
K. Fadzilah
School of Chemical Engineering, Universiti Sains Malaysia, Engineering Campus, 14300, Nibong Tebal, Malaysia
M.D. Mashitah
School of Chemical Engineering, Universiti Sains Malaysia, Engineering Campus, 14300, Nibong Tebal, Malaysia
Journal of Applied Sciences
Year: 2010 | Volume: 10 | Issue: 24 | Page No.: 3307-3312







ABSTRACT
Effect of aeration (0.5, 1.0 and 1.5 vvm) and agitation rate (100, 300 and 500 rpm) on cellulase production in submerged culture of Pycnoporus sanguineus was studied in a 2.5 L stirred-tank bioreactor using Palm Oil Mill Effluent (POME) as a substrate. Maximum cell biomass (3.16 g L-1) and cellulase activity (0.1748 FPU mL-1) was obtained at aeration rate of 1.0 vvm and agitation speed of 300 rpm. Volumetric mass transfer coefficient (kLa) was found to be dependent on aeration and agitation rate, with maximum kLa (124.2 h-1) attained at 300 rpm and 1.5 vvm.
PDF Abstract XML References Citation
Received: June 06, 2010;
Accepted: July 06, 2010;
Published: October 19, 2010
How to cite this article
K. Fadzilah and M.D. Mashitah, 2010. Cellulases Production in Palm Oil Mill Effluent: Effect of Aeration and Agitation. Journal of Applied Sciences, 10: 3307-3312.
DOI: 10.3923/jas.2010.3307.3312
URL: https://scialert.net/abstract/?doi=jas.2010.3307.3312
DOI: 10.3923/jas.2010.3307.3312
URL: https://scialert.net/abstract/?doi=jas.2010.3307.3312
INTRODUCTION
Large amount of wastes in varying nature are generated by the Malaysian palm oil processing industry on a regular basis. Of particular concern is the Palm Oil Mill Effluent (POME), a High Biochemical Oxygen Demand (BOD) and Chemical Oxygen Demand (COD) liquid waste generated during downstream processing or extraction of crude palm oil. POME consists of 95-96% water, 0.6-0.7% oil and 4-5% total solids, half of which is suspended solid consisting of debris from the fruit. The high amount of degradable organic matter, which can cause severe pollution of waterways due to oxygen-depletion and other related effects, renders POME as an environmental hazard. Freshly discharged POME is acidic (pH 4-5) with temperatures around 80-90°C and contains appreciable amounts of plant nutrients (Zinatizadeh et al., 2006). POME is non-toxic in nature as no chemicals are added during oil extraction (Zinatizadeh et al., 2006; Mashitah et al., 2002).
It is estimated that around 5-7.5 ton of water are required to produce 1 ton of crude palm oil and more than 50% of the water end up as POME (Ahmad et al., 2003). Disposal of this voluminous liquid waste is expensive due to high costs of transportation and limited availability of landfills. Development of cost effective process for treatment and utilization of POME is therefore essential in order to improve the competitiveness of industrial processing operations and to minimize the environmental hazards posed by suspended solids-containing effluent (Thani et al., 1999). Substantial amount of work has been carried out with regard to the treatment of POME and its potential for biotechnological application, which saw various products being developed such as enzymes, bio-fuels and bio-polymers.
It has been reported that Palm Oil Mill Effluent (POME) could be used as a potential substrate for cellulases production (Mashitah et al., 2002; Mashitah, 2007). Cellulases have seen notable increase of interest in recent years due to their many potential applications. The far-reaching roles of cellulases can be seen in their diverse industrial scope, including food, animal feed, textile, fuel, chemical, pulp and paper, waste management, medical/pharmaceutical and pollution treatment (Bhat and Bhat, 1997). Growing demand for microbial cellulases and related enzymes saw cellulase becoming the third largest industrial enzyme worldwide by dollar volume in 2008 (Wilson, 2009). Cellulase production has been attempted on a wide range of substrates ranging from pure cellulose to dairy manure and agro-residues have been used frequently as carbon sources in cellulose fermentation. Most of these are capable of inducing cellulase system in fungi often at par with known inducers or sometimes even better (Mathew et al., 2008).
A complete cellulase system consists of three general classes of enzymes: cellobiohydrolases (CBH I and II), endoglucanases (EG I and II) and beta-glucosidase (BGL), which together act synergistically to degrade cellulose to glucose (Reczey et al., 1996). Cellulolytic enzyme systems are extensively studied in a wide variety of microorganisms, both complex and non-complex, including aerobic and anaerobic bacteria, white rot and soft rot fungi and anaerobic fungi, though only a few produce significant amount of extracellular cellulase complex. Fungal cellulases have proved to be a better candidate than other microbial cellulases, with their secreted free cellulase complexes comprising all three components of cellulase (Mathew et al., 2008). The best characterized and most widely studied cellulase system is that of the soft rot fungus Trichoderma, particularly T. reesei (Reczey et al., 1996). While, Trichoderma is known to produce high activities of overall cellulases, it has been established that they secrete low amounts of β-glucosidase, resulting in the accumulation of cellobiose, which is a strong inhibitor of the cellobiohydrolase activity (Jorgensen and Olsson, 2006). Much research has thus been directed towards finding new microbial sources of cellulase.
Pycnoporus sanguineus, a white-rot fungus commonly found in the Malaysian forest, is generally known to produce laccase as the sole or major lignolytic enzyme (Lu et al., 2008). To a lesser extent, P. sanguineus has also been found to produce α-amylase and extracellular cellulases depending on the carbon source used to grow the microorganism (Almeida et al., 1997). However, no studies have been reported on the utilization of this particular species on the production of cellulases using POME as a substrate. Thus, the present study aims to investigate the potential of partially treated POME for cellulases production through submerged cultivation of P. sanguineus.
MATERIALS AND METHOD
Microorganism: Pycnoporus sanguineus was obtained from the Forest Research Institute of Malaysia (FRIM). It was grown on potato dextrose agar at 30°C for 7-10 days and stored at room temperature until further use. Stock cultures were sub-cultured monthly.
Fermentation medium: POME was collected from the United Oil Palm (M) Sdn. Bhd., Penang, Malaysia. Raw POME was subjected to ultrasonic treatment (40 kHz, 50°C) for an hour and subsequently stored at 4°C until further use. Diluted POME (70% v/v) was used as fermentation media. Sigmacell cellulose Type 101 (10 g L-1) and peptone (5 g L-1) were added to the media as co-substrate/cellulase inducer and nitrogen source, respectively.
Cell suspension preparation: Cell suspension was prepared by suspending cell discs taken from 2 weeks-old P. sanguineus culture plate in standard bottle containing sterilized distilled water and a few drops of Tween 80. Using a 5 mm (o.d.) cork borer, the discs are taken by count of 10 discs per 100 mL of distilled water.
Inoculum preparation: Seed culture or inoculum was prepared by inoculating 15 mL of cell suspension into a 500 mL Erlenmeyer flask containing 135 mL of fermentation medium. The flask was incubated in a rotary shaker at 30°C, 250 rpm for 48 h.
Fermentation study: Batch fermentation was carried out in a 2.5 L bioreactor (Minifors, Infors HT, Switzerland) with a working volume of 1.5 L. The pH was maintained at 7.0±0.1 using 2 N HCl and 2 N KOH. Three agitation (100, 300 and 500 rpm) and aeration (0.5, 1.0 and 1.5 vvm) rates were studied. Fermentation was conducted for 7 days with samples taken at 12 h interval for the determination of cell biomass, cellulase activity and residual cellulose.
Analytical study: The samples taken from the bioreactor were centrifuged (4500x g) for 30 min to separate the cells and residual solids from the supernatant. The collected supernatant was analyzed for cellulase activity. Sediments were used to determine dry cell weight and residual cellulose.
Residual cellulose: Residual cellulose was determined by a shortened procedure of Updegraff (Ahamed and Vermette, 2009). Sediments from centrifuged sample were suspended in 3 mL acetic-nitric reagent and boiled for 30 min. It was centrifuged again prior to washing and drying the filtered residues at 40°C until constant weight. The resulting solids were mainly composed of cellulose.
Biomass: The fungal dry biomass was determined by measuring the solid dry weight. Mycelial weight was calculated from the difference between the total dry weight of the solids (comprising mycelium and cellulose) and that of the residual cellulose as determined in the previous section.
Cellulase assay: Cellulase assay with the culture supernatant of P. sanguineus was carried out in 0.05 M citrate buffer (pH 4.8) at 50°C. Filter paper (FPase) activity was determined as total cellulase activity by the method of Ghose (1987). One Filter Paper Unit (FPU) was defined as 1 μmol of glucose equivalent liberated per minute under the assay conditions.
Volumetric mass transfer coefficient: The overall kLa value (gas to cell) was determined after inoculation by the dynamic gassing out method (Nadeem et al., 2009). This technique monitors the change of oxygen concentration when air supply is turned off in an oxygen-saturated culture broth with respiring organism. Dissolved oxygen tension was then measured using a polarographic electrode. The mass balance in a vessel give,
| (1) |
integrating Eq. 1 results in
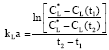 | (2) |
from which kLa can be calculated based on the measured values of CL(t1) and CL(t2).
RESULTS AND DISCUSSION
Dissolved oxygen tension and stirrer speed are important parameters for performing a desired fungal fermentation with a given strain and medium (Cui et al., 1998). The availability of oxygen is a major parameter to be considered for effective microbial cell growth rate; agitation is directly related to oxygen transported from the gas phase to liquid phase followed by oxygen uptake by individual microbial cell (Najafpour, 2007). Therefore, the effect of different aeration (0.5, 1.0, 1.5 vvm) and agitation rate (100, 300, 500 rpm) on growth of microorganism, substrate consumption, enzyme yield and volumetric mass transfer coefficient (kLa) were investigated in a 2.5 L stirred tank bioreactor.
Ultrasound as POME pre-treatment method: Ultrasonic treatment is one of the most promising technologies recently adopted in wastewater treatment plant, whereas application of ultrasound as a pretreatment step was found to enhance sludge biodegradability for subsequent aerobic oxidation (Sangave and Pandit, 2006; Laurent et al., 2009). As shown in our preliminary study in shake flask, culture cultivation of P. sanguineus in sonicated POME resulted in a 15% increase in cellulase activity [unpublished data]. It has been established by Sangave and Pandit (2006) and Wang et al. (2006) that low intensity ultrasound are sufficient to bring about transformation of the effluent constituents at a molecular level; sludge disintegration was reported to be predominant at lower frequencies (less than 100 kHz). The use of ultrasound resulted in cavitation phenomena, which is defined as the phenomena of the formation, growth and subsequent collapse of microbubbles or cavities (Sangave and Pandit, 2006). Several mechanisms have been suggested as to the effect of sonication on particles of a material in a liquid suspension, including: (1) surface erosion of particles due to cavitational collapse in the surrounding liquid, (2) particle size reduction due to fission caused by interparticle collisions and (3) the collapse of cavitation bubbles formed on the surface (Mason, 1998). As a result, hydro mechanical shear forces caused the disintegration of pollutant molecules into smaller fractions.
Effect of aeration rate: To determine the effects of aeration, the agitation rate was fixed at 300 rpm while aeration was varied (0.5, 1.0 and 1.5 vvm). It was observed that cell growth proceeded in two distinct stages: in the first two days, dry cell weight showed a rapid rise that was followed by slower growth (Fig. 1a). Dissolved Oxygen (DO) concentration, which was not controlled in all cultures, dropped rapidly during the first growth phase as oxygen demand was high (Fig. 1b). As growth slowed down in the second phase, the DO increased until it reached steady-state. This biphasic cell growth could be the result of rapid conversion of nutrients in the medium to cell mass, corresponding to the rapid growth in the first 2 days of culture. In this study, elevated aeration rate were accompanied by a corresponding increase in biomass, with highest growth (3.61 g L-1) observed at 1.0 vvm. As stated by Xu and Yun (2004), aeration results in better mixing of the exopolysaccharides from Paecilomyces tenuipes C240 in a stirred-tank fermenter. This in turn helps to maintain a concentration gradient between the interior and exterior of the cell, allowing better diffusion of nutrients to the cells. Increasing air flow rate further (1.5 vvm) saw cell dry weight dropping slightly. Reddy et al. (2002) reported that combining high flow rate with low agitation resulted in impeller flooding. This phenomenon occurred when the impeller is surrounded by air column, causing air flow pattern in the vessel to be dominated by air flow up the stirrer shaft. Improper contact between impeller and liquid then resulted in poor mixing, reduced air dispersion and diminished oxygen transfer rates. Culture broth exhibited non-Newtonian behavior for all aeration rates; broth became increasingly viscous during cell growth as a structured filamentous network was developed. Growth was still apparent at the end of fermentation period (day 7). As a wood-degrading fungus, P. sanguineus requires longer incubation time to reach stationary growth phase. However, this was not attempted due to time constraint. The heterogeneity of POME also presented a challenge in determining cell dry weight; direct determination of the cell dry weight was a problem due to the presence of suspended solids.
Enzyme activity and fungal biomass accumulated over the fermentation period suggested that there is a strong relationship between biomass formation and cellulase synthesis.
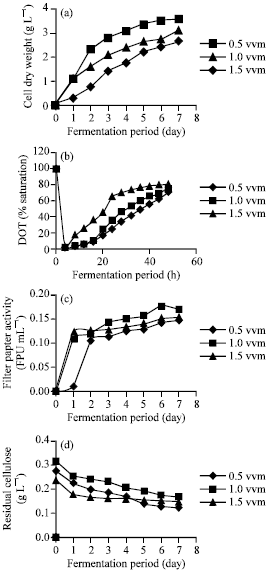 | |
| Fig. 1: | Effect of aeration rate on (a) cell dry weight, (b) dissolved oxygen tension (DOT), (c) cellulase activity and (d) residual cellulose |
Previous studies have reported that cellulase production is a growth-associated process (Mathew et al., 2008). Total cellulase activity, represented by filter paper (FPase) activity (Badhan et al., 2007), increased rapidly within the first 2 days of fermentation, following which cellulase activity was more or less stationary (Fig. 1c). Maximum cellulase activity (0.1748 FPU mL-1) was observed at 1.0 vvm after 6 days incubation. Enzymatic activity dropped at the seventh day of fermentation at 1.0 vvm aeration rate, possibly due to protease formation. Romero et al. (1999) reported that decrease in cellulase activity after a certain period could be due to protein hydrolysis by protease secreted by the fungus when there was a lack of essential nutrients in the medium. Cellulose consumption during cell growth, as shown by decreasing cellulose content (Fig. 1d) in the present work corroborated with this hypothesis.
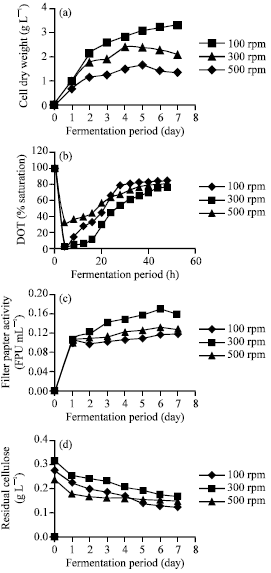 | |
| Fig. 2: | Effect of agitation rate on (a) cell dry weight, (b) dissolved oxygen tension (DOT), (c) cellulase activity and (d) residual cellulose |
Previous work by Agamuthu et al. (1986) also reported that aerobic digestion of POME decreased the carbon content.
Effect of agitation rate: Agitation is important for proper oxygen transfer and homogenous mixing of nutrients in fermentation system. To create an optimal environment in a bioreactor, agitation is required for cells to have access to all the substrates including oxygen in aerobic culture (Najafpour, 2007). To determine the effects of agitation, aeration rate was fixed at 1.0 vvm while stirrer speed was varied (100, 300 and 500 rpm). Similar pattern of two distinct growth phases as in the previous section were observed, although the difference in cell dry weight was found to be more pronounced by varying agitation rate. At 100 rpm, low cell growth was observed (Fig. 2a). This could be attributed to improper mixing which led to oxygen limitation, whereas death phase was observed after the fifth day of fermentation as limited dissolved oxygen in the fermentation media was used up. Cui et al. (1998) concluded that increasing agitation intensity enhances gas-liquid mass transfer as well as the mechanical forces that act on the fungal cells. The latter effect can be seen from the reduced cell growth at higher agitation (500 rpm), which could be attributed to shear stress and heterogenous mixing effects. Similar results were also reported in other studies (Nadeem et al., 2009). The 300 rpm of stirring speed was found to be most conducive for cell growth, whereas growth is still apparent towards the end of fermentation period at this rate. Agitation was also shown to be a factor for DO level, as they dropped rapidly during the first growth phase (Fig. 2b). As stated by Garcia-Ochoa et al. (2000), oxygen uptake rate increase during lag phase and especially during the exponential growth phase, followed by a constant rate during stationary growth phase.
Cellulase activity was also consistent with the growth trend, whereas a rapid increase in FPase activity was observed in the first 2 days of fermentation or during exponential growth phase. FPase activity increased as agitation speed was increased from 100 to 300 rpm. Further increase to 500 rpm saw cellulase activity decreasing. This may be the result of the formation of high shear zones close to the impellers caused by high impeller speed, with subsequent physical damage to the cells and a reduction in process productivity (Badino et al., 2001). Cellulase activity was also observed to decrease nearing the end of fermentation at both 300 and 500 rpm, possibly as a result of nutrient exhaustion and cell lysis. A study on cellulase deactivation in a stirred tank bioreactor by Gunjikar et al. (2001) concluded that: (1) the extent of deactivation increased with an increase in agitation speed, (2) the extent of deactivation for cellulase and its three components differed significantly and that (3) exoglucanase (FPase) contributes to the major decrease in cellulolytic activity in the initial stage of shearing.
Effect of aeration and agitation rate on volumetric mass transfer coefficient: Many factors are known to affect aeration efficiency including parameters such as agitation, air flow rate, air pressure, temperature, vessel geometry, fluid characteristics (density, viscosity, surface tension, etc.) and presence of antifoam agents (Ozbek and Gayik, 2001).
Overall oxygen transfer rate was assessed as the oxygen volumetric mass transfer coefficient (kLa). The effect of aeration and agitation rate on kLa was shown in Table 1 and 2, respectively. kLa values increased greatly when the air flow rate was increased up to 1.0 vvm.
| Table 1: | Volumetric mass transfer coefficient (kLa) at different aeration rate |
 | |
| Table 2: | Volumetric mass transfer coefficient (kLa) at different agitation rate |
 | |
At 1.5 vvm, however, kLa value was found to decrease. Although oxygen is necessary for the growth of aerobic cultures, high aeration rates probably disturb cell growth by damaging the cells. As stated by Nadeem et al. (2009), high aeration rate inhibited cell growth by creating air dispersion problem in the fermentation system which in turn affected cell biomass concentration. Elevating the stirring rate also increase the kLa values, although the increment was significantly lower than by varying aeration rate.
CONCLUSION
Submerged cultivation of P. sanguineus in ultrasonically-treated POME in a stirred tank bioreactor for the production of cellulase showed considerable potential with maximum cellulase production of 0.1728 FPU mL-1. Aeration and agitation rates were found to affect cell growth and cellulase production. Low aeration (0.5 vvm) and high agitation rate (500 rpm) adversely affected cell growth and cellulase production. Balance between aeration and agitation rates is thus essential for the optimum yield of cell biomass and cellulase production.
ACKNOWLEDGMENTS
The authors are grateful to the Federal Land Development Authority Foundation (Yayasan Felda) of Malaysia for their financial support (grant acc. No. 6050135) and Dr. Salmiah Ujang of Forest Research Institute of Malaysia (FRIM) for supplying the stock culture of P. sanguineus.
REFERENCES
- Ahamed, A. and P. Vermette, 2009. Effect of culture medium composition on Trichoderma reesei's morphology and cellulase production. Bioresour. Technol., 100: 5979-5987.
CrossRef - Ahmad, A.L., S. Ismail and S. Bhatia, 2003. Water recycling from Palm Oil Mill Effluent (POME) using membrane technology. Desalination, 157: 87-95.
CrossRefDirect Link - Badhan, A.K., B.S. Chadha, J. Kaur, H.S. Saini and M.K. Bhat, 2007. Production of multiple xylanolytic and cellulolytic enzymes by thermophilic fungus Myceliophthora sp. IMI 387099. Bioresour. Technol., 98: 504-510.
CrossRefPubMedDirect Link - Badino, Jr. A.C., M.C.R. Facciotti and W. Schmidell, 2001. Volumetric oxygen transfer coefficient (kLa) in batch cultivations involving non-Newtonian broths. Biochem. Eng. J., 8: 111-119.
CrossRef - Bhat, M.K. and S. Bhat, 1997. Cellulose degrading enzymes and their potential industrial applications. Biotechnol. Adv., 15: 583-620.
CrossRefDirect Link - Cui, Y.Q., J.N.W. Ouwehand, R.G.J.M. van der Lans and K.C.A.M. Luyben, 1998. Effects of dissolved oxygen tension and mechanical forces on fungal morphology in submerged fermentation. Biotechnol. Bioeng., 57: 409-419.
Direct Link - Almeida, M.S., K. Mizuta and J.R. Gigilo, 1997. Pycnoporus sanguineus: A novel source of α-amylase. Mycol. Res., 101: 188-190.
Direct Link - Garcia-Ochoa, F., E.G. Castro and V.E. Santos, 2000. Oxygen transfer and uptake rates during xanthan gum production. Enzyme Microbial Technol., 27: 680-690.
Direct Link - Ghose, T.K., 1987. Measurement of cellulase activities. Pure Applied Chem., 59: 257-268.
CrossRefDirect Link - Gunjikar, T.P., S.B. Sawant and J.B. Joshi, 2001. Shear deactivation of cellulase, exoglucanase, endoglucanase and β-glucosidase in a mechanically agitated reactor. Biotechnol. Prog., 17: 1166-1168.
Direct Link - Jorgensen, H. and L. Olsson, 2006. Production of cellulases by Penicillium brasilianum IBT 20888�Effect of substrate on hydrolytic performance. Enzyme Microbial Technol., 38: 381-390.
CrossRef - Laurent, J., M. Casellas, M.N. Pons and C. Dagot, 2009. Flocs surface functionality assessment of sonicated activated sludge in relation with physico-chemical properties. Ultrason. Sonochem., 16: 488-494.
Direct Link - Xu, C.P. and J.W. Yun, 2004. Influence of aeration on the production and the quality of the exopolysaccharides from Paecilomyces tenuipes C240 in a stirred-tank fermenter. Enzyme Microbial Technol., 35: 33-39.
CrossRef - Zinatizadeh, A.A.L., A.R. Mohamed, G.D. Najafpour, M. Hasnain Isa and H. Nasrollahzadeh, 2006. Kinetic evaluation of palm oil mill effluent digestion in a high rate up-flow anaerobic sludge fixed film bioreactor. Proc. Biochem., 41: 1038-1046.
CrossRefDirect Link - Mathew, G.M., R.K. Sukumaran, R.R. Singhania and A. Pandey, 2008. Progress in research on fungal cellulases for lignocellulose degradation. J. Sci. Indus. Res., 67: 898-907.
Direct Link - Nadeem, M., J.I. Qazi and S. Baig, 2009. Effect of aeration and agitation rates on alkaline protease production by Bacillus licheniformis UV-9 mutant. Turk. J. Biochem., 34: 89-96.
Direct Link - Reczey, K., Z.S. Szengyel, R. Eklund and G. Zacchi, 1996. Cellulase production by T. Reesei. Bioresour. Technol., 57: 25-30.
Direct Link - Reddy, V., P. Reddy, B. Pillay and S. Singh, 2002. Effect of aeration on the production of hemicellulase by T. lanuginosus SSBP in a 30 l bioreactor. Proc. Biochemi., 37: 1221-1228.
CrossRef - Romero, M.D., J. Aguado, L. Gonzalez and M. Ladero, 1999. Cellulase production by Neurospora crassa on wheat straw. Enzyme Microbial. Tech., 25: 244-250.
CrossRefDirect Link - Sangave, P.C. and A.B. Pandit, 2006. Ultrasound and enzyme assisted biodegradation of distillery wastewater. J. Environ. Manage., 80: 36-46.
Direct Link - Wang, F., S. Lu and M. Ji, 2006. Components of released liquid from ultrasonic waste activated sludge disintegration. Ultrason. Sonochem., 13: 334-338.
CrossRef