
S. Hamdan
Department of Industrial Biotechnology, Faculty of Biosciences and Bioengineering, Universiti Teknologi Malaysia, 81300 Skudai, Johor, Malaysia
F. Huyop
Department of Industrial Biotechnology, Faculty of Biosciences and Bioengineering, Universiti Teknologi Malaysia, 81300 Skudai, Johor, Malaysia
A.H. Zulkifly
Department of Biotechnology, Faculty of Agriculture and Biotechnology, University Sultan Zainal Abidin, 20400 Kuala Terengganu, Terengganu, Malaysia
D.D. Roslan
Department of Industrial Biotechnology, Faculty of Biosciences and Bioengineering, Universiti Teknologi Malaysia, 81300 Skudai, Johor, Malaysia
A.A.A. Hamid
Department of Industrial Biotechnology, Faculty of Biosciences and Bioengineering, Universiti Teknologi Malaysia, 81300 Skudai, Johor, Malaysia
S. Hamdan
Department of Industrial Biotechnology, Faculty of Biosciences and Bioengineering, Universiti Teknologi Malaysia, 81300 Skudai, Johor, Malaysia
F. Huyop
Department of Industrial Biotechnology, Faculty of Biosciences and Bioengineering, Universiti Teknologi Malaysia, 81300 Skudai, Johor, Malaysia
Journal of Applied Sciences
Year: 2010 | Volume: 10 | Issue: 22 | Page No.: 2940-2944







ABSTRACT
Haloacetic acids (HAAs) are group of chemicals formed due to disinfection by products that can be detected during chlorination and chloramination of processed drinking water. In this study, Bacillus sp. strain TW1 identified by morphological/biochemical and PCR-amplified 16S rRNA gene was isolated from Kuala Terengganu water treatment and distribution plant. TW1 was isolated due to its ability to grow in low concentration of monochloroacetic acid (MCA) of 0.5 mM, 10 times lower than normal MCA as sole carbon and energy source. Bacterial cell culture was grown in liquid minimal medium, pH 6.5 at 30°C on rotary shaker (150 rpm). Degradation of monochloroacetic acid was detected by measuring the amount of chloride ion released in the liquid minimal medium. Strain TW1 degraded monochloroacetic acid at best with maximum chloride ion released of 0.32 μmol Cl mL-1 using 0.5 mM MCA concentration. Current results demonstrated that this is the first reported study on low concentration of MCA degradation by Bacillus sp. TW1.
PDF Abstract XML References Citation
Received: June 30, 2010;
Accepted: August 19, 2010;
Published: October 11, 2010
How to cite this article
S. Hamdan, F. Huyop, A.H. Zulkifly, D.D. Roslan, A.A.A. Hamid, S. Hamdan and F. Huyop, 2010. Biodegradation of Low Concentration of Monochloroacetic Acid-Degrading Bacillus sp. TW1 Isolated from Terengganu Water Treatment and Distribution Plant. Journal of Applied Sciences, 10: 2940-2944.
DOI: 10.3923/jas.2010.2940.2944
URL: https://scialert.net/abstract/?doi=jas.2010.2940.2944
DOI: 10.3923/jas.2010.2940.2944
URL: https://scialert.net/abstract/?doi=jas.2010.2940.2944
INTRODUCTION
Most halogenated compounds are major environmental pollutants. Haloacetic acids (HAAs) are toxic organic chemicals that are frequently detected in surface waters and in drinking water distribution systems (McRae et al., 2004). They cause serious environmental pollution and human health problems as a result of their toxicity, persistence and transformation into hazardous metabolites. Most haloacetic acids (HAAs) are found throughout the biosphere due to high consumption of modern industrial and agricultural processes. Haloacetic acids are also used as herbicides and insecticides (Wilson and Mabury, 2000; Hashimoto et al., 2009). Haloacetic acids include many different compounds, such as monochloroacetic acid (MCA), dichloroacetic acid (DCA), trichloroacetic acid (TCA) and trifluoroacetic acids (TFA) (McRae et al., 2004; Ellis et al., 2001). Degradation of monochloroacetic acid (MCA), monobromoacetic acid (MBA) and fluoroacetic acid (FAA) have been reported by Olaniran et al. (2004), Sui-Yi et al. (2007), Donelly and Murphy (2009). Numerous studies have demonstrated that monochloroacetic acid (MCA) is toxic to aquatic life such as fishes and in particular to algae. These compounds, if present at high concentrations, are poisonous to plants and could lead to carcinogens (Hanson et al., 2002).
The destruction of organic chemicals by microorganisms may be influenced by environmental factors or the structure of the chemical itself. One of the reasons suggested for the lack of degradation of organic compounds by microbes is their low concentration (Boethling and Alexander, 1979). Microorganisms that metabolize and grow upon very low concentrations of substrates have been designated as oligotrophs (Poindexter, 1981). Such organisms appear to be adapted to low substrate concentrations by having high substrate affinity (low km value) systems. Biodegradation of very low concentrations of xenobiotic compounds has been neglected. However, it is useful to know about growth of microorganisms in low concentrations of pollutants because of the legal requirements. Normally this is set by a government to get the concentration of a pollutant down to a level that is not considered harmful. Therefore, if the microorganisms could only remove high concentration of pollutants, they could not be used to meet requirements of the law, since there still will be low concentrations of pollutants in the environment.
Current research focuses on the identification and partial characterization of potential bacteria from water treatment and distribution plants that can degrade low concentrations of MCA. Should growth be possible at low substrate concentrations, dehalogenase(s) km values reported earlier by Huyop et al. (2004) suggested that some dehalogenase(s) might function satisfactorily at low substrate concentrations.
MATERIALS AND METHODS
Chemicals: Various halogenated compounds of analytical grade were purchased from Sigma-Aldrich or Fluka. Bacterial agar from Difco and the rest of chemicals were of the highest purity commercially available.
Isolation and identification of organism: The bacterial isolate used in this study was originally isolated from Kuala Terengganu water treatment and distribution plant. About 5 mL of collected water sample were added into 250 mL shake flask liquid culture containing minimal salts medium (pH 6.5) with 5 mM MCA as the sole carbon source before sub-culturing onto a solid medium. After several sub-culture in 5 mM MCA solid minimal media (pH 6.5), one pure isolate was obtained by repeated streaking. The bacterial strain was characterized by morphological/biochemical tests as described in Bergey’s Manual of Systematic Bacteriology (Holt et al., 1994) and 16S rDNA gene analysis as described earlier (Hamid et al., 2010a, b).
Genomic DNA extraction: Bacterial isolates were either grown in liquid minimal medium supplemented with 5 mM of MCA or grown in liquid LB medium (Luria Bertani medium). Genomic DNA was then prepared using Wizard® Genomic DNA Purification Kit (Promega).
PCR amplification of 16S rRNA gene for bacteria identification: The primers used to amplify 16S rRNA were forward primer, FD1 (5’-aga gtt tga tcc tgg ctc ag-3’) and reverse primer, rP1 (5’-acg gtc ata cct tgt tac gac tt-3’) (Fulton and Cooper, 2005). The program used for amplification of 16S rRNA gene were: initial denaturation 94°C (5 min), denaturation 94°C (1 min), annealing 55°C (1 min) and extension 74°C (4 min).
Growth conditions: The culture was grown at 30°C on a rotary shaker in 250 mL flasks containing 100 mL medium as described earlier by Ismail et al. (2008). The liquid PJC minimal media was prepared as 10x concentrated basal salts containing K2HPO4. 3H2O (42.5 g L-1), NaH2PO4. 2H2O (10.0 g L-1) and (NH4)2SO4 (25.0 g L-1). The trace metal salts solution was a 10x concentrate that contained nitriloacetic acid (NTA) (1.0 g L-1), MgSO4 (2.0 g L-1), FeSO4. 7H2O (120.0 mg L-1), MnSO4. 4H2O (30.0 mg L-1), ZnSO4. H2O (30 mg L-1) and CoCl2 (10.0 mg L-1) in distilled water (Hareland et al., 1975). Minimal media for growing bacteria contained 10 mL of 10x basal salts and 10 mL of 10x trace metal salts per 100 mL of distilled water and were autoclaved (121°C, for 15 min).
The carbon source MCA was neutralized with NaOH and sterilized by filtration and added to the autoclaved salts medium to an appropriate final concentration. The extent of growth determined by measuring the absorbance at A680 nm and the release of chloride ions.
Measuring chloride ion released in the growth medium: Chloride ion released was detected in the growth medium by measuring the chloride ion at appropriate time intervals. Activity of the enzyme was measured by determining the release of chloride indicated by a colorimetric method employing mercuric thiocyanate as previously reported by Bergman and Sanik (1957). Samples (1.0 mL) were removed and assayed for halide ions. Each assay was carried out in triplicates. The absorbance of the mixture was measured at A460 nm and was proportional to the chloride ion concentration.
16S rRNA gene analysis: Following PCR, amplicons were purified using QIAGEN PCR purification kit. The amplicons were sent for DNA sequencing to 1st Base Laboratory, Malaysia using initial primers as described by Fulton and Cooper (2005). The partial 16S rRNA gene sequence was analyzed using BLASTn option (http://www.ncbi.nlm.nih.gov/BLAST/). The partial 16S rRNA gene sequence was submitted to NCBI under accession number HM598361.
RESULTS
Isolation of MCA degrading bacteria: A newly isolate bacterium strain TWI which degraded MCA as sole source of carbon was isolated from Kuala Terengganu water treatment and distribution plant. A single colony was observed on solid minimal media supplemented with 5 mM MCA (Fig. 1). The morphological, cultural and some physiological characteristics of the strain TWI suggested it belongs to Bacillus sp. (Table 1). According to Bergey’s Manual of Systematic Bacteriology, the strain TWI belongs to the genus Bacillus sp. The 16S rRNA gene fragment was also sequenced using FD1 and rP1 primers.
 | |
| Fig. 1: | A pure colonies (strain TW1) on solid minimal medium supplemented with 5 mM MCA incubated at 30°C after 16 h |
| Table 1: | Some morphological and general physiological properties of strain TWI |
 | |
The partial nucleotide sequence of 16S rRNA gene from the isolated bacterium also suggested that the species belongs to the genus Bacillus.
Growth of Bacillus sp. TW1 on 0.5 mM monochloroacetate (MCA): Bacillus sp. TW1 was confirmed to grow on MCA as sole source of carbon and energy. However, growth did not occur on MCA at concentrations in excess of 30 and 40 mM, suggesting toxicity of this compound to the organism (Zulkifly, 2008). The normal MCA concentration used to grow Bacillus sp. TW1 was 5 mM. To investigate the ability of strain TW1 to grow on low concentration of substrate, 0.5 mM MCA, 10x lower than the normal growth concentration was used.
An experiment was carried out to establish whether Bacillus sp. TW1 was able to grow on 0.5 mM MCA. The cell inoculum was prepared by growing TW1 in 10 mM glycolate minimal medium. Then 15 mL of an overnight culture was centrifuged and the cells washed twice with minimal medium before inoculation into 100 mL minimal medium supplied with 0.5 mM MCA. Growth was monitored by measuring the amount of chloride ions released at appropriate time intervals.
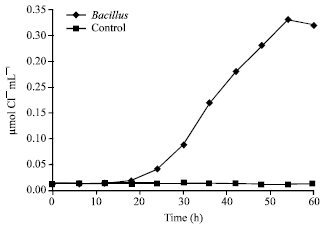 | |
| Fig. 2: | Growth of Bacillus sp. TW1 on 0.5 mM MCA |
An uninoculated flask treated in the same way was used as a control. This is important to make sure the chloride measured in the growth medium was due to the cells using the MCA rather than the auto-degradation of the substrate in the growth medium. A typical growth curve is shown in Fig. 2. with a doubling time of approximately 13 h. The maximum amount of chloride ion released detected in liquid culture was 0.32 μmol Cl¯ mL-1 from growth at 0.5 mM MCA. The chloride ion released indicated that TW1 strain consumed MCA. However, no chloride ion released was detected at 0.4 mM MCA concentration suggesting the affinity of the dehalogenase is not suitable under this condition.
DISCUSSION
The identification of Bacillus sp. TW1 was based on both biochemical tests/morphological observations and analysis of the 16S rRNA. The determination of 16S rRNA gene sequences is a routine procedure in prokaryotic taxonomy, resulting in large and growing databases, which improve phylogeny reconstructions, identification results and primer specificity evaluations (Nubel et al., 1997). The 16S rRNA gene sequence analysis confirmed that the organisms showing the highest identity are various Bacillus species. However, the analysis was not extensive enough to identify the species name of the organism.
According to Lane et al. (1985), in assessing the relationship of one organism to another by the comparison of their 16S rRNA sequence it is not important that the complete sequences be determined. The search in the database showed the sequence from the 5’ and 3’ matched the same organisms. The sequences from these organisms matched to the Bacillus suggesting the organism used in this study is from the genus Bacillus.
There is only one recent case associated with dehalogenation which involves the genus Bacillus. The dehalogenase from this organism was able to attack dichloromethane (Wu et al., 2009). Generally, it is quite rare for organisms from Bacillus sp. to be involved in hydrolytic dehalogenation. However, this kind of organism is always associated with bacteria of medical importance because they cause disease like food poisoning (From et al., 2007). Current investigation is the first reported study involving Bacillus genus able to degrade haloalkanoic acid especially at low substrate concentration of MCA.
This study strongly suggested that Bacillus sp. TW1 was able to grow on 0.5 mM MCA with a doubling time of approximately 13 h, similar to the growth rate obtained at 5 mM MCA concentration indicating that the organism was a facultative oligotroph (Zulkifly, 2008). However, growth above 30 mM MCA was inhibited possibly due to toxicity of MCA.
The identification of so many dehalogenases and the presence of multiple dehalogenases in many genera and species so far raise the question of their environmental significance. These enzymes are specifically induced by halo-acetic and propionic acids involving complex regulatory controls which respond to the growth environment. Some dehalogenases are specific and react to carbon-halogen bonds adjacent to the carboxyl group, whether at α or β-position of halogen group. Are these dehalogenases in natural environment will be expressed as dehalogenases or less specific hydrolases? The study of a complex regulatory control which responds to the growth environment may shed light on this question.
CONCLUSION
In conclusion, our research demonstrates that degradation of low MCA concentration was possible using strain TW1 in natural waters and in treated drinking waters. This is the first reported study on suppressing low concentration of MCA in drinking water so far which would help protect public health. Thus, in situ studies of MCA biodegradation in natural waters and drinking water distribution systems are important especially degradation at low concentration was possible.
ACKNOWLEDGMENT
This project was funded by Ministry of Higher Education (Malaysia) under Vot No. 78370.
REFERENCES
- Bergman, J.G. and J. Sanik, 1957. Determination of trace amounts of chlorine in naptha. Anal. Chem., 29: 241-243.
CrossRefDirect Link - Boethling, R.S. and M. Alexander, 1979. Effect of concentration of organic chemicals on their biodegradation by natural microbial communities. Applied Environ. Microbiol., 37: 1211-1216.
Direct Link - Donelly, C. and C.D. Murphy, 2009. Purification and properties of fluoroacetate dehalogenase from Pseudomonas fluorescens DSM 8341. Biotechnol. Lett., 31: 245-250.
CrossRefDirect Link - Ellis, D.A., M.L. Hanson, P.K. Sibley, T. Shahid and N.A. Fineberg et al., 2001. The fate and persistence of trifluoroacetic and chloroacetic acids in pond waters. Chemosphere, 42: 309-318.
CrossRef - From, C., V. Hormazabal and P.E. Granum, 2007. Food poisoning associated with pumilacidin-producing Bacillus pumilus in rice. Int. J. Food Microbiol., 115: 319-324.
PubMed - Fulton, C.K. and R.A. Cooper, 2005. Catabolism of sulfamate by Mycobacterium sp. CF1. Environ. Microbiol., 7: 378-381.
Direct Link - Hamid, A.A.A., S. Hamdan, S.H.Z. Ariffin and F. Huyop, 2010. Molecular prediction of dehalogenase producing microorganism using 16S rDNA analysis of 2,2-dichloropropionate (Dalapon) degrading bacterium isolated from volcanic soil. J. Biol. Sci., 10: 190-199.
CrossRefDirect Link - Hamid, A.A.A., S. Hamdan, Rolando V. Pakingking Jr. and F. Huyop, 2010. Identification of Pseudomonas sp. strain S3 based on small subunit ribosomal RNA gene sequences. Biotechnology, 9: 33-40.
CrossRefDirect Link - Hanson, M.L., P.K. Sibley, D.A. Ellis, S.A. Mabury, D.C. Muir and K.R. Solomon, 2002. Evaluation of monochloroacetic acid (MCA) degradation and toxicity to Lemna gibba, Myriophyllum spicatum and Myriophyllum sibiricum in aquatic microcosms. Aquat. Toxicol., 61: 251-273.
PubMed - Hareland, W.A., R.L. Crawford, P.J. Chapman and S. Dagley, 1975. Metabolic function and properties of a 4-hydroxyphenylacetic acid 1-hydroxylase from Pseudomonas acidovorans. J. Bacteriol., 121: 272-285.
Direct Link - Hashimoto, S., T. Azuma and A. Otsuki, 2009. Distribution, sources and stability of haloacetic acids in Tokyo Bay, Japan. Environ. Toxicol. Chem., 17: 798-805.
CrossRefDirect Link - Ismail, S.N., A.M. Taha, N.H. Jing, R.A. Wahab, A.A. Hamid, R.V. Pakingking Jr. and F. Huyop, 2008. Biodegradation of monochloroacetic acid by a presumptive Pseudomonas sp. strain R1 bacterium isolated from Malaysian paddy (rice) field. Biotechnology, 7: 481-486.
CrossRefDirect Link - Sui-Yi, K., A.F.M. Siu, S.M. Ngai, C.M. Che and J.S.H. Tsang, 2007. Proteomic analysis Burkholderia cepacia MBA4 in the degradation of monochloroacetate. Proteomics, 7: 1107-1116.
CrossRefDirect Link - Lane, D.J., B. Pace, G.J. Olsen, D.A. Stahl, M.L. Sogin and N.R. Pace, 1985. Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. Proc. Natl. Acad. Sci. USA., 82: 6955-6959.
PubMedDirect Link - McRae, B.M, T.M. LaPara and R.M. Hozalski, 2004. Biodegradation of haloacetic acids by bacterial enrichment cultures. Chemosphere, 55: 915-925.
CrossRef - Nubel., U, F. Garcia-Pichel and G. Muyzer, 1997. PCR primers to amplify 16S rRNA genes from cyanobacteria. Applied Environ. Microbiol., 63: 3327-3332.
Direct Link - Olaniran, A.O., D. Pillay and B. Pillay, 2004. Haloalkane and haloacid dehalogenases from aerobic bacterial isolates indigenous to contaminated sites in Africa demonstrate diverse substrate specificities. Chemosphere, 55: 27-33.
CrossRef - Poindexter, J.S., 1981. Oligotrophy: Fast and famine existence. Adv. Microb. Ecol., 5: 63-89.
Direct Link - Wilson, R.I. and S.A. Mabury, 2000. Photodegradation of metolachlor: Isolation, identification and quantification of monochloroacetic acid. J. Agric. Food Chem., 48: 944-950.
CrossRefDirect Link - Wu, S., H. Zhang, X. Yu and J. Chen, 2009. Identification and cloning of a gene encoding dichloromethane dehalogenase from a methylotrophic bacterium, Bacillus circulans WZ-12 CCTCC M 207006. Bioprocess Biosyst. Eng., 32: 845-852.
CrossRefDirect Link