
Jutti Levita
Faculty of Pharmacy, Universitas Padjadjaran,Jl. Raya Bandung-Sumedang Km. 21, Jatinangor, West Java, Indonesia
As `ari Nawawi
School of Pharmacy, Institut Teknologi Bandung,Jl. Ganesha 10 Bandung, West Java, Indonesia
Abdul Mutholib
Center for Radioisotope and Radiopharmaca, National Nuclear Agency, Serpong, Indonesia
Slamet Ibrahim
School of Pharmacy, Institut Teknologi Bandung,Jl. Ganesha 10 Bandung, West Java, Indonesia
Journal of Applied Sciences
Year: 2010 | Volume: 10 | Issue: 14 | Page No.: 1481-1484







ABSTRACT
The aim of this study was to identify and analyze the interaction of andrographolide with COX-2, followed by in vitro study of the effect of this compound on COX-2 expression in human fibroblast cells. The molecular modeling study was performed by docking andrographolide to COX-2 enzyme at the site where SC-558, a selective inhibitor of this enzyme, was co-crystallized and compared its interaction to the enzyme with SC-558’s. The inhibition of COX-2 expression was determined by measuring PGE2 production in human fibroblast cells stimulated with LPS with and without andrographolide preincubations. Andrographolide interacted with Arg513 and His90 in the cyclooxygenase site of COX-2 and inhibited PGE2 production in human fibroblast cells (IC50 = 4 μM). These data confirm that andrographolide’s anti-inflammatory activity occurs via inhibition of COX-2 expression.
PDF Abstract XML References Citation
Received: March 25, 2010;
Accepted: May 06, 2010;
Published: June 10, 2010
How to cite this article
Jutti Levita, As `ari Nawawi, Abdul Mutholib and Slamet Ibrahim, 2010. Andrographolide Inhibits COX-2 Expression in Human Fibroblast Cells Due to its Interaction with Arginine and Histidine in Cyclooxygenase Site. Journal of Applied Sciences, 10: 1481-1484.
DOI: 10.3923/jas.2010.1481.1484
URL: https://scialert.net/abstract/?doi=jas.2010.1481.1484
DOI: 10.3923/jas.2010.1481.1484
URL: https://scialert.net/abstract/?doi=jas.2010.1481.1484
INTRODUCTION
Andrographolide, an active component of Andrographis paniculata, is the major labdane diterpenoidal constituent in this plant, which is used extensively in the traditional medicine in Indonesia to treat inflammations. This compound has an α-alkylidene γ-butyrolactone, two olefin bonds at C8(17) and C12(13) and three hydroxyl groups at C3, C19 and C14 (Nanduri et al., 2004). These features make andrographolide able to form hydrogen bonds, both as donors and acceptors and undergo hydrophobic interactions via its methyl group and olefin bonds.
Andrographolide (Fig. 1) has been reported to have anti-inflammatory activity by suppressing inducible nitric oxide synthase expression in RAW 264.7 cells (Chiou et al., 2000), prevents oxygen radical production by human neutrophils (Shen et al., 2002), inhibits NF-kappaB activation (Xia et al., 2004) and reduced COX-2 expression induced by platelet activating factor and N-formyl-methionyl-leucyl-phenylalanine in HL60/neutrophils (Hidalgo et al., 2005). Direct antimicrobial activity of two ethanolic Andrographis paniculata extracts was observed for two human pathogens, Legionella pneumophila and Bordetella pertussis (Xu et al., 2006).
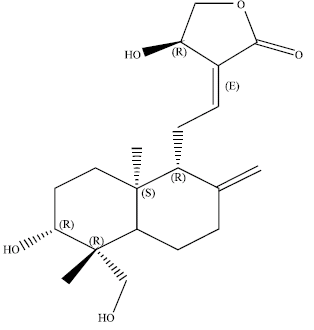 | |
| Fig. 1: | Andrographolide |
The two definitely known isoforms of COX, named COX-1 and COX-2, show distinct expressions patterns
and distinct biological activities. COX-1 is a constitutively expressed protein that is responsible for the physiological production of prostaglandins. COX-2 is rapidly up-regulated at inflammatory sites and appeared responsible for the formation of prostaglandin. In inflammatory processes COX-2 is over expressed. Non-steroidal anti-inflammatory drugs (NSAIDs) are COX inhibitors and prevent prostaglandin synthesis, thus exhibiting analgesic, antipyretic and anti-inflammatory actions. However, NSAIDs cause gastrointestinal adverse effects, mainly because of their inhibition of the constitutive isoform of COX. Since selective COX-2 inhibitors fail to inhibit constitutive COX-1 isoform, they have no gastrointestinal adverse effects. (Dilber et al., 2008). A new class of compounds has recently been developed, named SC-58125, that have a high degree of selectivity for the inducible form of COX-2 over the constitutive form of COX-1. This unique class of compounds exhibits a time dependent irreversible inhibition of COX-2, while reversibly inhibiting COX-1. The molecular basis of this selectivity was probed by site-directed mutagenesis at the active site of COX-2. One difference is observed at the cyclooxygenase active site, where the isoleucine at position 523 in COX-1 is a valine in COX-2 (Gierse et al., 1996).
In this study we sought to identify and analyze the interaction of andrographolide with COX-2 in terms of hydrogen bonds, followed by in vitro study of the effect of this compound on COX-2 expression in human fibroblast cells.
MATERIALS AND METHODS
Molecular modeling study (December 2008 to March 2009): The X-ray crystallographic 3D structures of COX-2 (PDB code: 1CX2) complexed with its selective inhibitor SC-558 crystallized by Kurumbail et al. (1996) was downloaded from online Protein Data Bank. 3D structure of andrographolide was built by using Molecular Operating Environment (MOE-2007.09.02). Energy minimization was carried out by using MMFF94x. Docking of andrographolide to COX-2 enzyme was carried out at the site where SC-558 was co-crystallized.
In vitro study (November 2009 to March 2010)
Chemicals and samples: Andrographolide 98% 500 mg CAS 5508-58-7 for R and D use was purchased from Aldrich. Prior to use the compound was dissolved in 2% dimethylsulfoxide (DMSO) in Phosphate Buffer Saline (PBS). All other chemicals were dissolved in phosphate buffer saline or MilliQ water (Sentra). High glucose Dulbecco’s Modified Eagle Medium (DMEM) which contained D-glucose, L-glutamine, sodium pyruvate for culture medium was purchased from Gibco, lipopolysaccharide (LPS) 10 mg derived from Escherichia coli 0127:B8 was purchased from Sigma-Aldrich.
Human fibroblast cell culture and differentiation: Human fibroblast cells were a kind gift from Dr. Endang Winiarti (Research Laboratory, Faculty of Dentistry, Universitas Indonesia, Jakarta, Indonesia). The cells were grown in high glucose Dulbecco’s Modified Eagle Medium (DMEM) which contained D-glucose, L-glutamine, sodium pyruvate (Gibco), supplemented with 10% heat-inactivated FBS (fetal bovine serum), penicillin (100 U mL–1), streptomycin (100 μg mL–1) and fungizone, at 37°C under 5% CO2. The cells were differentiated by incubating them in their culture medium for 48 h and were collected at the third day for further assay.
Cyclooxygenase assay by measuring PGE2 production: The inhibition of cyclooxygenase was determined based on procedure described by Young et al. (1996) with slight modifications. Briefly, fibroblast cells (5.2x104 cells) in high glucose DMEM supplemented with 10% heat-inactivated FBS, penicillin (100 U mL–1), streptomycin (100 μg mL–1) and fungizone, were placed in a 96-well microplate and were stimulated with LPS (10 μg mL–1) to produce prostaglandin (PGE2), a protein which production was catalyzed by cyclooxygenase enzyme. Various concentrations of andrographolide (2 to 200 μM) were added into the wells and the mixtures were incubated for 18 h at 37°C under 5% CO2. Acetyl salicylic acid (0.7 to 40 μM) was used as the standard. The production of PGE2 was measured by using microplate reader at 450 nm.
RESULTS AND DISCUSSION
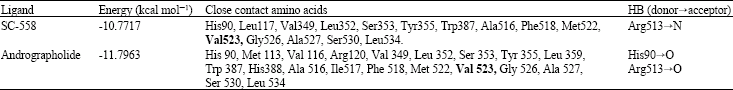
Molecular modeling study: The binding site of COX-2 where its selective inhibitor, SC-558, was bound (Fig. 2a) contained His90, Leu117, Val349, Leu352, Ser353, Tyr355, Trp387, Ala516, Phe518, Met522, Val523, Gly526, Ala527, Ser530, Leu534. This molecular modeling s tudy of COX-2 binding site which contained Valine at position 523 confirmed Gierse et al. (1996)‘s study.
Based on our study, SC-558 showed only one hydrogen bond with Arg513 (Fig. 2a). The hydrogen bond was formed between SC-558 amine and Arg513 at 3.15 (Table 1), while andrographolide showed interaction with the COX-2 binding pocket via two hydrogen bonds (Fig. 2b) which were formed between oxygen atom of andrographolide’s lactone as HB acceptor and hydrogen atom in the Arg513’s amine and His90’s pyridine (Table 1).
| Table 1: | Interaction of Andrographolide with COX-2 binding pocket in comparison with SC-558 |
 | |
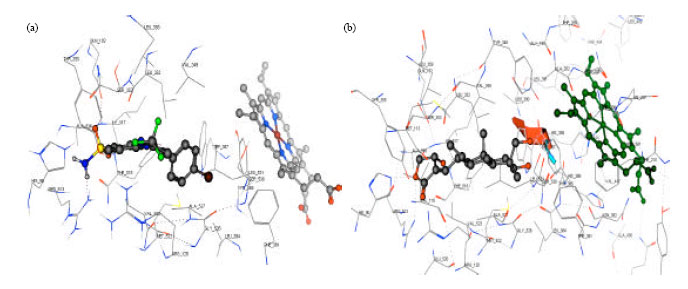 | |
| Fig. 2: | (a) Interaction of SC-558 and (b) andrographolide in COX-2 binding pocket. Hydrogen bonds were showed by dotted lines |
The energy needed for these interactions, either for SC-558 or andrographolide, -10.7717 and -11.7963 kcal mol–1, respectively, were small and not significantly different (Table 1). These data suggested that the interaction of andrographolide with cyclooxygenase site of COX-2 enzyme was favorable.
In vitro study: Andrographolide has been reported to have anti-inflammatory activity by suppressing inducible nitric oxide synthase expression in RAW 264.7 cells (Chiou et al., 2000), prevents oxygen radical production by human neutrophils (Shen et al., 2002), inhibits NF-kappaB activation (Xia et al., 2004) and reduced COX-2 expression induced by platelet activating factor and N-formyl-methionyl-leucyl-phenylalanine in HL60/neutrophils (Hidalgo et al., 2005). The anti-inflammatory activity of this compound on stimulated human fibroblast cells has not been reported.
Fibroblast is a type of cell that synthesizes the extracellular matrix and collagen and plays critical role in wound healing. During inflammation, any damage which occurs in the tissue will stimulate the mitosis of fibroblasts.
In this study andrographolide showed inhibitory activity on COX-2 expression in human fibroblast cells induced by lipopolysaccharide in the concentration range between 2 to 200 μM (IC50 = 4 μM), which was 0.7 times than acetosal’s (IC50 = 2.8 μM) (Fig. 3a, b).
An unexpected result showed from this work. Preincubation of fibroblast cells with low concentrations of acetosal induced the production of prostaglandin, but with the increasing of acetosal concentration, PGE2 level decreased. Andrographolide at increasing concentration (up to 200 μM) did not stimulate PGE2 production. These data confirmed that andrographolide’s anti-inflammatory activity was occurred via inhibition of COX-2 expression.
The behavior of acetosal in correlation with PGE2 production was compared with Morgan et al. (2009) which concluded that COX-2 appeared to be differentially regulated in aspirin-sensitive patients. Morgan and her colleagues observed that aspirin and LPS increased COX-2 expression on blood monocytes of aspirin-induced asthmatic patients, a finding in contrast with the lack of an effect of the same stimuli on COX-2 expression on monocytes from healthy subjects. They found significantly higher COX-2 expression levels after stimulation with LPS and aspirin (mean 78.8, range 44.9-92.3; p = 0.0002) in comparison to LPS alone (mean 65.9%, range 33.6-82.6) in aspirin-induced asthmatic patients (Morgan et al., 2009).
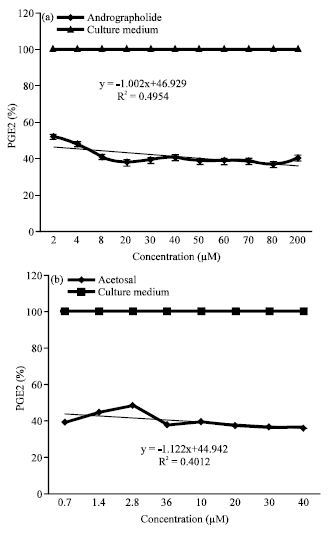 | |
| Fig. 3: | Spectrometric analysis of PGE2 production in fibroblast cells with and without (a) Andrographolide and (b) Acetosal |
CONCLUSIONS
Andrographolide showed interaction with arginine and histidine in the cyclooxygenase site of COX-2 and reduced PGE2 production in human fibroblast cells induced by lipopolysaccharide. Its inhibitory activity to COX-2 enzyme was 0.7 times than acetosal’s, but unlike acetosal, andrographolide did not stimulate PGE2 production.
ACKNOWLEDGMENTS
We would like to thank Dr. Endang Winiarti (Research Laboratory, Faculty of Dentistry, Universitas Indonesia, Jakarta, Indonesia) for her kind gift, human fibroblast cells used in this project, the Rector of Universitas Padjadjaran for his continuous supports and Institute Technology Bandung, Indonesia that has partially funded this work via IM HERE grant 2010. The authors have declared no conflict of interest.
REFERENCES
- Chiou, W.F., C.F. Chen and J.J. Lin, 2000. Mechanisms of suppression of inducible nitric oxide synthase (iNOS) expression in RAW 264.7 cells by andrographolide. Br. J. Pharmacol., 129: 1553-1560.
CrossRef - Dilber, S.P., S.L. Dobric, Z.D. Juranic, B.D. Markovic, S.M. Vladimirov and I.O. Juranic, 2008. Docking studies and anti-inflammatory activity of β-hydroxy-β-arylpropanoic acids. Molecules, 13: 603-615.
CrossRef - Gierse, J.K., J.J. McDonald, S.D. Hauser, S.H. Rangwala, C.M. Coboldt and K. Seibert, 1996. A single amino acid difference between cyclooxygenase-1 (COX-1) and −2 (COX-2) reverses the selectivity of COX-2 specific inhibitors. J. Biol. Chem., 271: 15810-15814.
CrossRef - Hidalgo, M.A., A. Romero, J. Figueroa, P. Cortes, I.I. Concha, J.L. Hancke and R.A. Burgos, 2005. Andrographolide interferes with binding of nuclear factor-kB to DNA in HL-60-derived neutrophilic cells. Br. J. Pharmacol., 144: 680-685.
CrossRef - Kurumbail, R.G., A.M. Stevens, J.K. Gierse, J.J. McDonald and R.A. Stegenman et al., 1996. Structural basis for selective inhibition of cyclooxygenase-2 by anti-inflammatory agents. Nature, 384: 644-648.
PubMedDirect Link - Morgan, T., N. Bajrovic, M. Silar, M. Kosnik and P. Korosec, 2009. Aspirin-induced COX-2 overexpression in monocytes of aspirin-intolerant patients. Int. Arch. Allergy Immunol, 149: 378-384.
CrossRef - Nanduri, S., V.K. Nyavanandi, S.S.R. Thunuguntla, S. Kasu and M.K. Pallerla et al., 2004. Synthesis and structure-activity relationship of andrographolide analogues as cytotoxic agents. Bioorg. Med. Chem. Lett., 14: 4711-4717.
CrossRef - Shen, Y.C., C.F. Chen and W.F. Chiou, 2002. Andrographolide prevents oxygen radical production by human neutrophils: Possible mechanisms involved in its anti-inflammatory effect. Br. J. Pharmacol., 135: 399-406.
CrossRef - Xia, Y.F., B.Q. Ye, Y.D. Li, J.G. Wang and X.J. He et al., 2004. Andrographolide attenuates inflammation by inhibition of NF-kB activation through covalent modification of reduced cysteine62 of p50. J. Immunol., 173: 4207-4217.
Direct Link - Xu, Y., R.L. Marshall and T.K.S. Mukkur, 2006. An investigation on the antimicrobial activity of Andrographis paniculata extracts and andrographolide in vitro. Asian J. Plant Sci., 5: 527-530.
CrossRefDirect Link - Young, J.M., S. Panah, C. Satchawatcharaphong and P.S. Cheung, 1996. Human whole blood assays for inhibition of prostaglandin G/H synthases-1 and -2 using A23187 and lipopolysaccharide stimulation of thromboxane B2 production. Inflamm. Res., 45: 246-253.
CrossRef