 0-89815-388-3 pp: 263.
Direct Link - Bolger, T., 2001. The functional value of species biodiversity: A review. Biol. Environ.: Proce. Royal Irish Acad., 101B: 199-224.
Direct Link - Chao, A., L.R. Chazdon, K.R. Colwell and J.T. Shen, 2005. A new statistical approach for assessing compositional similarity based on incidence and abundance data. Ecol. Lett., 8: 148-159.
CrossRefDirect Link - Chao, A., L.R. Chazdon, K.R. Colwell and J.T. Shen, 2006. Abundance-based similarity indices and their estimation when there are unseen species samples. Biometrics, 62: 361-371.
CrossRefDirect Link - Crous, W.P., H.I. Rong, A. Wood, S. Lee and H. Glen et al., 2006. How many species of fungi are there at the tip of Africa? Stud. Mycol., 55: 13-33.
CrossRefDirect Link - Flynn, B.F.D., M. Gogol-Prokurat, T. Nogeire, N. Molinari and T.B. Richers et al., 2009. Loss of functional diversity under land use intensification across multiple taxa. Ecol. Lett., 12: 22-33.
CrossRefDirect Link - Jonathan, S.G. and I.O. Fasidi, 2005. Antimicrobial activities of some selected Nigerian mushrooms. Afr. J. Biomed. Res., 8: 83-87.
Direct Link - Giller, S.P. and G. O`Donovan, 2002. Biodiversity and ecosystem functions: Do species matter? Biol. Environ.: Proc. Royal Irish Acad., 102B: 129-139.
Direct Link - Hawksworth, D.L., 2001. The magnitude of fungal diversity: The 1.5 million species estimate revisited. Mycol. Res., 105: 1422-1432.
CrossRef - Jumpponen, A., W.A. Claridge, M.J. Trappe, T. Lebel and L.D. Claridge, 2004. Ecological relationships among hypogeous fungi and trees: inferences from association analysis integrated with habitat modeling. Mycologia, 96: 510-525.
Direct Link - Labarere, J. and G.U. Menini, 2000. Collection, characterization, conservation and utilization of mushrooms, germplasm resources in Africa. Proceedings of the 1st International Congress for the Characterization, Conservation, Evaluation and Utilization of Mushroom Genetic Resources for Food and Agriculture, March 9-13, 1998, FAO, Bordeaux, France, pp: 17-63.
- Laitung, B. and E. Chauvet, 2005. Vegetation diversity increases species richness of leaf-decaying fungal communities in woodland streams. Arch. Hydrobiol., 164: 217-235.
CrossRefDirect Link - Lodge, J.D., I. Chapela, G. Samuels, A.F. Uecker and D. Desjardin et al., 1995. A survey of pattern of diversity in non-lichenized fungi. Mitt. Eidgenoss. Forsch.anst. Wald Schnee Landsch, 70: 157-173.
Direct Link - Loreau, M., S. Naeem, P. Inchausti, J. Bengtsson and J.P. Grime et al., 2001. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science, 294: 804-808.
CrossRefDirect Link - Lynch, M.D.J. and R.G. Thorn, 2006. Diversity of basidiomycetes in Michigan agricultural soils. Applied Environ. Microbiol., 72: 7050-7056.
CrossRefDirect Link - Mueller, M.G., J.P. Schmit, P.R. Leacock, B. Buyck and J. Cifuentes et al., 2007. Global diversity and distribution of macrofungi. Biodivers. Conserv., 16: 37-48.
CrossRefDirect Link - Okhuoya, J.A. and E.O. Akpaja, 2005. Mycomedicine and ethnomycology: The Nigerian experience. Int. J. Med. Mushrooms, 7: 439-439.
Direct Link - Osemwegie, O.O., E.G. Eriyaremu and J. Abdulmalik, 2006. A survey of macrofungi in Edo/Delta region of Nigeria, their morphology and uses. Global J. Pure Applied Sci., 12: 149-157.
Direct Link - Ozinga, A.W., C. Romermann, M.R. Bekker, A. Prinzing and M.L.W. Tamis et al., 2009. Dispersal failure contributes to plant losses in NW Europe. Ecol. Lett., 12: 66-74.
CrossRefDirect Link - Sala, O.E., F.S. Chapin III, J.J. Armesto, E. Berlow and J. Bloomfield et al., 2000. Global biodiversity scenarios for the year 2100. Science, 287: 1770-1774.
CrossRefDirect Link - Schmit, P.J., 2005. Species richness of tropical wood-inhabiting macrofungi provides support for species-energy theory. Mycologia, 97: 751-761.
Direct Link - Shigeki, I., S. Shigeki and Y. Osamu, 1994. Analysis of mushroom diversity in successional young forests and equilibrium evergreen broad-leaved forests. Mycoscience, 35: 1-14.
Direct Link - Straatsma, G. and I. Krisai-Greilhuber, 2003. Assemblage structure, species richness, abundance and distributon of fungal fruit bodies in a seven year plot-based survey near Vienna. Mycol. Res., 107: 632-640.
CrossRef - Tsui, K.M., CS. Fryar, J.I. Hodgkiss, D.K. Hyde, D.A. Poonyth and E.J. Taylor, 1998. The effect of human disturbances on fungal diversity in the tropics. Fungal Diversity, 1: 19-26.
Direct Link - Wasser, P.S., 2007. A book review: Mycelium running: How mushrooms can help save the world. Herbalgram, 76: 50-57.
Direct Link - Straatsma, G., F. Ayer and S. Egli, 2001. Species richness, abundance, and phenology of fungal fruit bodies over 21 years in a Swiss forest plot. Mycol. Res., 105: 515-523.
CrossRefDirect Link - Osemwegie, O.O. and J.A. Okhuoya, 2009. Diversity of macrofungi in oilpalm agroforest of Edo State Nigeria. J. Biol. Sci., 9: 584-593.
CrossRef