
N. Ganikhodjaev
Institute of Mathematics and Information Technology, 100125, Tashkent, Uzbekistan
J.I. Daoud
Faculty of Engineering, IIUM, 50728, Kuala Lumpur, Malaysia
M. Usmanova
Faculty of Science, IIUM, 25200, Kuantan, Malaysia
Journal of Applied Sciences
Year: 2010 | Volume: 10 | Issue: 16 | Page No.: 1748-1754







ABSTRACT
We consider linear and nonlinear stochastic models for transmission of blood types and Rhesus factor from parents to their offspring and investigate long run behavior of these models. In this study we will consider an application of the theory of Markov chains and the theory of nonlinear transformations in medicine. It is well known that the gene which determines blood group in humans has three different alleles, A, B, C and that there are four groups of blood, A, B, AB and O. The aim is to investigate the transmission of blood groups from parents to their offspring. For simplicity, we will consider only positive Rhesus factors, since the portion of the population with negative Rhesus is around 1%. It is well known that the blood groups of parents do not determine unambiguously their offspring’s blood group. To describe this transmission, we have rather extensive statistics for blood groups of parents and their offspring. In connection with these statistics, we construct the following two Markov chains. The first Markov chain describes the transmission from a father to his sons; the second Markov chain describes the transmission from a mother to her daughters. Lastly we construct and study a quadratic stochastic operators that describe the transmission of blood groups from parents to their offspring.
PDF Abstract XML References Citation
Received: February 23, 2010;
Accepted: May 23, 2010;
Published: June 26, 2010
How to cite this article
N. Ganikhodjaev, J.I. Daoud and M. Usmanova, 2010. Linear and Nonlinear Models of Heredity for Blood Groups and Rhesus Factor. Journal of Applied Sciences, 10: 1748-1754.
DOI: 10.3923/jas.2010.1748.1754
URL: https://scialert.net/abstract/?doi=jas.2010.1748.1754
DOI: 10.3923/jas.2010.1748.1754
URL: https://scialert.net/abstract/?doi=jas.2010.1748.1754
INTRODUCTION
Blood groups are distinguished by the blood’s antigenic properties. These properties are determined by the substances found on the surface of the red blood cells. There are approximately 200 blood group substances identified and categorized into 19 distinct systems. The most common system is the ABO system. The human ABO blood group was discovered by Karl Landsteiner in 1900 (Landsteiner, 1900) and its mode of inheritance through multiple alleles at a single generic locus was established by Felix Bernstein a quarter century later (Bernstein, 1925). The ABO blood group antigens appear to have been important throughout our evolution because the frequencies of different ABO blood types vary among different populations, suggesting that a particular blood type conferred a selection advantage.
There are three alleles or versions of the blood type gene: A, B and C. Since, humans are diploid organisms (meaning we carry a double set of chromosomes-one from each parent), blood types are determined by two alleles (Table 1). There are six possible combinations of such alleles: AA, BB, OO, AB, OA and OB. In genetic terms, these combinations are called genotypes and they describe the genes that an offspring inherited from his parents.
| Table 1: | The ABO blood system |
| Table 2: | ABO phenotype in the offspring |
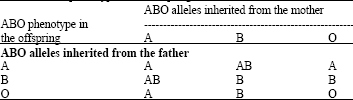 | |
The expression of the O allele is recessive to that of A and B, which are said to be co-dominant. Thus, the genotypes AO and AA express blood type A, BO and BB express blood type B, AB expresses blood type AB and OO expresses blood type O.
The ABO blood group antigens are encoded by one genetic locus, the ABO locus, which has three alternative (allelic) forms- A, B and O. It is well known that the heredity of blood groups behaves according to the Mendelian rules. A child receives one of the three alleles from each parent, giving rise to six possible genotypes and four possible blood types (phenotypes) (Table 2).
Offspring blood type is established by specific genes inherited from his parents. He receives one gene from his mother and one from his father; these two combine to establish his blood type.
| Table 3: | Offspring’s blood type |
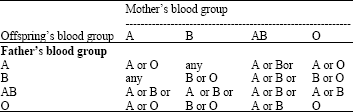 | |
| Table 4: | Offspring’s Rhesus genes |
 | |
As mentioned above, the heredity of blood groups behaves according to the Mendelian rules (Table 3).
According to Table 3, a group AB person cannot be the parent of a group O child, nor can a group O person be the parent of a group AB child. However, transmission of blood group AB in a family as if by a single chromosome or allele, instead of by two separate chromosomes or genes were reported and this led to the discovery of a very rare blood group called cis-AB. The ABO groups are inherited through multiple alleles at one locus, as seen by Bernstein nearly 80 yeas ago (Bernstein, 1925). Now concerning the inheritance of the ABO groups, the precise mechanism may change, although the triple allele theory of Bernstein is adequate for all practical purposes. The ABO blood group locus includes some sites where mutation can occur and between which, as an extreme rarity, crossing over can happen. The Rhesus system is the second most significant blood group system in human blood transfusion. Individuals either have, or do not have, the Rhesus factor (or Rh(D) antigen) on the surface of their red blood cells. This is usually indicated by Rh+ (does have the Rh(D) antigen) or Rh¯ (does not have the antigen) suffix to the ABO blood type.
A child inherits two Rhesus genes, one from each parent, where gene D corresponds to a positive Rhesus factor and gene d corresponds to a negative Rhesus factor (Table 4).
Offspring are Rhesus negative if they have inherited a d gene from each parent (d, d) and offspring are Rhesus positive if they inherited a D gene from both parents. If offspring have inherited a Rhesus positive gene D and a Rhesus negative gene d, they are most likely to be Rhesus positive as the D gene is more dominant as compared to the d gene. Hence, it is possible to have a Rhesus negative child and a Rhesus positive father. According Table 5, a husband and wife with negative Rhesus factor cannot be the parent of a positive Rhesus factor child. However, collected data shows the existence of families that contradict Mendel laws.
| Table 5: | Offspring’s Rhesus factor |
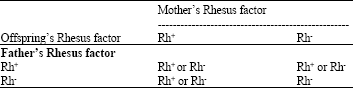 | |
Thus, blood groups and Rhesus factor of parents do not determine unambiguously their offspring’s blood group and Rhesus factor (Table 3, 5). The transmission of blood group and its Rhesus factor from parents to their offspring are random and events that contradict Mendel laws increase this randomness. To study these transmissions we consider two types of stochastic modeling:
| • | Markov chains |
| • | Quadratic stochastic operators |
The theory of Markov chains is well known and describes a linear model.
QUADRATIC STOCHASTIC OPERATORS
Quadratic stochastic operators were first introduced by Bernstein (1924). Such operators frequently arise in models of mathematical genetics (Jenks, 1969; Kesten, 1970; Lyubich, 1971; Reed, 1997; Ganikhodjaev, 1999). Consider a biological population, such as a community of organisms that is closed with respect to reproduction. Assume that each individual in this population belongs to precisely one species i = 1,...,m. Below, we consider the scale of blood type with m = 4 and Rhesus factor with m = 2. The scale of species is such that the species of the parents i and j unambiguously determines the probability of every species k .for the first generation of direct descendants. Denote this probability, that is to be called the heredity coefficient, by pij,k. It is obvious that pij,k≥0 and
Assume that the population is so large that frequency fluctuations can be neglected. Then, the state of the population can be described by the m-tuple (x1,x2,...,xm) of species probabilities, where, xk is the fraction of the species k in the total population. In the case of panmixia (random interbreeding) the parent pairs i and j arise for a fixed state x =(x1,x2,...,xm) with probability xixj. Hence, the total probability of the species k in the first generation of direct descendants is defined by:
| (1) |
Let,
be the (m-1) dimensional canonical simplex in Rm. The transformation V : Sm-1⇒Sm-1 is called a quadratic stochastic operator if:
| (2) |
where, a: ![]() , b:
, b:![]() and c:
and c:
for arbitrary i,j, k = 1,...,m.
Note that the condition (b) is not overloaded, since otherwise one can determine new heredity coefficients:
while preserving the operator V:
| (3) |
The set of transformations Eq. 2 or 3, which describes a model of heredity is called a quadratic stochastic operator. This model of heredity is uniquely determined by heredity coefficients:
Assume {x(k): k = 0,1,2,...} is the trajectory of the initial point x∈Sm-1, where, x(k+1) = V(x(k)) for all k = 0,1,2,... with x(0) = x.
To investigate limit behavior of trajectories and fixed points of the quadratic stochastic operator V, (2) plays an important role in many applied problems.
A fixed point of a quadratic stochastic operator V is a point x =a where V(a) = a.
A quadratic stochastic operator V is called regular if, for each initial point x∈Sm-1, the limit:
exists. Note that the limit point is a fixed point of a quadratic stochastic operator V.
Thus, the fixed points of a quadratic stochastic operator describe limit or long run behavior the trajectories of any initial points.
Let, us consider the linearization of the Eq. 3 near the fixed point a. Assume that a is an internal fixed point. We write y = x-a where, x∈Sm-1 implies:
Then, the evolution equation for y becomes:
where, J is a Jacobian that is a matrix whose entries are:
and the approximation is valid as long as y remains small.
Proposition 1: A Jacobian J, of quadratic stochastic operator V at a fixed point is a double- column stochastic matrix.
Proof: Direct calculus proves this statement.
Corollary: If J is a Jacobian of quadratic stochastic operator V at a fixed point, then:
| • | λ1 = 2 is an eigenvalue of J. The entries of the corresponding eigenvector b1 are all nonnegative. We can normalize b1 so that the sum of its entries is 1 |
| • | The sum of the entries of y(n) = Jny(0) is the same as the sum of entries of y(0), i.e., zero |
| • | If J is a regular, then the only neutrally stable mode is b1 |
All statements are simple reformulations corresponding to properties of stochastic matrices. J. The entries of the corresponding eigenvector b1 are all.
RHESUS FACTOR TRANSMISSION
Firstly, we consider a linear model of transmission, namely, the Markov chain. A Markov chain describes transmission, on some scale, from one of the parents to their offspring of the same gender. Note that only for such Markov chains, can one study their limiting distribution or long run behavior. Below, we consider two Markov chains: first, a Markov chain that describes a transmission of Rhesus factor from fathers to their sons and second, one that describes transmission of Rhesus factor from mothers to their daughters. For the collected data, let Ns(Fx) be the number of sons of fathers Fx (fathers with Rhesus factor X) and NYS (Fx) be the number of sons with Rhesus factor Y of fathers Fx where, X, Y∈{+,-}. Then:
To describe the transmission of Rhesus factor from fathers to their sons, we need to find the probability Pxy that a son will inherit a Rhesus factor Y from a from a father with Rhesus factor X, where, X,Y∈{+,-}. Thus, we denote:
Then, according to the collected data the transmission probability matrix of the first Markov chain has the form:
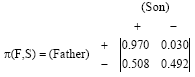 | (4) |
and the transmission probability matrix of the second Markov chain, which describes the transmission of Rhesus factor from a mother to her daughters, has the form:
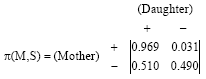 | (5) |
Both Markov chains are regular with limiting distribution:
for the first Markov chain and
for the second one.
Now we consider nonlinear model of transmission, namely quadratic stochastic operators. Let a set of species be a set of Rhesus factor {1, 2}, where, 1 denotes positive Rhesus and 2 denotes negative Rhesus. To describe the transmission of Rhesus factor from parents to their offspring, we need to find the probability PXY,Z that a child receives Rhesus factor Z from a father with Rhesus factor X and a mother with Rhesus factor Y, where, X, Y, Z∈{1,2}. Let, N(Fx,MY) be the number of offspring of fathers Fx and mothers My (fathers with Rhesus factor X and mothers with Rhesus factor Y) and NZ(Fx,My) be the number of offspring with Rhesus factor Z of fathers Fx and mothers My. Then the transmission probability PXY,Z is defined as:
| (6) |
Let,
be the heredity coefficients for i, j, k∈ {1,2}.
For the data collected and according to Eq. 6, we have the following:
| (7) |
where,
and the corresponding quadratic stochastic operator has the form:
| (8) |
where, x1 is the fraction of the population with positive Rhesus factor and x2 is the fraction of the population with negative Rhesus factor.
The Eq. 8 has single fixed point:
| (9) |
The Jacobian of the quadratic stochastic operator Eq. 8 at the fixed point has following form:
with eigenvalues λ1 = 0.685 and λ2 = 2. Then, the fixed point Eq. 9 is stable and any trajectory of the quadratic stochastic operator Eq. 8 converges to the fixed point Eq. 9. Thus, the quadratic stochastic operator Eq. 8 is a regular.
BLOOD TYPE TRANSMISSION
Now, let us consider transmission of blood type. As above, first we study the transmission of blood type from a father to his sons.
Let, NS(Fx) be the number of sons of fathers Fx (fathers with blood group X) and NYS(Fx) be the number of sons with blood group Y of fathers Fx where, X, Y∈{A, B, AB, 0}.Then ![]() .
.
The state of the population can be described by the quadruple (xA, xB, xAB, xO) where, xA is the fraction of the population with blood group A. Similarly, fractions xB, xAB, xO, represent blood groups B, AB and O, respectively.
The transmission probabilities matrix:
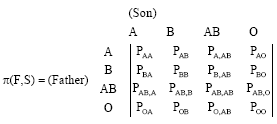 |
is defined as:
| (10) |
For the collected data of 3965 sons, using Eq. 10 we obtained the following transmission probability matrix:
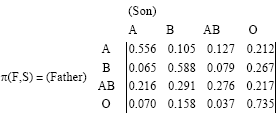 | (11) |
Remark 1: According to Table 3, we should have PAB,O = 0 and PO, AB = 0. However, one can see that PAB,O = 0.195 and PO, AB = 0.040. The Markov chain with transmission probability matrix Eq. 11 has the following limiting distribution:
| (12) |
Similarly, in the case of transmission of blood types from a mother to her daughters, for the collected data of 5637 daughters, we have the following transmission probability matrix:
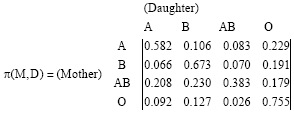 | (13) |
Remark 2: Note that according to Table 3, we should have PAB,O = 0 and PO,AB = 0. However, PAB,O = 0173 and PO,AB = 0.024. The Markov chain with transmission probability matrix Eq. 13 has the following limiting distribution:
| (14) |
Now, consider the transmission of blood groups from parents to their offspring. To describe this transmission we will apply quadratic stochastic operators.
Let a set of species be a set of blood types {A,B,AB,O}. To study the transmission of blood group from parents to their offspring we need to find a probability PXY,Z that from a father with blood group X and a mother with blood group Z, their child receives the blood group Z, where X, Y, Z∈ {A,B,AB,O}. Let, N(FX,MY) be the number of offspring of fathers FX (with blood group X) and mothers MY (with blood group Y) and NZ(FX,MY) be the number of offspring with blood group Z of fathers FX and mothers MY.
Then, the transmission probability PXY,Z is defined as:
| (15) |
For brevity let’s replace symbols {A,B,AB,O} by {1,2,3,4}, respectively, so that blood group A corresponds to 1, blood group B corresponds to 2, blood group AB corresponds to 3 and blood group O corresponds to 4.
Then, the coefficients of heredity PXY,Z for i,j,k∈{1,2,3,4} are defined in the following way:
where, X corresponds i, Y corresponds to j and Z corresponds to k.
Generally, the coefficients pij,k do not satisfy the condition pij,k = pji,k.
Let,
For the collected data we obtained:
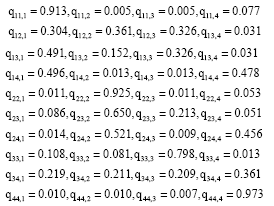 |
Remark 3: One can see that, the values of some qij,k contradict those in Table 3. Clear examples of this can be seen in q11,2, q11,3 q44, 3 and so on.
The corresponding quadratic stochastic operator has the following form:
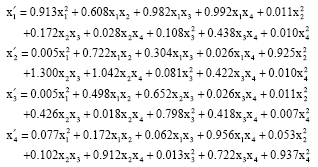 | (16) |
This quadratic stochastic operator has a single fixed point in simplex S3.
| (17) |
The Jacobian of the quadratic stochastic operator Eq. 16 at the fixed point has the following form:
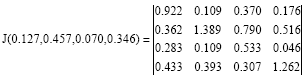 |
with eigenvalues![]() . Then the fixed point Eq. 17 is stable and any trajectory of quadratic stochastic operator Eq. 16 converges to fixed point Eq. 17. Thus, the quadratic stochastic operator Eq. 16 is a regular.
. Then the fixed point Eq. 17 is stable and any trajectory of quadratic stochastic operator Eq. 16 converges to fixed point Eq. 17. Thus, the quadratic stochastic operator Eq. 16 is a regular.
DISCUSSION
Rhesus factor: For the linear models we have, we see that frequencies of Rhesus factor among men and women are the same and there is no significant association between sex and Rhesus factor. Secondly, the probability that a son or daughter of a father or mother (respectively) with negative Rhesus factor will inherit positive Rhesus factor is equal to 0.5. For the nonlinear model from Eq. 7 one can see that if parents have opposite Rhesus factors then two thirds (2/3) of their offspring inherit positive Rhesus factor and one third (1/3) inherit negative Rhesus factor.
Now consider the received heredity coefficients qij,k: One can see that there are instances when the chart of Table 2 is not accurate, since, q22,1 = 0.092 means that a child of parents with negative Rhesus factor receives positive Rhesus factors with probability 0.092. In the case of a mutation, the Rhesus factor typing may not hold true in the question of parentage.
We can state that both models, linear and nonlinear, predict the same long run distribution of Rhesus factor.
Blood type transmission: In different parts of the world, the fraction of individuals with blood type A, B AB or O differs. The frequency with which blood types are observed is determined by the frequency with which the three alleles of the ABO gene are found in different parts of the world (allele frequency). Variation in the allele frequency at the ABO gene reflects the social tendency of populations to marry and reproduce within a national, regional, or ethnic group.
Using two different models, linear and nonlinear, we received the following limiting distributions:
| • | Transmission from fathers to their sons: |
| • | Transmission from mothers to their daughters: |
| |
| • | Transmission from parents to their offspring: |
| |
We state that the last distribution (c) more exactly predicts the distribution of blood type after a long period.
Now let us consider Remarks 1-3 (Seyfried et al., 1964) and Yamaguchi et al. (1965, 1966) described instances in which blood group O was inherited from one parent and both blood group A and blood group B from the other parent. This was referred to as cis-AB to discriminate this rare phenotype from ordinary trans-AB. Yoshida et al. (1980) reported two possible genetic mechanisms: unequal chromosomal crossing over and structural mutation in the blood group glycosyltransferase. In the latter instance, mutation in either the A or the B gene had produced a single abnormal enzyme with bifunctional activity.
In the case of a mutation, the blood typing may not hold true in the question of parentage. Occasionally, an O mother and an AB father may give birth to an AB child. The interpretation is cis-AB, i.e., both alleles on the same chromosome, or one allele with both specificities. Hummel et al. (1977) traced such through three generations. Inherited mosaicism in the ABO system consists of a situation in which, in an autosomal dominant pedigree pattern, family members show mosaicism of A cells and O cells, or B cells and O cells. A mixed field agglutination pattern results. This phenotype is probably caused by a weak allele rather than by a modifier gene. Bird et al. (1978) found that in a B-O mosaic family, affected persons had low levels of B-specific transferase.
Thus, in general, the rules of transmission of blood group or Rhesus factor are random and we cannot predict the blood group of the child. Using the models of heredity considered above, we can find a limiting distribution of blood group and Rhesus factor that have certain applied significance.
Finally, note that even if the data was collected in two regions of Malaysia, our results are probably valid within any national, regional, or ethnic group.
ACKNOWLEDGMENT
This study supported by Research Endowment Grant EDW B0801-59 of International Islamic University Malaysia.
REFERENCES
- Bernstein, F., 1925. Zusammenfassende betrachtungen uber die erblichen blutstrukturen des menschen. Mol. General Genet., 37: 237-370.
CrossRef - Bird, G.W., J. Wingham, W.M. Watkins, P. Greenwell and A.H. Cameron, 1978. Inherited mosaicism within the ABO blood group system. J. Immunogenet., 5: 215-219.
PubMed - Hummel, K., J. Badet, W. Bauermeister, K. Bender and G. Duffner et al., 1977. Inheritance of cis-AB in three generations (family Lam.). Vox Sang., 33: 290-298.
PubMed - Kesten, H., 1970. Quadratic transformations: A model for population growth. I. Adv. Appl. Prob., 2: 1-82.
Direct Link - Landsteiner, K., 1900. Zur Kenntnis der antifermentativen, lytischen und agglutinierenden Wirkungen des Blutserums und der Lymphe. Zentralblatt Bakt., 27: 357-362.
Direct Link - Lyubich, Y.I., 1971. Basic concepts and theorems of the evolution genetics of free populations. Russian Math. Surveys, 26: 51-116.
CrossRefDirect Link - Reed, M.L., 1997. Algebraic structure of genetic inheritance. Bull. Am. Math. Soc., 34: 107-130.
CrossRefDirect Link - Yamaguchi, H., Y. Okubo and F. Hazama, 1965. An A2B3 phenotype blood showing atypical mode of inheritance. Proc. Jap. Acad., 41: 316-320.
Direct Link - Yamaguchi, H., Y. Okubo and F. Hazama, 1966. Another Japanese A2B3 blood-group family with propositus having O-group father. Proc. Jap. Acad., 42: 517-520.
Direct Link - Yoshida, A., H. Yamaguchi and Y. Okubo, 1980. Genetic mechanism of cis-AB inheritance. II. Cases associated with structural mutation of blood group glycosyltransferase. Am. J. Hum. Genet., 32: 645-650.
Direct Link