
T.M. San
Jeffrey Cheah School of Medicine and Health Sciences, Monash University Sunway Campus, Jalan Lagoon Selatan, Bandar Sunway, 46150, Selangor Darul Ehsan, Malaysia
J. Vejayan
Jeffrey Cheah School of Medicine and Health Sciences, Monash University Sunway Campus, Jalan Lagoon Selatan, Bandar Sunway, 46150, Selangor Darul Ehsan, Malaysia
K. Shanmugan
Jeffrey Cheah School of Medicine and Health Sciences, Monash University Sunway Campus, Jalan Lagoon Selatan, Bandar Sunway, 46150, Selangor Darul Ehsan, Malaysia
H. Ibrahim
Institute of Biological Sciences, University of Malaya, 50603 Kuala Lumpur, Malaysia
Journal of Applied Sciences
Year: 2010 | Volume: 10 | Issue: 19 | Page No.: 2328-2332







ABSTRACT
The study was carried out to screen antimicrobial activity in venoms obtained from 11 species of snakes that are all common in Malaysia. Snake venom, which constitutes a diverse range of proteins, has long been identified as a potential source of therapeutics. Therefore, in times when antimicrobial resistance is becoming an increasingly severe issue, it is unsurprising that snake venoms are being investigated for antimicrobial components. Antibacterial activity was assessed using the hole-plate method. Venoms of Calloselasma rhodostoma and Ophiophagus hannah were capable of producing the most prominent bacterial inhibition zones with maximum values as high as 12 mm, while the other venoms screened only produced bacterial inhibition zones that were not more than 10 mm. These two venoms were selected for further determination of Minimum Inhibitory Concentration (MIC). The MIC values were tested with Staphylococcus aureus ATCC25923, ATCC29213 and ATCC43300. The MIC values obtained for Calloselasma rhodostoma were 125 μg mL-1 when tested against S. aureus ATCC25923 and ATCC43300, while it was 250 μg mL-1 when tested against S. aureus ATCC29213. MIC values obtained for Ophiophagus hannah were 250 μg mL-1 when tested against all three strains. Since the potential of snake venoms for antimicrobial activity has been established, further study is in the progress to purify the active antimicrobial component and to screen a wider range of bacterial strains.
PDF Abstract XML References Citation
Received: February 07, 2010;
Accepted: May 03, 2010;
Published: July 27, 2010
How to cite this article
T.M. San, J. Vejayan, K. Shanmugan and H. Ibrahim, 2010. Screening Antimicrobial Activity of Venoms from Snakes Commonly Found in Malaysia. Journal of Applied Sciences, 10: 2328-2332.
DOI: 10.3923/jas.2010.2328.2332
URL: https://scialert.net/abstract/?doi=jas.2010.2328.2332
DOI: 10.3923/jas.2010.2328.2332
URL: https://scialert.net/abstract/?doi=jas.2010.2328.2332
INTRODUCTION
Resistance of bacteria against antimicrobial agents has always been an immense burden to healthcare worldwide. High resistance of Staphylococcus aureus against ciprofloxacin, amoxicillin and chloramphenicol has been documented in a study (Nafeesa et al., 2001). The problem has led to wider search for antimicrobial agents from other sources against the antibiotics resistance bacterial strains. Among these new sources are plant extracts, which have shown promising activities against methicillin resistant S. aureus (MRSA), vancomycin resistant S. aureus (VRSA) and vancomycin intermediate S. aureus (VISA) (Dadgar et al., 2006; Zakaria et al., 2007). Snake venom is another new source of antimicrobial agents that we will describe in this study.
Snake venoms constitute a diverse range of proteins that include neurotoxins, enzymes and peptides. Consequently, the idea of harvesting proteins from snake venoms to be developed as commercial, therapeutic products has not been new.
Given its vast potential as a source of therapeutics, it is, therefore, unsurprising that snake venoms have also been investigated for antimicrobial components. Despite heavy colonization of pathogenic bacteria in the oral cavity of the snake, snake bite victims are not frequently observed to suffer from wound infection. This observation led to the theory that snake venom may have antimicrobial activity (Talan et al., 1991). While, the significance of antimicrobial component in snake venoms is yet to be fully elicited, it has been hypothesized that these components in the oral secretions of the snake have been developed under evolutionary pressure as a defense mechanism of the snake against the microorganisms on its prey (Shivik, 2006).
Among some of the common antimicrobial components that have been isolated from snake venoms are the enzymes L-amino oxidase (LAAO) and phospholipase A2 (PLA2). The LAAO purified from Pseudechis australis showed significant antimicrobial activity against different strains of Aeromonas (Stiles et al., 1991). PLA2 purified from Crotalus durissus terrificus and Daboia russelli siamensis have shown antibacterial activity against the Gram-negative bacteria Burkholderia pseudomallei (Samy et al., 2006). PLA2 isolated from Agkistrodon halys also showed significant inhibition against Staphylococcus aureus, Proteus vulgaris and Proteus mirabilis (Samy et al., 2008).
Apart from enzymatic proteins, antimicrobial peptides have also been purified in recent studies. Cathelicidins isolated from Bungarus fasciatus and Ophiophagus hannah have reported potent antimicrobial activity against many strains of Gram negative bacteria (Wang et al., 2008; Chen et al., 2009). A small peptide vgf-1, isolated from Naja atra, has shown to have antimycobacterial activity (Xie et al., 2003). Omwaprin, purified from Oxyuranus microlepidotus, is reported to show antimicrobial activity against Gram positive bacteria such as Staphylococcus wagneri and Bacillus megaterium (Nair et al., 2007).
In this study, we describe the preliminary screening of 11 snake venoms for antibacterial activity against three different strains of Staphylococcus aureus and two Gram negative bacteria, Pseudomonas aeruginosa and Escherichia coli. Similar screening focusing on antimicrobial property has not been attempted previously among indigenous Malaysian snakes.
MATERIALS AND METHODS
Venoms: All venoms used were of common venomous snakes in Malaysia, obtained from the snake farm in Sungai Batu Pahat, Perlis, Malaysia, in the year of 2009. The venoms were all, freeze-dried and stored in -20°C.
Bacterial strains: The bacterial strains used in the antibacterial screening assays were Staphylococcus aureus ATCC25923, Staphylococcus aureus ATCC29213, Methicillin-resistant Staphylococcus aureus (MRSA) ATCC43300, Pseudomonas aeruginosa ATCC25873 and Escherichia coli ATCC25922. The bacteria were streaked onto Mueller-Hinton (MH) agar plates, incubated overnight for growth and stored at 4°C.
Bacterial inoculum: Bacterial inoculum that would be used for antibacterial screening assays was prepared using the standard method of log-phase growth. A single-unit colony was picked from the bacterial culture plates and added to 10 mL of Mueller-Hinton (MH) broth. The inoculated MH broth was incubated at 37°C until a colony-forming unit (CFU) mL-1 value of 105. This was confirmed by measuring the absorbance value of the inoculum with the spectrophotometer (Shimadzu) to give A600=0.1.
Antibacterial screening assays: Lyophilized venoms (1 mg mL-1) were prepared in deionized water. Three milliliter of bacterial inoculum (cfu mL-1 = 105, A600 = 0.1), was spread onto an agar plate and left standing for 3 min. Excessive inoculum was poured away and the agar plate was left standing for 3 min again. Using a modified hole-plate method, 4 mm diameter wells were made onto the agar using an immunodiffusion cutter (Nair et al., 2007). Thirty microliter of each dissolved venom sample was loaded into each well. Vancomycin and methicillin (30 μg mL-1) were used as drug controls. The bacterial plates were incubated at 37°C for 18 h and the inhibition zones were measured in millimetre diameters.
Minimum inhibitory concentration: Minimum Inhibitory Concentration (MIC) was determined using a modified method described by Nair et al., 2007. The two venoms which showed the highest diameter of inhibition zones were chosen for MIC determination. Crude venom of concentration 1 mg mL-1 was serially diluted in the range of 0.5, 0.25, 0.125, 0.0625 and 0.03125 mg mL-1 using cation-adjusted MH broth. Three milliliter of bacterial inoculum was spread onto 30 mL of MH agar as described above for the antibacterial screening assays. Six wells of diameter 4 mm were made onto the agar plate. Each well was loaded with 30 μL of one concentration of the diluted venom. The plates were incubated at 37°C for 18 h and MIC was defined as the lowest concentration of venom which gave a detectable inhibition zone.
RESULTS
Table 1 shows the antibacterial activity of 11 crude venoms tested against five different strains of bacteria, three different strains of S. aureus and two different strains of Gram negative bacteria. From this preliminary screening, the Gram negative bacteria were found to be resistant against all the crude venoms as no inhibition zone was exhibited in the antibacterial assay.
The antibacterial activities of the crude venom against the Gram positive bacteria were found to be more significant among the Viperidae species. Crude venoms from all four Viperidae species demonstrated significant inhibition zones between 6.6-12.5 mm. Calloselasma rhodostoma showed the largest inhibition zones between of 10.2-12.5 mm.
Among the crude venoms from the six Elapidae species, only three exhibited antibacterial activity. The inhibition zones of Bungarus candidus and Bungarus fasciatus demonstrated against the three strains of S. aureus were only between 6.3-8.0 mm. Ophiophagus hannah was the only exception among the Elapidae species, exhibiting large inhibition zones between 9.5-11.5 mm.
| Table 1: | Antibacterial activity of 11 crude venoms against five strains of bacteria tested using hole-plate method |
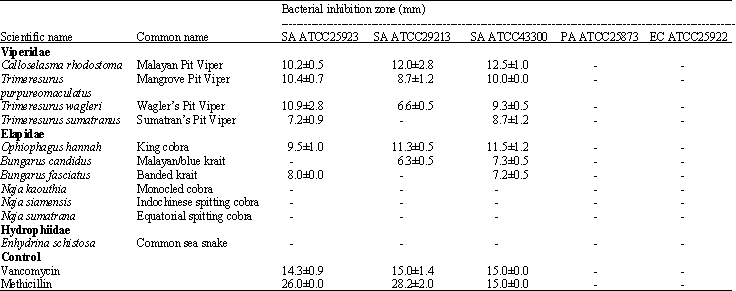 | |
The values are presented as Mean±SD (n = 4). The inhibition zones were measured after 18 h of incubation at 37°C and the inhibition zone diameters include the 4 mm diameters of the wells containing either 30 μL of crude venom (1 mg mL-1) or antibiotic (30 μg mL-1). Bacterial inoculum of 105 cfu mL-1 (A600 = 0.1) is added per plate. SA: Staphylococcus aureus; PA: Pseudomonas aeruginosa; EC: Escherichia coli. -: No activity | |
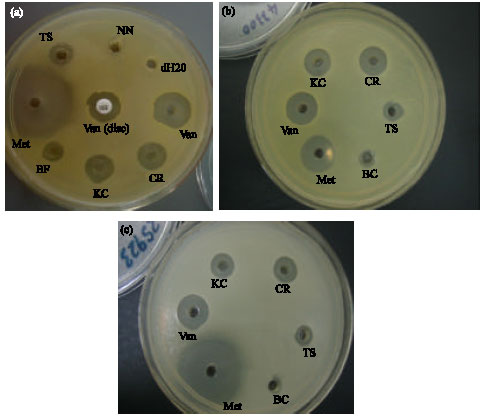 | |
| Fig. 1: | Antibacterial assays of crude venom using the hole-plate method. (a) SA ATCC29213, (b) SA ATCC25923 and (c) SA ATCC43300. SA: S. aureus; KC: O. hannah (King Cobra); CR: C. rhodostoma; TS: T. sumatranus; BC: B. candidus; BF: B. fasciatus; NN: N. kaouthia; Van, Vancomycin; Met, Methicillin. A vancomycin disc (30 μg mL-1) was also used against SA ATCC29213, as shown in (1a) in comparison with diluted vancomycin solution (30 μg mL-1). The diluted vancomycin showed higher inhibition diameter than the vancomycin disc |
The venom of the single Hydrophiidae species screened in this study exhibits no antibacterial activity.
The crude venoms also have lower antibacterial activity as compared to that of the conventional antibiotics, as shown by the larger inhibition zones of vancomycin and methicillin.
Figure 1a-c showed the antibacterial assays of the various crude venoms tested against the three strains of S. aureus ATCC29213, ATCC25923 and ATCC43300 using the hole-plate method.
| Table 2: | MIC values of C. rhodostoma, O. hannah and antibiotics |
 | |
MIC was determined as the lowest concentration of venom which gave a detectable inhibition zone after 18 h of incubation at 37°C. The venoms were diluted in the range of 1, 0.5, 0.25, 0.125, 0.0625 and 0.03125 mg mL-1. Each concentration was added to one well. Bacterial inoculum of 105 cfu mL-1 (A600 = 0.1) is added per plate. SA, Staphylococcus aureus | |
In all three assays shown, it could be seen that venoms from Calloselasma rhodostoma and Ophiophagus hannah demonstrated clear inhibition zones that were significantly larger than the other crude venoms screened.
Since the crude venoms from Calloselasma rhodostoma and Ophiophagus hannah demonstrated the most significant inhibition zones against the three strains of S. aureus, including the MRSA strain, these two venoms were selected for MIC determination. Table 2 shows the MIC values of Calloselasma rhodostoma and Ophiophagus hannah. The MIC values of both venoms were found to be within the range of 125-250 μg mL-1. When tested against S. aureus ATCC25923 and ATCC43300, Calloselasma rhodostoma demonstrated lower MIC as compared to Ophiophagus hannah. The MIC values of these two venoms were found to be much higher than that of the antibiotics.
DISCUSSION
One of the main patterns that can be identified in this preliminary screening is that the Viperidae venoms exhibited larger inhibition zones than Elapidae venoms, with the exception of Ophiophagus hannah. This pattern is very similar to the pattern found when enzymatic properties of Viperidae and Elapidae venoms were compared, which showed that venoms of Viperidae species had higher enzymatic activities than venoms of Elapidae species, with the exception of Ophiophagus hannah venom, an Elapidae venom that showed high enzymatic activities (Kocholaty et al., 1971). The results of this preliminary screening, therefore, highly suggest that the antibacterial activity of snake venoms is due to enzymatic components. This is also in tandem with other findings, which have shown that snake venom antimicrobial activity is due to enzymes such as PLA2 (Samy et al., 2008).
Another pattern that can be identified from this preliminary screening is the susceptibility of the three strains of Gram-positive S. aureus to the crude venoms and the resistance of the two strains of Gram negative bacteria against the venoms.
Similar patterns of dose-dependent species specific susceptibility and resistance against Gram negative have also been noted in omwaprin, a peptide isolated from the venom of Oxyuranus microlepidotus. Omwaprin is effective against Gram positive bacteria such as Bacillus megaterium and S. wagneri, but shows resistance when tested against Gram negative bacteria and other Gram positive bacteria such as B. thuringiensis, S. aureus and Streptococcus clavuligerus (Nair et al., 2007). The resistance against Gram negative bacteria could possibly be due the outer membrane of the bacteria. The outer membrane of Gram negative bacteria has lipopolysaccharides (LPS). Charges on the LPS molecule can affect the uptake of antimicrobial peptide and such similar mechanism might have resulted in the resistance we observed among the Gram negative bacteria tested in this screening (Devine and Hancock, 2002). It has also been observed that Gram negative bacteria, when treated with EDTA that disrupts the outer membrane, become susceptible to antibacterial agents such as bronchochin (Gao et al., 1999).
Nevertheless, this preliminary screening is by no means representative and conclusive of the resistance of Gram negative bacteria against snake venoms, as the number of bacterial strains used is very minimal. Moreover, PLA2 and cathelicidin previously isolated from other snake venoms were found to be effective against Gram negative bacteria such as Burkholderia pseudomallei and E. coli (Samy et al., 2006; Wang et al., 2008), contradicting our findings from this screening. Species specific susceptibility also could not be identified from this screening as only one type of Gram positive bacterium has been used.
While it has been observed with electron microscopy that the antibacterial mechanism of snake venoms involves cell surface membrane blebbing and subsequent cell contents leakage (Nair et al., 2007; Samy et al., 2008) the exact mechanism of the cell membrane disruption leading to cell death is still very much unclear. Snake venom antimicrobial mechanism is complex and is affected by factors such as amino acid sequence, net charge of the protein, three-dimensional structure, bacterial membrane composition and salinity of the environment (Nair et al., 2007). Hence, the different proteins/peptides from different species of snakes can have varied mechanisms of cell membrane disruption, resulting in the differences of susceptibility among the different bacterial strains.
While the MIC values obtained from this screening is many times higher than that of the conventional antibiotics, it has been shown in a comparison study that purified LAAO exhibited significantly higher inhibition than crude venoms (Samy et al., 2007). The inhibition exhibited by fractions obtained from each stage of purification also increases (Samy et al., 2008).
In summary, we have successfully established the potential of antimicrobial activity using snake venom and have also identified two promising venoms, Calloselasma rhodostoma and Ophiophagus hannah, which show the highest inhibition. Further study is in the progress to purify the active antimicrobial proteins/peptides from these two venoms, as well as to screen a larger number of bacteria to characterize the susceptibility of these two venoms more accurately.
ACKNOWLEDGMENTS
Authors would like to thank Ministry of Science, Technology and Innovation (MOSTI), Malaysia for funding under EScience grant: 02-02-10-SF0033 and Monash University Sunway Campus for funding under Internal grant: MED2010-INI(MG)-001-JV.
REFERENCES
- Dadgar, T., M. Asmar, A. Saifi, M. Mazandarani and H. Bayat et al., 2006. Antibacterial activity of certain iranian medicinal plants against methicillin-resistant and sensitive Staphylococcus aureus. Asian J. Plant Sci., 5: 861-866.
CrossRefDirect Link - Nafeesa, A., M.A. Sheikh, Ikram-ul-Haq, A. Jamil and Z. Parveen, 2001. Microbial resistance of Staphylococcus aureus against commonly used antibiotics. J. Medical Sci., 1: 97-100.
CrossRefDirect Link - Nair, D.G., B.G. Fry, P. Alewood, P.P. Kumar and R.M. Kini, 2007. Antimicrobial activity of omwaprin, a new member of the waprin family of snake venom proteins. Biochem. J., 402: 93-104.
CrossRefDirect Link - Samy, R.P., A. Pachiappan, P. Gopalakrishnakone, M.M. Thwin and Y.E. Hian et al., 2006. In vitro antimicrobial activity of natural toxins and animal venoms tested against Burkholderia pseudomallei. BMC Infect. Dis., 6: 100-100.
PubMed - Samy, R.P., P. Gopalakrishnakone, H. Bow and V.T.K. Chow, 2008. Purification, characterization and bactericidal activities of basic phospholipase A2 from the venom of Agkistrodon halys (Chinese pallas). Biochimie, 90: 1372-1388.
PubMed - Samy, R.P., P. Gopalakrishnakone, M.M. Thwin, T.K.V. Chow, H. Bow, E.H. Yap and T.W.J. Thong, 2007. Antibacterial activity of snake, scorpion and bee venoms: A comparison with purified venom phospholipase A2 enzymes. J. Appl. Microbiol., 102: 650-659.
CrossRefPubMedDirect Link - Shivik, J.A., 2006. Are vultures birds and do snakes have venom, because of macro-and microscavenger conflict?. BioScience, 56: 819-823.
Direct Link - Stiles, B.G., F.W. Sexton and S.A. Weinstein, 1991. Antibacterial effects of different snake venoms: Purification and characterization of antibacterial proteins from Pseudechis australis (Australian King Brown or mulga snake) venom. Toxicon, 29: 1129-1141.
PubMed - Talan, D.A., D.M. Citron, G.D. Overturf, B. Singer, P. Froman and E.J.C. Goldstein, 1991. Antibacterial activity of crotalid venoms against oral snake flora and other clinical bacteria. J. Infect. Dis., 164: 195-198.
Direct Link - Wang, Y., J. Hing, X. Lai, H. Yang and R. Liu et al., 2008. Snake cathelicidin from Bungarus fasciatus is a potent peptide antibiotics. PLoS ONE, 3: e3217-e3217.
CrossRefDirect Link - Xie, J.P., J. Yue, Y.L. Xiong, W.Y. Wang, S.Q. Yu and H.H. Wang, 2003. In vitro activities of small peptides from snake venom against clinical isolates of drug-resistant Mycobacterium tuberculosis. Int. J. Antimicrobol. Agents, 22: 172-174.
PubMedDirect Link - Zakaria, Z.A., A.M.M. Jais, M. Mastura, S.H.M. Jusoh and A.M. Mohamed et al., 2007. In vitro antistaphylococcal activity of the extract of several neglected plants in Malaysia. Int. J. Phamacol., 3: 428-431.
Direct Link - Gao, Y., M.J. van Belkum and M.E. Stiles, 1999. The outer membrane of Gram-negative bacteria inhibits antibacterial activity of Brochocin-C. Applied Environ. Microbiol., 65: 4329-4333.
PubMedDirect Link - Chen, X.X., G.Y. Yu, Y. Zhan, Y. Zhang, J.H. Shen and W.H. Lee, 2009. Effects of the antimicrobial peptide OH-CATH on Escherichia coli. Zool. Res., 30: 171-177.
CrossRef - Kocholaty, W.F., E.B. Ledford, J.G. Daly and T.A. Billings, 1971. Toxicity and some enzymatic properties and activities in the venoms of crotalidae, elapidae and viperidae. Toxicon, 9: 131-138.
PubMed