
B. Akbar John
Institute of Oceanography and Maritime Studies, International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
K.C.A. Jalal
Institute of Oceanography and Maritime Studies, International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
Y.B. Kamaruzzaman
Institute of Oceanography and Maritime Studies, International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
K. Zaleha
Institute of Aquatrop, Universiti Malaysia Terengganu, 21030 Kuala Terengganu, Terengganu, Malaysia
Journal of Applied Sciences
Year: 2010 | Volume: 10 | Issue: 17 | Page No.: 1930-1936







ABSTRACT
Endotoxins from gram-negative microorganisms initiates clot formation in blood when it is accidentally encountered by horseshoe crab. This property was extensively studied by various researchers as a result Limulus Amebocyte Lysate (LAL) test was established. The LAL tests in general were 3 to 300 times more sensitive than the United States Pharmacopeial (USP) rabbit pyrogen test method. It should be noted that major differences among the LAL preparations lie in the area of sensitivity. Differences, up to 100-fold, exist in the sensitivity of the various LAL preparations to the same endotoxin. It is also considerate that the Information on the molecular level interaction between horseshoe crab blood and endotoxin would provide detailed information about the clotting process. Based on the above perspective review of literature was established to elucidate the compounds involved in the clot formation in horseshoe crab blood while encountering with bacterial endotoxin. Since LAL preparation is important for its application in clinical, pharmaceutical, public health and environmental areas, this paper will also discuss some molecular level details in Lysated Amebocyte interaction in clot formation.
PDF Abstract XML References Citation
Received: February 05, 2010;
Accepted: April 15, 2010;
Published: July 10, 2010
How to cite this article
B. Akbar John, K.C.A. Jalal, Y.B. Kamaruzzaman and K. Zaleha, 2010. Mechanism in the Clot Formation of Horseshoe Crab Blood during Bacterial Endotoxin Invasion. Journal of Applied Sciences, 10: 1930-1936.
DOI: 10.3923/jas.2010.1930.1936
URL: https://scialert.net/abstract/?doi=jas.2010.1930.1936
DOI: 10.3923/jas.2010.1930.1936
URL: https://scialert.net/abstract/?doi=jas.2010.1930.1936
INTRODUCTION
Horseshoe crabs are at the epicenter of one of the most interesting marine resource Management issues along the coast. They are marine chelicerate arthropod. Despite their name, they are more closely related to spiders, ticks and scorpions than to crabs. There are four extant species of horseshoe crabs, Tachypleus tridentatus, Tachypleus gigas, Carcinoscorpius rotundicauda and Limulus polyphemus The first three species inhabit the Southeast Asian coast and the Limulus polyphemus inhabits in the east coast of North America (Chliboyko and Ancestors, 2008; Kassim et al., 2008).
Horseshoe crabs have been used in eye research in biomedical industries for the manufacture of toxin free surgical sutures and for the development of wound dressings for burn victims. But perhaps most important is the use of a component of the horseshoe crab’s blood called Limulus Amebocyte Lysate (LAL), which is indispensable for the detection of bacterial contamination in drugs and intravenous devices. Endotoxin is a poisonous substance that is found in the cell wall of Gram negative bacteria which can withstand steam sterilization and it is made up of lipopolysaccharide and lipoprotein complexes. The horseshoe crab lacks an immune system; it cannot develop antibodies to fight infection. However the horseshoe crab does contain a number of compounds that will bind to and inactivate bacteria, fungi and viruses. The components of LAL are part of this primitive immune system. The components in LAL for example, not only bind and inactivate bacterial endotoxin, but the clot formed as a result of activation by endotoxin provides wound control by preventing bleeding and forming a physical barrier against additional bacterial entry and infection. It is one of the marvels of evolution that the horseshoe crab uses endotoxin as a signal for wound occurrence and as an extremely effective defense against infection.
A unique and intriguing feature of this ancient species is that it has blue copper-based blood (Shuster, 2004). About 37 years, the rabbit pyrogen test was the only practical pyrogen procedure described in regulatory compendia throughout the world (Addendum, 1989; Anonymous, 1983; Burdon-Sanderson, 1876; Dubczanski and Naunyn, 1873; Hort and Penfold, 1912; Kristansen, 1984; Levin and Bang, 1964a, b; Panum, 1874). Although, it has met its purposes relatively well over the years, it remains an elaborate procedure subject to the variability inherent in all biological assays. The need for a simple, specific and accurate pyrogen test has long been recognized. Although a number of test systems have been devised for endotoxin detection, only the Limulus Amebocyte Lysate (LAL) Test is potentially a satisfactory alternative to the USP rabbit pyrogen test. With the advent of a sensitive, specific, inexpensive and rapid assay for endotoxin, a number of workers investigated its potential use in the pharmaceutical industry as an alternate to the rabbit pyrogen test. The development of a viable alternative to the rabbit pyrogen test, began with the innovative work of John Hopkins University investigators Levin and Bang (Prior and Spagna, 1979) observed that bacterial infection causes intravascular coagulation in the American Horseshoe crab. Levin and Bang further reported that the agent responsible for clotting phenomenon resided inside the amebocytes (blood cells) for Horseshoe crab. Bacterial endotoxin was found to be responsible for initiating this gelation, characterized by an enzymatic gelation reaction. The LAL test is derived from the blue, copper-based blood of the horseshoe crab which is helpful in detecting as little as on million of a billionth of a gram of endotoxin (Mikkelsen, 1988).
HEMOLYMPH OF HORSESHOE CRAB
The limulus hemolymph contains two types of cells, granular hemocytes and cyanocytes (Dumont et al., 1966; Murer et al., 1975; Ornberg and Reese, 1981; Armstrong and Rickles, 1982; Toh et al., 1991). Based on cell morphology, there appears to be only one type of hemocyte in the systemic circulation of the adult intermolt animal, the so-called granulocyte or amebocyte. This cell is an oval, plate-shaped structure, 15-20 μm in its longest dimension. The cell contains numerous dense granules classed into two major types: Large (L) and Small (S) granules. The L-granules are larger (up to 1.5 μm in diameter) and less electron dense than the S-granules (0.6 μm in diameter). The L-granules contain at least 20 protein components including, 4 clotting factors and 1 antimicrobial factor (anti-LPS factor), whereas the S-granules exclusively contain the other antimicrobial substance, in addition to 6 major protein components. After treatment with bacterial endotoxin, the L-granules are released more rapidly than the S-granules, although almost all granules are finally exocytosed (Toh et al., 1991). This exocytosis is associated with clot formation, the process being complete within 90 sec. The clot is softer than mammalian fibrin clot and contains coagulin gel generated from its precursor, named coagulogen. This clottable protein, coagulogen, has a functional similarity with vertebrate fibrinogen and is known to play a central role in the limulus clotting system (Solum, 1970, 1973; Nakamura et al., 1976). This protein has been purified and its primary structure was recently elucidated (Tai et al., 1977; Miyata et al., 1984a).
LIMULUS AMEBOCYTE LYSATE (LAL)
Limulus Amebocyte Lysate (LAL) is a lyophilized preparation made from the amebocytes of the horseshoe crab Limulus polyphemus. The LAL will clot to form a gel in the presence of endotoxin from gram-negative microorganisms. Since the LAL test method is far more rapid, convenient, economical and sensitive than the USP rabbit pyrogen test method, it would be valuable for the determination of bacterial pyrogens in clinical diagnosis, pharmaceutical preparations, public health and environmental monitoring (Vanhaecke et al., 2008). The Food and Drug Administration (FDA) has proposed guidelines to standardize the sensitivity of LAL by controlling its production processes (Food and Drug Administration, 1973). The LAL has been sold with labeling against its use as a replacement for the official rabbit pyrogen test or as a diagnostic tool for determining the presence of endotoxemia in man. The Center for Devices and Radiological Health (CDRH) has adopted the USP Endotoxin Reference Standard and revised the limit expressions from ng mL-1 to EU mL-1. The new limit for medical. devices is 0.5 EU mL-1 except for devices in contact with cerebrospinal fluid for which the limit is 0.06 EU rnL-1. The Center for Drug Evaluation and Research (CDER) has added a listing of the maximum doses per kg per hour and the corresponding endotoxin limits for most of the aqueous injectable drugs and biologics currently on the market.
Limulus test: Amebocyte lysate is widely employed as the basis for a simple and sensitive in vitro assay (the Limulus test) for bacterial endotoxins (Levin and Bang, 1964a, b). At least three stages are involved in coagulation of the lysate (Nakamura and Iwanaga, 1978). The initial stage is activation of proclotting enzyme by direct action of endotoxin or by another factor(s) mediating the effect of endotoxin, resulting in production of an activated enzyme, designated clotting enzyme. The next step is limited proteolysis of the clottable protein, designated coagulogen, by the enzyme. The last phase is polymerization of proteolyzed coagulogen, designated coagulin, into a gel. Hemolymph coagulation cascade system in Tachypleus tridentatus is shown in Fig. 1.
Coagulogen: Coagulogen is a major protein in the hemolymph of many invertebrates and its role as a substrate in clot formation has been studied extensively (Nakamura et al., 1976; Tai et al., 1977; Moseson et al., 1979; Takagi et al., 1984; Cheng et al., 1986). The complete amino acid sequences of coagulogen have been reported for four species of horseshoe crabs (Miyata et al., 1984a, b; Srimal et al., 1985).
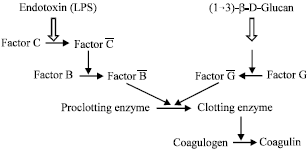 | |
| Fig. 1: | Hemolymph coagulation cascade system in Tachypleus tridentatus |
Previous studies have shown the presence of transglutaminase- catalyzed covalent cross-linking reaction of polypeptide chains during clot formation. However, there is no substantial evidence to demonstrate that the coagulin generated following cleavage of coagulogen is a substrate for such an enzyme. In contrast to the fibrin clot produced in the mammalian system, the coagulin clot can be dissociated either by simple mechanical disruption or dilution (Roth et al., 1989). Agglutination activities have also been observed in the hemolymph of other species of horseshoe crabs (Marehalonis and Edelman, 1968; Pistole, 1976; Shishikura and Sekiguchi, 1983). These lectin-like agglutination activities in invertebrates bear some similarities to the vertebrate immune system and could assume a crucial role in the recognition of foreign substances, resulting in their ultimate removal from the circulation (Vasta, 1990). It has also been observed that the ability of the amebocyte lysate to promote cell agglutination increases with the activation of the clotting enzymes (Fortes-Dias et al., 1993). Figure 2 shows how the horseshoe crab hemocyte functlons against invading microorganisms.
On incubation of horseshoe crab (Tachypleus tridentatus) coagulogen with an endotoxin-activated clotting enzyme, a peptide, named peptide C, is released; the resulting gel protein consists of two chains, named A and B.
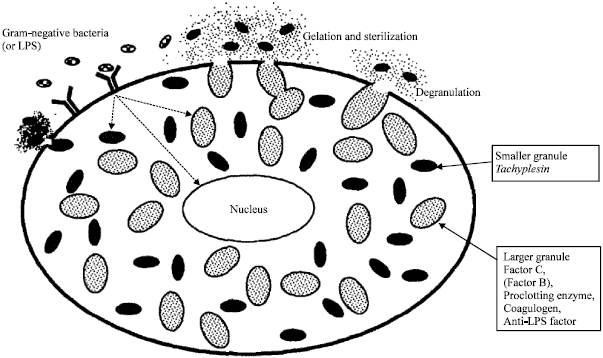 | |
| Fig. 2: | The figure Illustrates how the horseshoe crab hemocyte functlons against invading microorganisms. Bacterial endotoxin (lipopolysaccharide (LPS)) first contacts with an LPS receptor/LPSbinding protein, which is probably located in the plasma membrane of the cell. Through this contact, some cellular reactions induce the fusion of granules and the plasma membrane, resulting in dispersal of the components of the large granules into the plasma. This exocytosis activates the intracellular clotting system and the clot generated during activation immobilizes the invaders. The peptide tachyplesins and anti-LPS factor released from small and large granules, respectively, act as the bactericidal substances |
| Table 1: | Components closely associated with the biological defense system in limulus (Tachypleus tridentatus). Source: Iwanaga (1993) |
 | |
The complete amino acid sequences of peptide C and A-chain have been established (Nakamura et al., 1976a, b).
To establish the molecular events in such a reaction system, some researchers have reported on the gelation mechanism of coagulogen to coagulin using a purified active clotting enzyme on the complete amino acid sequence of coagulogen and on the development of a new method for determination of endotoxin, using chromogenic or fluorogenic peptide substrate of the clotting enzyme (Nakamura et al., 1976a, b, 1977; Takagi et al., 1979; Iwanaga et al., 1978). During these investigations, they have found out that proclotting enzyme is not an endotoxinsensitive protein but that another unknown factor (tentatively named factor B) sensitive to endotoxin is involved in the reaction sequence which mediates the activation of proclotting enzyme
PROCLOTTING AND CLOTTING ENZYMES
A clotting enzyme that catalyzes the transformation of coagulogen to coagulin exists as its zymogen form in the hemocytes (Muta et al., 1990). This proclotting enzyme is activated by both factor B and factor G (Fig. 1) Upon activation by factor B, it is converted to a two chain active form, clotting enzyme, composed of a Light (L) chain and a heavy (H) chain, with molecular masses of 25 and 31 kD, respectively. The clotting enzyme cleaves two, bonds in coagulogen to yield a coagulin gel. Thus, proclotting enzyme/clotting enzyme seem to be an invertebrate prothrombin/a-thrombin counterpart, since both catalyze a final step for gelation.
Factor B: The zymogen factor B is activated by factor c, which then, in turn, activates proclotting enzyme (Fig. 1). Factor B is a single chain glycoprotein with a molecular mass of 64 kD (Table 1). Upon activation of the zymogen factor B by active factor C, it is converted into active factor s, which is composed of an H chain (32 kD) and an L chain (25 kD), with the release of an activation peptide (Nakamura et al., 1986).
Factor C: As shown in Fig. 1, factor C is an initial activator of the clotting cascade tiggered by LPS (Tokunaga et al., 1987). The zymogen factor C is a glycoprotein with a calculated molecular mass of 123 kD. It consists of an H chain (80 kD) and an L chain (43 kD). Factor C is converted autocatalytically to an activated form, designated factor C (123 kD), in the presence of LPS. Upon activation, a single cleavage of the Phe-Ile bond in the L chain occurs, resulting in the accumulation of two new fragments, a B chain (34 kD) and an A chain (8.5 kD). Immunohistochemical observation with electron microscopy utilizing anti-factor antibody demonstrates that factor C is localized in the L granules in the cell (Toh et al., 1991).
INTERMEDIATION OF TACHYPLESIN
Tachyplesin I is an antimicrobial peptide isolated from the acid extracts of the membranes of Japanese horseshoe crab (Tachypleus tridentatus) hemocytes (Nakamura et al., 1988). Its isopeptides, tachyplesin II and polyphemusins I and II, have also been found in the hemocytes of the Japanese horseshoe crab (T. tridentatus) and the American horseshoe crab (Limulus polyphemus) (Miyata et al., 1989). Tachyplesins and polyphemusins were shown to inhibit the growth of both Gram-negative and positive bacteria at low concentrations and to form a complex with bacterial lipopolysaccharide (LPS), causing precipitation. It was reported that tachyplesin I binds to DNA and that this binding might be related to its biological action. Furthermore, these peptides were shown to have antiviral activity against vesicular stomatitis virus, influenza A virus and human immunodeficiency virus (HIV)-I (Yonezawa et al., 1992; Murakami et al., 1991; Morimoto et al., 1991; Masuda et al., 1992). Kawano et al. (1990) determined the secondary structure of tachyplesin I by nuclear magnetic resonance (NMR). This peptide was found to take an anti-parallel β-sheet structure (residues 3-8 and 11-16) connected by a β-turn (residues 8-11). Tachyplesin I was found to act as an inhibitor of LPS-mediated activation of factor C, which is the initiation factor in the limulus clotting cascade (Nakamura et al., 1988).
CONCLUSION
From all the results described above, we speculate that the β-glucan-mediated pathway may also lead to activation of this system. The clot generated during the activation encapsulates and immobilizes the Gram-negative bacteria and the peptide tachyplesin and anti-LPS factor released from S and L granules, respectively, act as the bactericidal substances. Therefore, this intracellular clotting system may have a crucial role in host defense against invading microorganisms. The precise gene responsible for the production of anti LPS in horseshoe crab need to be sequenced for the production of anti endotoxin compound in in vitro condition.
ACKNOWLEDGMENT
Authors express their sincere gratitude to the International Islamic University Malaysia (IIUM) and Institute of Oceanography and Maritime Studies (INOCEM) for providing infrastructure facilities.
REFERENCES
- Armstrong, P.B. and F.R. Rickles, 1982. Endotoxin-induced degranulation of the Limulus amebocyte. Exp. Cell Res., 140: 15-24.
Direct Link - Cheng, S.M., A. Suzuki, G. Zon and T.Y. Liu, 1986. Characterization of a complementary deoxyribonucleic acid for the coagulogen of Limulus polyphemus. Biochim. Biophys. Acta, 868: 1-8.
PubMed - Fortes-Dias, C.L., C.A. Minetti, Y. Lin and T.Y. Liu, 1993. Agglutination activity of Limulus polyphemus coagulogen following limited proteolysis. Comp. Biochem. Physiol. B, 105: 79-85.
Direct Link - Dumont, J.N., E. Anderson and G. Winner, 1966. Some cytologic characteristics of the hemocytes of Limulus during clotting. J. Morphol., 119: 181-207.
Direct Link - Hort, E.C. and W.J. Penfold, 1912. The relation of salvarsan fever to other forms of injection fever. Proc. R. Soc. Med., 5: 131-139.
Direct Link - Iwanaga, S., T. Morita, T. Harada, S. Nakamura and M. Niwa et al., 1978. Chromogenic substrates for horseshoe crab clotting enzyme: Its application for the assay of bacterial endotoxins. Haemostasis, 7: 183-188.
Direct Link - Kawano, K., T. Yoneya, T. Miyata, K. Yoshikawa, F. Tokunaga, Y. Terada and S. Iwanaga, 1990. Antimicrobial peptide, tachyplesin I, isolated from hemocytes of the horseshoe crab (Tachypleus tridentatus): NMR determination of the β-sheet structure. J. Biol. Chem., 265: 15365-15367.
Direct Link - Levin, J. and F.B. Bang, 1964. The role of endotoxin in the extracellular coagulation of limulus blood. Bull. Johns Hopkins Hosp., 115: 265-274.
Direct Link - Levin, J. and F.B. Bang, 1964. A description of cellular coagulation in the limulus. Bull. Johns Hopkins Hosp., 115: 337-345.
Direct Link - Marehalonis, J.J. and G.M. Edelman, 1968. Isolation and characterization of a hemagglutinin from Limulus polyphemus. J. Mol. Biol., 32: 453-465.
CrossRef - Masuda, M., H. Nakashima, T. Ueda, H. Naba and R. Ikoma et al., 1992. A novel anti-HIV synthetic peptide, T-22 ([TYR5,12, Lys7]-polyphemusin II). Biochem. Biophys. Res. Commun., 189: 845-850.
Direct Link - Miyata, T., M. Hiranaga, M. Umezu and S. Iwanaga, 1984. Amino acid sequence of the coagulogen from Limulus polyphemus hemocytes. J. Biol. Chem., 259: 8924-8933.
Direct Link - Miyata, T., F. Tokunaga, T. Yoneya, K. Yoshikawa and S. Iwanaga et al., 1989. Antimicrobial peptides, isolated from horseshoe crab hemocytes, tachyplesin II and polyphemusins I and II: Chemical structures and biological activity. J. Biochem., 106: 663-668.
Direct Link - Morimoto, M., H. Mori, T. Otake, N. Ueda and N. Kunita et al., 1991. Inhibitory effect of tachyplesin i on the proliferation of human immunodeficiency virus in vitro. Chemotherapy, 37: 206-211.
Direct Link - Murakami, T., M. Niwa, F. Tokunaga, T. Miyata and S. Iwanaga, 1991. Direct virus inactivation of tachyplesin i and its isopeptides from horseshoe crab hemocytes. Chemotherapy, 37: 327-334.
Direct Link - Murer, E.H., J. Levin and R. Holm, 1975. Isolation and studies of the granules of the amebocytes of Limulus polyphemus, the horseshoe crab. J. Cell Physiol., 86: 533-542.
Direct Link - Muta, T., R. Hashimoto, T. Miyata, H. Nishimura, Y. Toh and S. Iwanaga, 1990. Proclotting enzyme from horseshoe crab hemocytes: cDNA cloning, disulfide locations and subcellular localization. J. Biol. Chem., 265: 22426-22433.
Direct Link - Nakamura, T., T. Horiuchi, T. Morita and S. Iwanaga, 1986. Purification and properties of intracellular clotting factor, factor b, from horseshoe crab (Tuchypleus tridentatus) hemocytes. J. Biocbem., 99: 847-857.
Direct Link - Nakamura, S. and S. Iwanaga, 1978. Haemostasis and coagulation in horseshoe crab-on the mechanism of gel filtration of the amebocytes lysate induced by endotoxins (authors transl). Protein Nucleic Acid Enzyme, 23: 277-290.
Direct Link - Nakamura, S., T. Morita, S. Iwanaga, M. Niwa and K. Takahashi, 1977. A sensitive substrate for the clotting enzyme in horseshoe crab hemocytes. J. Biochem., 81: 1567-1569.
Direct Link - Nakamura, S., T. Takagi, S. Iwanaga, M. Niwa and K. Takahashi, 1976. . Amino acid sequence studies on the fragments produced from horseshoe crab coagulogen during gel formation: Homologies with primate fibrinopeptide B. Biochem. Biophys. Res. Commun., 72: 902-908.
Direct Link - Nakamura, S., S. Iwanaga, T. Harada and M. Niwa, 1976. A dottable protein (coagulogen) from amebocyte lysate of Japanese horseshoe crab Tachypleus tridentatus. J. Biochem., 80: 1011-1021.
Direct Link - Nakamura, T., H. Furunaka, T. Miyata, F. Tokunaga and T. Muta et al., 1988. Tachyplesin, a class of antimicrobial peptide from the hemocytes of the horseshoe crab (Tachypleus tridentatus): Isolation and chemical structure. J. Biol. Chem., 263: 16709-16713.
Direct Link - Ornberg, R.L. and T.S. Reese, 1981. Beginning of exocytosis captured by rapid-freezing of Limulus amebocytes. J. Cell Biol., 90: 40-54.
Direct Link - Pistole, T.G., 1976. Naturally occurring bacterial agglutinin in the serum of the horseshoe crab Limulus polyphemus. J. Invertebr. Pathol., 28: 153-154.
Direct Link - Prior, R.B. and V.A. Spagna, 1979. Adaptation of a microdilution proceduer to the limulus lysate assay for endotoxin. J. Clin. Microbiol., 10: 394-395.
Direct Link - Roth, R.I., J.C. Chert and J. Levin, 1989. Stability of gels formed following coagulation of Limulus amebocyte iysate: Lack of covalent crosslinking of coagulin. Thromb. Res., 55: 25-36.
Direct Link - Shishikura, F. and K. Sekiguchi, 1983. Agglutinins in the horseshoe crab hemolymph: Purification of a potent agglutinin of horse erythrocytes from the hemolymph of Tachypleus tridentatus, the Japanese horseshoe crab. J. Biochem., 93: 1539-1546.
Direct Link - Solum, N.O., 1970. Some characteristics of the clottable protein of Limuluspolyphemus blood cells. Thromb. Diath. Haemorrh., 23: 170-181.
Direct Link - Solum, N.O., 1973. The coagulogen of Limuluspolyphemus hemocytes. A comparison of the clotted and non-clotted forms of the molecule. Thromb. Res., 2: 55-69.
CrossRef - Miyata, T., K. Usui and S. Iwanaga, 1984. The amino acid sequence of coagulogen isolated from southeast Asian horsechoe crab Tachypleus gigas. J. Biochem., 95: 1793-1801.
Direct Link - Food and Drug Administration, 1973. Limulus amebocyte lysate, additional standards. Code of federal regulations 21 CFR part 273. Fed. Regist., 38: 26130-26132.
Direct Link - Tai, J.Y., R.C. Seid, R.D. Huhn and T.Y. Liu, 1977. Studies on Limulus amebocyte lysate. II. Purification of the coagulogen and the mechanism of clotting. J. Biol. Chem., 252: 4773-4776.
Direct Link - Takagi, T., Y. Hokama, T. Myiata, T. Morita and S. Iwanaga, 1984. Amino acid sequence of Japanese horseshoe crab (Tachypleus tridentatus) coagulogen B. chain: Completion of the coagulogen sequence. J. Biochem., 95: 1445-1457.
Direct Link - Toh, Y., A. Mizutani, F. Tokunaga, T. Muta and S. Iwanaga, 1991. Structure of hemocytes of the Japanese horseshoe crab Tachypleus tridentatus: Fine structure, morphological changes during coagulation and localization of clotting factors and antimicrobial substances. Cell Tissue Res., 266: 137-147.
Direct Link - Tokunaga, F., T. Miyata, T. Nakamura, T. Morita, K. Kuma, T. Miyata and S. Iwanga, 1987. Lipopolysaccharide-sensitive serineprotease zymogen (Factor C) of horseshoe crab hemocytes: Identification and alignment of proteolytic fragment produced during the activation show that it is a novel type of serine protease. Eur. J. Biochem., 167: 405-416.
- Yonezawa, A., J. Kuwahara, N. Fujii and Y. Sugiura, 1992. Binding of tachyplesin I to DNA revealed by footprinting analysis: Significant contribution of secondary structure to DNA binding and implication for biological action. Biochemistry, 31: 2998-3004.
Direct Link - Srimal, S., T. Miyata, S. Kawabata, T. Miyata and S. Iwanaga, 1985. The complete amino acid sequence of coagulogen isolated from southeast Asian horseshoe crab, Carcinoscorpius rotundicauda. J. Biochem., 98: 305-318.
Direct Link
Amuzat Aliyu olalekan Reply
It is a useful and beneficial article to me