
Theerawit Poeikhampha
Department of Animal Science, Faculty of Agriculture, Kasetsart University, Bangkok, Thailand
Chaiyapoom Bunchasak
Department of Animal Science, Faculty of Agriculture, Kasetsart University, Bangkok, Thailand
Journal of Applied Sciences
Year: 2010 | Volume: 10 | Issue: 14 | Page No.: 1471-1475







ABSTRACT
This study was conducted to evaluate the effect of SG supplementation in batch culture of pig cecal digesta (in vitro) on pH value, ammonia and Short Chain Fatty Acids (SCFAs) concentration. The dried ileal and cecal digesta of commercial crossbred female pigs were used as the substrate of fermentation and inoculums, respectively. The trial was divided into 5 groups according to the SG supplementations at 0, 1,250, 2,500, 5,000 or 7,500 ppm of dried ileal digesta (each group contained 6 replications). The experimental design was completely randomized design. The SG was added into each tube containing 8 mg of ileal digesta and 20 mL of inoculums. The tubes were immediately filled with CO2 and incubated at 39°C for 24 h. During the fermentation period, the samples of incubated fluid were collected at 4, 8 and 24 h for determination of pH and ammonia concentration, while SCFAs were determined at 24 h. At 8 and 24 h of fermentation period, SG supplementation (7,500 ppm) reduced pH value (p<0.01) and ammonia concentrations (p<0.05) in batch culture of porcine cecal digesta. At the end of fermentation period (24 h), supplementing SG evidently increased acetic acid, propionic acid and total SCFAs (p<0.01), although significant increase in butyric acids was not observed. It may imply that SG could be the useful prebiotic to depress pH and ammonia in cecal digesta of pigs due to high production of SCFAs.
PDF Abstract XML References Citation
Received: March 08, 2010;
Accepted: April 22, 2010;
Published: June 10, 2010
How to cite this article
Theerawit Poeikhampha and Chaiyapoom Bunchasak, 2010. Effect of Sodium Gluconate on pH Value, Ammonia and Short Chain Fatty Acids Concentration in Batch Culture of Porcine Cecal Digesta. Journal of Applied Sciences, 10: 1471-1475.
DOI: 10.3923/jas.2010.1471.1475
URL: https://scialert.net/abstract/?doi=jas.2010.1471.1475
DOI: 10.3923/jas.2010.1471.1475
URL: https://scialert.net/abstract/?doi=jas.2010.1471.1475
INTRODUCTION
In recent year, using prebiotics as feed additive is extremely focused since antibiotics are being banned around the world because consumers demand for a food chain that is free of drugs. Particularly, European Union has completely banned use of antibiotics as growth promoters since January 2006 due to spreading of bacteria that are resistant to such antimicrobials (Bager et al., 1997; Phillips et al., 2004).
Sodium Gluconate (SG) that derives from incomplete oxidation of glucose may be used as a good source of prebiotics. In addition, it has been promoted as functional food for human and defined as non digestible food ingredient that beneficially affects the host by selectively stimulating the growth and/or the activity of one or limited member of bacteria in lower gastrointestinal tract (Gibson and Roberfroid, 1995). It has been reported that SG is poorly digested and absorbed in the small intestine, but it can be utilized by lactic acid bacteria such as Lactobacillus sp. and Bifidobacterium sp., subsequent produce lactic acid and short chain fatty acids (SCFAs) in the large intestinal tract (Asano et al., 1994). Lactic acid and SCFAs are substrates to form butyric acid by Megasphaera elsdenii (Tsukahara et al., 2002) and the butyric acid clearly stimulates growth of epithelial cells in the intestine of pigs (Biagi et al., 2006; Scheppach et al., 1995). Therefore, supplementation of SG in diet may indirectly produce energy to support the growth of epithelial cell.
It is interesting to know whether SG supplementation can be beneficial to piglets as prebiotic or not. Effect of supplementing SG in diet of piglets is limited and examination of SG in animal (in vivo) is costly, therefore this study was conducted to determine the effect of SG as prebiotic using batch culture of porcine cecal digesta (in vitvo).
MATERIALS AND METHODS
The in vitro study was conducted at Animal Research Farm, Department of Animal Science, Faculty of Agriculture, Kasetsart University, Thailand in August to September, 2009. The experimental animals were kept, maintained and treated in adherence to accepted standards for the humane treatment of animals. In this study, 10 new born female commercial crossbred piglets (Duroc x Large White x Landrace) were used; the diet and water without antimicrobial agents were provided ad libitum until the body weight reached approximately 25 kg. Broken rice and soybean based diet was used. The composition of the diet is shown in Table 1. Piglets were raised in open house system. The house was cleaned two days interval, while the feces of piglets were removed everyday.
Collection of ileal and caecal digesta: The ileal digesta from 5 pigs were used as the substrate of fermentation. Five pigs were putdown and 200 g of ileal digesta was instantaneously removed from each pig, pooled and kept in a sealed plastic bag at 4°C. The ileal digesta was homogenized and dried at 60°C overnight and subsequently analyzed for chemical composition according to AOAC (2000) methods. The chemical analysis of dried ileal digesta were crude protein 8.6%, crude fat 3.06%, calcium 0.93%, total phosphorus 0.51% and ash 2.14%; as presented in Table 1.
The caecal digesta from 5 pigs were used as the inoculums, therefore remaining five pigs were putdown and 600 g of the caecal digesta was immediately removed from each pig, pooled and kept in a sealed plastic bag at 39°C. The caecal digesta were diluted with buffer (ratio = 1:2) and filtered through 4 layers of cheesecloth.
| Table 1: | Composition of the diet and chemical analysis of dried ileal digesta (in vivo) |
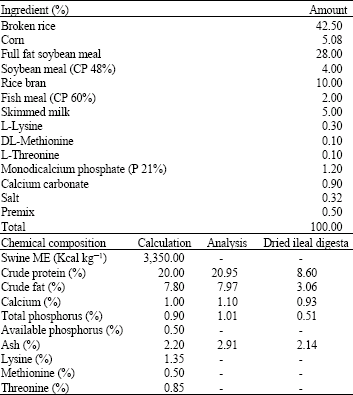 | |
| Premix content; Vitamin A 4 MIU, D 0.64 MIU, E 24,000 IU, K3 1.4 g, B1 0.6 g, B2 0.3 g, B6 0.75 g, B12 14 mg, Nicotinic acid 20 g, Pantothenic acid 10 g, Folic acid 0.44 g, Biotin 0.04 g, Choline 60 g, Fe 45 g, Cu 40 g, Mn 15 g, Zn 40 g, Co 0.2 g, I 0.4 g, Se 0.06 g, Carrier Added to 1 kg | |
The pH of the inoculums was adjusted to 6.7 and immediately used.
The buffer composition was as follows: 116.7 mmol of NaHCO3 , 7.6 mmol of KCl , 0.4 mmol of CaCl2·6H2O, 26.0 mmol of Na2HPO4·12H2O , 11.5 mmol of NaCl, 0.5 mmol of MgSO4·7H2O in 1 L of distilled water. Buffer pH was adjusted to 6.7 by adding 3 N HCl. The buffer solution was kept at 39°C and flushed with CO2 for 20 min before use (McDougall, 1948).
Fermentation process: There were 5 treatments according to SG supplementation levels (0, 1,250, 2,500, 5,000 or 7,500 ppm of dried ileal digesta). Each treatment was divided into six replications of four 50 mL tubes. Each tube was filled with 8 mg of ileal digesta and 20 mL of inoculums and induced to anaerobic condition by immediately flushing with CO2. The SG was added to each tube before sealed by plastic caps and each tube was shaked and incubated at 39°C for 24 h.
Chemical analysis: Samples of fermentation fluid were collected from each tube after 4, 8 and 24 h of fermentation period according to Biagi et al. (2006) and Tsukahara et al. (2002). The pH was determined using pH meter (pH meter model PH17-SS, IQ Scientific Instruments, Inc., Carlsbad, CA, USA) and ammonia concentration was determined by commercial test kit (Urea/BUN-Color, Biotechnical, Bangkok, Thailand).
Each samples of fermentation fluid collected at 24 h was mixed with 6 N HCl (ratio 5:1) to stop the fermentation and centrifuged for 15 min at 14,000 g at 4°C. Then supernatants were collected in order to determine the concentration of Short Chain Fatty Acids (SCFAs) such as acetic acid, propionic acid and butyric acid by gas chromatography in the following manner; 1 μL of the supernatant was injected into the silica capillary column (DB-Wax, J and W 30 mx0.25 mm i.d.). The GC-2010 High-end Gas Chromatograph (Shimadzu, Tokyo, Japan) was used. The GC oven was temperature-programmed from 50 to 220°C at a rate of 4°C min–1. The carrier gas (He) flow rate was 1.0 mL min–1 and a split ratio was 1:20. The temperatures of the injection port and detector were 225 and 250°C, respectively.
Statistical analysis: All data were statistically analyzed using Analysis of Variance (ANOVA) of SAS (SAS Institute, 1988). The differences between the means of groups were separated by Duncan’s Multiple Range Test (Duncan, 1955). Statements of statistical significance were based on p<0.05. All statistical analyses were carried out in accordance with the method of Steel and Torrie (1980).
RESULTS
The pH values: The pH values in batch culture of pig caecal digesta incubated with different concentrations of SG are shown in Table 2, supplementation of SG into dried ileal digesta did not significantly influence to the pH values within 4 h of fermentation (average of pH value was ~5.20). At 8 h of fermentation period, SG supplementation at 1,250 ppm significantly declined the pH value compare to the control group (5.21 vs. 5.17) and supplementation of SG at 2,500 or 5,000 or 7,500 ppm induced lower pH values than the 1,250 ppm supplementation group (p<0.01). At 24 h, the supplementations of SG at 2,500 or 5,000 ppm induced lower pH than the control and the 1,250 ppm supplementation groups. Moreover, supplementation of SG at 7,500 ppm gave the lowest pH value in batch culture (p<0.01).
Ammonia concentration: The ammonia concentrations in batch culture of pig caecal digesta incubated with different concentrations of SG supplementation are shown in Table 3, at 4 h of fermentation, the ammonia concentration (~0.21 mmol L–1) was not influenced by SG supplementation. At 8 and 24 h of fermentation, however, SG supplementation at 7,500 ppm significantly decreased the concentration of ammonia compared to the 0, 1,250 and 2,500 ppm SG supplementation groups (p<0.05). There was no significant difference of ammonia concentration between 5,000 and 2,500 ppm or among 0, 1,250 and 2,500 ppm supplementation groups.
Short Chain Fatty Acids (SCFAs) concentration: The yield of acetic acid, propionic acid, butyric acid and total SCFAs (sum of the acetic acid, propionic acid and butyric acid) in batch culture of pig caecal digesta with different concentrations of supplementing SG after 24 h of fermentation period are shown in Table 4. Supplementations of SG (1,250, 2,500, 5000 and 7500 ppm) clearly increased acetic acid concentration (p<0.01). Furthermore, increments of SG linearly increased the concentration of propionic acid (R2= 0.93; p<0.01), subsequently adding SG at 7,500 ppm produced highest yield of propionic acid (15.69 mmol L–1). However, the concentration of butyric acid was only slightly increase by SG supplementation. Focusing on total SCFAs, batch culture of pig caecal digesta without SG supplementation produced lowest concentration of total SCFAs (20.50 mmol L–1). The concentration were sharply increased by SG supplementation at 1,250 ppm (33.98 mmol L–1), then increased gradually and linearly from 35.11, 38.07 and 39.96 mmol L–1 by supplementation of SG at 2,500, 5,000 and 7,500 ppm, respectively.
| Table 2: | The pH values in batch culture of pig caecal digesta incubated with different concentrations of sodium gluconate |
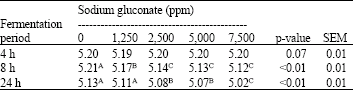 | |
| Means within a row with different superscripts differ highly significant (p<0.01) | |
| Table 3: | Ammonia concentrations in batch culture of pig caecal digesta incubated with different concentrations of sodium gluconate |
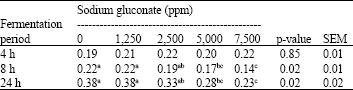 | |
| Means within a row with different superscripts differ significantly (p<0.05) | |
| Table 4: | Short chain fatty acids concentrations in batch culture of pig caecal digesta incubated with different concentrations of sodium gluconate for 24 h |
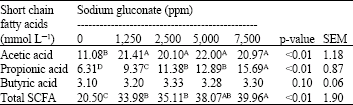 | |
| Means within a row with different superscripts differ highly significant (p<0.01). Total short chain fatty acids: sum of the concentrations of acetic acid, propionic acid and butyric acid | |
DISCUSSION
After 4 h of fermentation periods (at 8 and 24 h), lowering pH values in batch culture of pig caecal digesta by SG supplementation may come from high growth and/or high activities of beneficial microorganisms (lactobacillus bacteria) (Fuller, 1997; Gibson and Roberfroid, 1995) and Streptococci (Tsukahara and Ushida, 2001). This infers that within 4 h of the fermentation period, production of SCFA may be low since the pH value was not changed. This phenomenon indicates that SG is slowly degraded by microorganism in the batch culture. Similarly, Michel et al. (1996) reported that SCFA production from an algal oligosaccharide was delayed and mainly occurred between 12 and 24 h of fermentation. According to the in vivo study, SG significantly decreased pH in the intestine of pigs by stimulation of beneficial bacteria growth after 6 week of feeding period (Biagi et al., 2006). Modulating fermentation and lowering pH in large intestine of animal can be implemented by several ways such as supplementation of organic acids, prebiotics or probiotics in diet, while various mechanisms and effects of each feed additive have been reported (Vanbelle et al., 1990; Steed and Macfarlane, 2009; Casadei and Biagi, 2002). It seems that SG supplementation reduces pH through indirect mechanism (fermentation process), while the organic acid such as potassium diformate directly reduces the pH and population of coliform bacteria in the batch culture of pig digesta fermentation (in vitro) (Knarreborg et al., 2002). Hence, supplementation of SG at 5,000 to 7,500 ppm of dried ileal digesta may significantly promote acid bacteria growth, reduce pH and consequently inhibit the growth of pathogenic bacteria.
In piglet within first 3 or 4 week after birth, the protein digestion is not complete due to an insufficient activation of pepsinogen that occurs rapidly at pH 2 and very slowly at pH 4 (Taylor, 1962) and gastric acid secretion does not reach appreciable levels until 3 to 4 week after weaning (Cranwell and Moughan, 1989). Therefore, this occurrence decreases the protein digestion and protein utilization of piglets. Moreover, the undigested protein may leads to increase proteolysis, consequently releasing toxic substances such as ammonia and amines (Russell et al., 1983; Biagi et al., 2006). In this study, supplementation of SG probably decreases the potentially toxic substances since supplementing SG at 5,000 or 7,500 ppm significantly decreased ammonia concentration in batch culture during 8 and 24 h of fermentation period. Lowering ammonia concentration by SG supplementation was also reported in both studies of in vitro and in vivo (Biagi et al., 2006). Hence, stimulation of beneficial microorganism growth by supplementing SG may decrease the proteolysis of protein, or the N compound in batch culture.
At 24 h of fermentation, acetic acid, propionic acid and total SCFAs concentrations were dramatically increased by SG supplementation. SG supplementation at a level of 7,500 ppm of dried ileal digesta increased concentrations of acetic acid, propionic acid, butyric acid and total SCFAs compared to the control by 189, 248, 106 and 181%, respectively. This mean that using SG as substrates of fermentation mainly produce propionic acid and acetic acid (at least during 24 h of fermentation period), while increase of acetic acid (not propionic acid) in batch culture of pig cecal digesta induced by SG supplementation at 24 h of fermentation period was reported by Tsukahara et al. (2002). Biagi et al. (2006) reported that production of total SCFAs was linearly increased by SG supplementation after 4 h of adaptation periods. On the other hand, 4 h of fermentation period did not acidify the pH of batch culture in our study, thus we suggested that in order to produce high SCFAs product, the adaptation period of the SG fermentation should be longer than 4 h. This assumption is confirmed by Tsukahara et al. (2002), who also reported that SG is fermented slowly.
Butyric acid is a relatively minor component compared to acetic acid and propionic acid in the lumen of the large intestine (Kameue et al., 2004), whereas it is considered to be the most effective for gut health. For example, butyric acid is major energy sources for epithelial cells of the large intestine (Roediger, 1980; Scheppach et al., 1995), stimulate mucus release and epithelial cell proliferation (Slomiany et al., 1980; Sakata, 1987; Young and Gibson, 1995). Acetic acid and propionic acid are used as substrate to form butyric acid by acid utilizing bacteria such as Megasphaera elsdenii (Tsukahara et al., 2002). Linear increasing butyric acid caused by SG fermentation in in vitro has been reported by Biagi et al. (2006).
In this study, however, the butyric acid was only slightly increased by SG supplementation; therefore the effect of SG on butyric acid production remains inconclusive. In order to produce high butyric acid yield, the fermentation period may needs longer than 24 h and further evaluation is necessary.
ACKNOWLEDGMENTS
The authors gratefully acknowledge that the funding has come from Sumitomo Chemical Co., Ltd., Japan and extend thanks to the staff of Department of Animal Science, Kasetsart University and of Faculty of Veterinary Science, Mahidol University Thailand.
REFERENCES
- Asano, T., K. Yuasa, K. Kunugita, T. Teraji and T. Mitsuoka, 1994. Effects of gluconic acid on human faecal bacteria. Microbial Ecol. Health Dis., 7: 247-256.
CrossRefDirect Link - Biagi, G., A. Piva, M. Moschini, E. Vezzali and F.X. Roth, 2006. Effect of gluconic acid on piglet growth performance, intestinal microflora and intestinal wall morphology. J. Anim. Sci., 84: 370-378.
Direct Link - Fuller, R., 1977. The importance of lactobacilli in maintaining normal microbial balance in the crop. Br. Poult. Sci., 18: 85-94.
CrossRefDirect Link - Gibson, G.R. and M.B. Roberfroid, 1995. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr., 125: 1401-1412.
CrossRefPubMedDirect Link - Kameue, C., T. Tsukahara, K. Yamada, H. Koyama, Y. Iwasaki, K. Nakayama and K. Ushida, 2004. Dietary sodium gluconate protects rats from large bowel cancer by stimulating butyrate production. J. Nutr., 134: 940-944.
Direct Link - Knarreborg, A., N. Miquel, T. Granli and B.B. Jensen, 2002. Establishment and application of an in vitro methodology to study the effects of organic acids on coliform and lactic acid bacteria in the proximal part of the gastrointestinal tract of piglets. Anim. Feed Sci. Technol., 99: 131-140.
Direct Link - McDougall, E.I., 1948. Studies on ruminant saliva. I. The comparison and output of sheep`s saliva. Biochem. J., 43: 99-109.
Direct Link - Phillips, I., M. Casewell, T. Cox, B. de Groot and C. Friis et al., 2004. Does the use of antibiotics in food animals pose a risk to human health? A critical review of published data. J. Antimicrob. Chemother., 53: 28-52.
CrossRefDirect Link - Casadei, P.G. and G. Biagi, 2002. An organic acid blend can modulate swine intestinal fermentation and reduce microbial proteolysis. Can. J. Anim. Sci., 82: 527-532.
Direct Link - Roediger, W.E., 1980. Role of anaerobic bacteria in the metabolic welfare of the colonic mucosa in man. Gut, 21: 793-798.
PubMedDirect Link - Russell, J.B., C.J. Sniffen and P.J. van Soest, 1983. Effect of carbohydrate limitation on degradation and utilization of casein by mixed rumen bacteria. J. Dairy Sci., 66: 763-775.
Direct Link - Scheppach, W., H.P. Bartram and F. Richter, 1995. Role of short-chain fatty acids in the prevention of colorectal cancer. Eur. J. Cancer, 31: 1077-1080.
CrossRefDirect Link - Slomiany, B.L., V.L.N. Murty and A. Slomiany, 1980. Isolation and characterization of oligosaccharides from rat colonic mucus glycoprotein. J. Biol. Chem., 255: 9719-9723.
Direct Link - Taylor, W.H., 1962. Proteinases of the stomach in health and disease. Physiol. Rev., 42: 519-553.
Direct Link - Tsukahara, T. and K. Ushida, 2001. Organic acid profiles in feces of pigs with pathogenic or non-pathogenic diarrhea. J. Vet. Med. Sci., 63: 1351-1354.
Direct Link - Tsukahara, T., H. Koyama, M. Okada and K. Ushida, 2002. Stimulation of butyrate production by gluconic acid in batch culture of pig cecal digesta and identification of butyrate-producing bacteria. J. Nutr., 132: 2229-2234.
Direct Link - Vanbelle, M., E. Teller and M. Focant, 1990. Probiotics in animal nutrition: A review. Arch. Anim. Nutr., 40: 543-567.
CrossRefPubMedDirect Link