
F. Zarrineh
Department of Exercise Physiology, Science and Research Branch, Islamic Azad University, Tehran, Iran
F. Salami
Department of Physical Education, Faculty of Physical Education and Sport Sciences, Tarbiat Moallem University, Tehran, Iran
H.N. Bakht
Department of Exercise Physiology, Science and Research Branch, Islamic Azad University, Tehran, Iran
M. Salavati
Department of Physical Therapy, University of Social Welfare and Rehabilitation, Tehran, Iran
M. Hedayati
Research Institute for Endocrine Sciences, Shahid Beheshti University, Tehran, Iran
Journal of Applied Sciences
Year: 2009 | Volume: 9 | Issue: 13 | Page No.: 2469-2474







ABSTRACT
To assess the change in plasma level of 22 and 20 kDa GH isoforms following acute aerobic exercise and chronic aerobic training, 30 male athletes in two pubertal and pre-pubertal groups were studied. Acute aerobic exercise consisted of 30 min running at 65% Vo2max on the treadmill. Plasma was sampled 30 min before and immediately after the aerobic exercise and assayed for total concentrations. These measurements were repeated at the end of six weeks of aerobic training (30 min running on the treadmill, three times weekly, at 70-80% Vo2max). In both groups, acute aerobic exercise was followed by increased 22 kDa and decreased 20 kDa concentration. These changes were larger in the pre-pubertal group. Chronic aerobic training was associated with decreased 22 kDa and increased 20 kDa response to acute aerobic exercise in both groups. There was no significant change in the rest levels of 22 and 20 kDa concentration in either groups following chronic aerobic training.
PDF Abstract XML References Citation
How to cite this article
F. Zarrineh, F. Salami, H.N. Bakht, M. Salavati and M. Hedayati, 2009. Effect of Acute and Chronic Aerobic Training on Plasma GH Isoforms Concentration in Pubertal and Pre-Pubertal Male Athletes. Journal of Applied Sciences, 9: 2469-2474.
DOI: 10.3923/jas.2009.2469.2474
URL: https://scialert.net/abstract/?doi=jas.2009.2469.2474
DOI: 10.3923/jas.2009.2469.2474
URL: https://scialert.net/abstract/?doi=jas.2009.2469.2474
INTRODUCTION
Human Growth Hormone (hGH), also called somatotropin, a pleiotropic polypeptide, exerts multiple effects on the metabolic and anabolic processes of the human body (Nindl et al., 2003). The heterogeneity of circulating GH has been recognized (Baumann, 1999). The hGH-N gene expresses the main pituitary molecular weight variant in the GH family, known as 22 kDa form. This protein is 191 amino acids in length (monomeric 22 kDa represents nearly 43% of all plasma GH). The next most abundant form is the 20 kDa molecule (monomeric 20 kDa represents about 8% of all plasma GH) formed by deletion of amino acids 32-46 during an alternative mRNA splicing (Nindl, 2007). The evidence favors the concept that the short peptide potentiates the physiological effects of insulin while the larger peptide has anti-insulin properties (Lewis et al., 2000).
Aerobic exercise of approximately 15 min duration has been shown to stimulate GH release which depends on some factors such as gender, age, type of exercise, body composition and exercise intensity (Wideman et al., 2000; De Palo et al., 2008). A single bout of endurance exercise in the form of continuous cycle ergometry at approximately 80% Vo2max for 20 min led to an increase in all molecular isoforms of GH such as 22 and 20 kDa at the end of acute exercise in pubertal males (Wallace et al., 2001). However, there is limited data on the response of 22 and 20 kDa in pre-pubertal boys. On the other hand, findings on the GH responses to chronic aerobic training when compared to acute aerobic exercise have brought conflicting results. Some studies showed that sustained aerobic training limits acute exercise-induced GH release (Stokes et al., 2000). A response down regulation was evident within the first 3 weeks of training in young pubertal men (Weltman et al., 1997). In contrast, although 16 weeks of aerobic training yielded a significant training effect (i.e., increased Vo2 peak) no change in GH response to acute exercise occurred (Kanaley et al., 1999). Given their different physiological properties, it would be necessary to investigate the possible differential response of GH dominant molecular isoforms to endurance exercise.
Therefore, the present study was designed to determine if chronic aerobic training evokes changes in the response of prevalent GH isoforms (22 and 20 kDa) to a single bout of aerobic exercise. The second aim was to compare these changes between pubertal and pre-pubertal men. It was hypothesized that the acute response of GH isoforms to aerobic exercise will change following chronic aerobic training in both groups.
MATERIALS AND METHODS
Subjects: Fifteen pubertal male soccer players (17±2 year, 67±5 kg, 178±4 cm) and 15 pre-pubertal male soccer players (12±2 year, 58±3 kg, 170±5 cm) were recruited. The subjects were asked to read and sign an informed consent form before participation. This study was approved by a research institutional review board. The procedures followed were in accordance with the Helsinki Declaration of 1975. The main exclusion criteria were the presence of major medical problems including orthopedic, cardiovascular and endocrinological conditions, smoking and drug abuse. All subjects were screened for nutritional habits by aerobic registered dietician 2 weeks before initial testing to assure normal dietary intakes and food behavior and to eliminate confounding influences of aberrant diets. All subjects had habitually consumed adequate energy including 55% carbohydrate, 25% protein and 15% fat.
Protocol: Three days before participating in the study, all subjects performed Bruce test on a treadmill (Technogym Co., Gambettola, Italy) to determine Vo2max. A single bout of acute aerobic exercise with 3 min duration (including 5 min warm up followed by 25 min running on the treadmill at 65% Vo2max) was administered three days before the initiation of chronic aerobic training. Chronic aerobic training lasted 6 weeks, three sessions per week and 30 min per session. Each session consisted of 5 min warm up followed by 25 min running on the treadmill at 70-80% Vo2max. Exercise intensity was increased by 5% every 2 weeks. As the second bout, acute aerobic exercise was repeated 3 days after the completion of chronic aerobic training. All the sessions were carried out at about 10:00 am. Each subject had a light breakfast (2-3 slices of toasted bread with honey and a cup of tea, milk or juice) about 2 h before the exercise.
Blood samples were collected 30 min before (rest level) and immediately after the end of acute aerobic exercise. A sample of 5 mL venous blood was obtained by sterile syringe from the left antecubital vein at the sitting position in EDTA containing test tube as an anticoagulant. Blood was centrifuged at room temperature for 15 min at 800 x g. Suitable aliquots of the plasma specimens were immediately stored at -20°C in a freezer. When plasma samples were defrosted before the analysis, each sample was recentrifuged (10 min, 2000 x g).
Biochemical analysis: Plasma levels of 22 and 20 kDa concentrations were determined by specific enzyme-linked immunosorbent assay kits (ELISA). In the 20 kDa ELISA, 0.1 mL assay buffer (150 mmol L-1 phosphate-buffered saline containing 1% BSA, 1 mol L-1 NaCl and 10 mg L-1 heterophilic blocking reagent) and 0.025 mL of standards (Mitsui Pharamaceuticals, Inc., Tokyo, Japan) or plasma samples were added to monoclonal anti -20 kDa antibody-precoated microtiter plates (anti-hGH-antibody DO5, Mitsui Parmaceuticals, Inc., Tokyo, Japan). It was incubated for 2 h at the room temperature. After thorough washing (0.01 mol L-1 Tris-HCl , pH 8.0, containing 0.05% Tween-20), 0.1 mL (0.5 mg L-1) peroxidase-labeled anti -20 kDa monoclonal antibody (POD-D14, Mitsui pharmaceuticals, Inc.) was added and incubated for 2 h at the room temperature. After a further washing step, 0.1 mL substrate solution (100 mmol L-1 citrate buffer containing 65 mg L-1 3/3’, 5/5’ -tetramethylbenzidine and 4 mmol L-1 H2O2, pH 3.8) was added and the plates were incubated for 30 min at the room temperature. The absorbance were read at 450 nm (reference length was 620 nm) after stopping the enzyme reaction with 0.1 mL H2SO4. The detection limit was 5 pg mL-1 and the cross reactions with 22 kDa, hPRL and human placental lactogen were less than 0.1%. In 22 kDa ELISA, microtiter plates were precoated with the anti-hGH antibody (anti-hGH antibody Mitsui Pharmaceuticals, Inc., Tokyo, Japan). Other procedures were the same as described above. The detection limit was 50 pg mL-1 and the cross-reactions with 20 kDa, hPRL and human placental lactogen were less than 0.1%. The coefficient of variation of both assays was less than 5%.
Statistical analysis: Paired t-tests were used to compare the Pre-training and Post-training rest levels and to analyze the changes of plasma GH isoforms concentration within each bout of acute aerobic exercise (exercise-induced change score). A separate 2x2 analysis of variance with one between-subject factor (group: pubertal and pre-pubertal) and one within-subject factor (time: Pre-training and Post-training) was used to analyze the exercise-induced change scores of each dependent variable. Significant level was considered as p≤0.05. Statistical analysis was performed manually.
RESULTS
Paired t-test showed no significant difference between rest levels of pre-training and post-training sessions in either group (Table 1).
| Table 1: | Pre-training-Post-training difference of 22 and 20 kDa rest levels in pubertal and pre-pubertal groups |
 | |
| Values are expresed as Mean (SD); SD: Standard Deviation; Significant levels at p≤0.05 are in bold | |
| Table 2: | Pre-training and Post-training change scores of 22 and 20 kDa in pubertal and pre-pubertal groups |
 | |
| SD: Standard Deviation; Significant levels at p≤0.05 are in bold | |
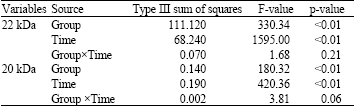
| Table 3: | Group by time analysis of variance for change scores of 22 and 20 kDa |
 | |
| Significant levels at p≤0.05 are in bold | |
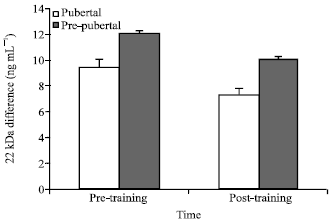 | |
| Fig. 1: | The response of plasma 22 kDa concentration to chronic aerobic training during a single bout of aerobic exercise in both pubertal and pre-pubertal groups |
For all sessions, a single bout of aerobic exercise increased 22 kDa and decreased 20 kDa concentrations significantly (Table 2).
For 22 kDa, there was no significant interaction between group and time (F1, 14 = 1.68, p = 0.21). Main effect for both group (F1,14 = 330.34, p<0.01) and time (F1,14 = 1595, p<0.01) factors was significant (Table 3).
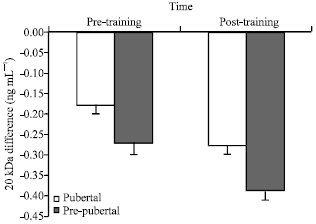 | |
| Fig. 2: | The response of plasma 20 kDa concentration to chronic aerobic training during a single bout of aerobic exercise in both pubertal and pre-pubertal groups |
Increment in 22 kDa during one session for pubertal was significantly lower than pre-pubertal group and for Post-training significantly lower than Pre-training session (Fig. 1).
For 20 kDa, no significant interaction was observed between group and time (F1,14 = 3.81, p = 0.06), but there were significant main effect for both group (F1,14 = 180.32, p<0.01) and time (F1,14 = 420.36, p<0.01) factors. Decrement in 20 kDa during one session for pre-pubertal was significantly more than pubertal group and for Post-training significantly more than pre training session (Fig. 2).
DISCUSSION
The results showed that a single bout of aerobic exercise significantly increased plasma 22 kDa concentration and decreased plasma 20 kDa concentration in pubertal and pre-pubertal groups. The changes in plasma 22 and 20 kDa concentration were larger in pre-pubertal group. Also, there was no change in the rest level of plasma 22 and 20 kDa concentration following chronic aerobic training in both groups. Chronic aerobic training decreased the 22 kDa and increased the 20 kDa response to a single bout of aerobic exercise.
A single bout of aerobic exercise increased plasma 22 kDa concentration immediately after exercise, which is in accordance with the result obtained by Wallace et al. (2001). Those studies which show increase in GH during a single bout of aerobic exercise (Wideman et al., 2000; Pritzlaff et al., 1999; De Palo et al., 2008) may support the above finding probably because 22 kDa is the most prevalent (43-73%) GH isoform (Nindl, 2007; Wallace et al., 2001). Aerobic exercise at 65% Vo2max may lead to a moderate elevation in blood lactate and hydrogen ion (H+) accumulation. These elevations have been showed to be significantly correlated with plasma GH response during 20 min of aerobic exercise (Gordon et al., 1994). So, it has been suggested that any stimulus for GH release associated with lactate elevation would be more likely to act through H+ accumulation (Stokes et al., 2005), which in turn H+ accumulation activates the neural afferent signal from metabolic receptors and might participate in catecholamine release (Kjaer, 1989; Weltman et al., 1996). A decrease in pH in contracting muscles due to accelerated lactate production and associated metabolic changes may stimulate the sympathetic out flow by neural afferent signals from muscle metabolic receptors causing rapid release of catecholamine (Kjaer, 1989). Catecholamine released following afferent signals from muscle metabolic receptors might, in turn, play a role in the regulation of GH and its most prevalent isoform (22 kDa) release (Weltman et al., 1996). In human exercising, a significant positive correlation between plasma nor-adrenaline and serum GH concentrations has been identified (Chwalbinska-Moneta et al., 2005; Weltman et al., 2000). In animal studies, catecholamine has been shown to directly stimulate GH release from rat gland at the in vitro level (Giustina and Veldhuis, 1998). It has also been proposed that aerobic exercise induces GH responses through activation of the central cholinergic system, resulting in a reduction in hypothalamic somatostatin release (Maas et al., 2000). In addition, GH release may be stimulated directly by motor centers involved in exercise (Ju, 1999).
In the present study, plasma 20 kDa concentration decreased immediately after a single bout of aerobic exercise which conflicted with that of Wallace et al. (2001). They demonstrated that 20 kDa concentration increased with a single bout of aerobic exercise during recovering period in contrast to present study in which 20 kDa concentration was assessed immediately after exercise. Considering decreased insulin concentration during aerobic exercise (Hymer et al., 2005) the change in 20 kDa concentration may represent its insulin-like action (Smal et al., 1987).
The changes in plasma 22 and 20 kDa concentrations following a single bout of aerobic exercise were larger in pre-pubertal than pubertal group. Since, the rest levels of GH in pre-pubertal boys are higher (Ishikawa et al., 1999) their GH response to aerobic exercise may be different from those of pubertal men. The exact mechanisms responsible for these differences and the interaction of factors involved, needs to be clarified by further studies.
There was no change in the rest levels of plasma 22 and 20 kDa concentration following chronic aerobic training in both groups. This is in agreement with the results of other studies showing that training has no effect on the rest level of GH concentration (Zaccaria et al., 1999). Any interpretation about the resting level of GH concentration must be made cautiously because of pulsatile nature of its secretion (Stokes, 2003).
Finally, the decreased 22 kDa and increased 20 kDa response to a single bout of aerobic exercise after chronic aerobic training is consistent with Veldhuis and Yoshida (2000). It seems that mechanisms regulating the response of both GH isoforms to a single bout of aerobic exercise have been suppressed after chronic aerobic training. This may be due to a combination of reduced GH secretion and enhanced GH clearance (Weltman et al., 1997). The half-life of endogenous GH is shorter in exercising than resting individuals (Thompson et al., 1993). On the other hand, aerobic training can increase both the number and sensitivity of β2 adrenergic receptors and decrease catecholamine release (Chwalbinska-Moneta et al., 2005; Mazzeo, 2000). This in turn excites somatostatin secretion which leads to the inhibition of GH release (Giustina and Veldhuis, 1998; Vries et al., 2002).
Further investigations are needed to compare the effects of aerobic and resistive training on GH isoforms, the effects of training on GH isoforms other than 22 and 20 kDa and the changes of GH isoforms during recovery period after exercise. Also, more parameters like changes in glucagon, insulin levels may be included in future studies to explore the possible mechanisms behind the changes obsereved for GH isoforms during pre and post aerobic exercise.
In conclusion, it was demonstrated that the changes of plasma level of GH isoforms induced by a single bout of aerobic exercise decreased after a peiod of aerobic training, with a similar pattern in both pubertal and pre-pubertal male athletes.
ACKNOWLEDGMENT
This research was supported by Science and Research Branch, Islamic Azad University, Tehran, Iran. The authors speciallly thank Masood Mazaheri, Ph.D. PT, for his invaluable comments.
REFERENCES
- Baumann, G., 1999. Growth hormone heterogeneity in human pituitary and plasma. Horm. Res., 51: 2-6.
CrossRefDirect Link - Chwalbinska-Moneta, J., B. Kruk, K. Nazar, K. Krzeminski, H. Kaciuba-Uscilko and A. Ziemba, 2005. Early effects of short-term endurance training on hormonal responses to graded exercise. J. Physiol. Pharmacol., 56: 87-99.
Direct Link - De Palo, E.F., G. Antonelli, R. Gatti, P. Spinella and E. Cappellin, 2008. Effects of two different types of exercise on GH/IGF axis in athletes. Is the free/total IGF-1 ratio a new investigative? Clin. Chim. Acta, 387: 71-74.
Direct Link - Giustina, A. and J.D. Veldhuis, 1998. Pathophysiology of the neuroregulation of growth hormone secretion in experimental animals and the human. Endocrnol. Rev., 19: 717-797.
CrossRefDirect Link - Gordon, S.E., W.J. Kraemer, N.H. Vos, J.M. Lynch and H.G. Knuttgen, 1994. Effect of acid-base balance on the growth hormone response to acute high-intensity cycle exercise. J. Applied Physiol., 76: 821-829.
Direct Link - Ishikawa, M., S. Yokoya, K. Tachibana, Y. Hasegawa and T. Yasuda et al., 1999. Serum levels of 20-kilodalton human growth hormone (GH) are parallel those of 22-kilodalton human GH in normal and short children. J. Clin. Endocrinol. Metab., 84: 98-104.
CrossRefDirect Link - Ju, G., 1999. Evidence for direct neural regulation of the mammalian anterior pituitary. Clin. Exp. Pharmacol. Physiol., 26: 757-759.
CrossRefDirect Link - Kanaley, J.A., M.M. Weatherup-Dentes, E.B. Jaynes and M.L. Hartman, 1999. Obesity attenuates the growth hormone response to exercise. J. Clin. Endocrinol. Metab., 84: 3156-3161.
CrossRefDirect Link - Kjaer, M., 1989. Epinephrine and some other hormonal responses to exercise in man: With special reference to physical training. Int. J. Sports Med., 10: 2-15.
CrossRefDirect Link - Lewis, U.J., Y.N. Sinha and G.P. Lewis, 2000. Structure and properties of members of the hGH family: A review. Endocrine J., 47: S1-S8.
Direct Link - Maas, H.C.M., W.R. de Vries, I. Maitimu, E. Bol, C.Y. Bowers and H.P. Koppeschaar, 2000. Growth hormone responses during strenuous exercise: The role of GH-releasing hormone and GH-releasing peptide-2. Med. Sci. Sports Exerc., 32: 1226-1232.
Direct Link - Mazzeo, R.S., 2000. Aging, immune function and exercise: Hormonal regulation. Int. J. Sports Med., 1: S10-S13.
Direct Link - Nindle, B.C., W.J. Kraemer, J.O. Marx, A.P. Tuckow and W.C. Hymer, 2003. Growth hormone molecular heterogeneity and exercise. Exerc. Sport Sci. Rev., 31: 161-166.
Direct Link - Nindl, B.C., 2007. Exercise modulation of growth hormone isoforms: Current knowledge and future directions for the exercise endocrinologist. Br. J. Sports Med., 41: 346-348.
Direct Link - Pritzlaff, C.J., L. Wideman, J.Y. Weltman, R.D. Abbott and M.E. Gutgesell et al., 1999. Impact of acute exercise intensity on pulsatile growth hormone release in men. J. Applied Physiol., 87: 498-504.
Direct Link - Smal, J., J. Closset, G. Hennen and P. De Meyts, 1987. Receptor binding properties and insulin-like effects of human growth hormone and its 20 kDa-variant in rat adipocytes. J. Biol. Chem., 262: 11071-11079.
Direct Link - Stokes, K., 2003. Growth hormone responses to sub-maximal and sprint exercise. Growth Horm. IGF Res., 13: 225-238.
Direct Link - Stokes, K., M. Nevill, J. Frystyk, H. Lakomy and G. Hall, 2005. Human growth hormone responses to repeated bouts of sprit exercise with different recovery periods between bouts. J. Applied Physiol., 99: 1254-1261.
Direct Link - Thompson, D.L., J.Y. Weltman, A.D. Rogol, D.L. Metzger, J.D. Veldhuis and A. Weltman, 1993. Cholinergic and opioid involvement in release of growth hormone during exercise and recovery. J. Applied Physiol., 75: 870-878.
Direct Link - Veldhuis, J.D. and K. Yoshida, 2000. Impact of chronic training on pituitary hormone secretion in the human. Sports Endocrinol., 1: 57-76.
CrossRefDirect Link - Vries, W.D., S.A. Abdesselam and T.J. Schers, 2002. Complete inhibition of hypothalamic somatostatin activity is only partially responsible for the growth hormone response to strenuous activity. Metabolism, 51: 1093-1096.
Direct Link - Wallace, J.D., R.C. Cuneo, M. Bidlingmaier, P.A. Lundberg and L. Carlsson et al., 2001. The response of molecular isoforms of growth hormone to acute exercise in trained adult males. J. Clin. Endocrinol. Metab., 86: 200-206.
Direct Link - Chwalbinska-Moneta, J., H. Krysztofiak, A. Ziemba, K. Nazar, H. Kaciuba-Uscilko, 1996. Threshold increases in plasma growth hormone in relation to plasma catecholamine and blood lactate concentrations during progressive exercise in endurance-trained athletes. Eur. J. Applied Physiol. Occup. Physiol., 73: 117-120.
CrossRefDirect Link - Weltman, A., J.Y. Weltman, C.J. Womack, S.E. Davis, J.L. Blumer, G.A. Gaesser and M.L. Hartman, 1997. Exercise training decreases the growth hormone (GH) response to acute constant-load exercise. Med. Sci. Sports Exerc., 29: 669-676.
CrossRefDirect Link - Weltman, A., C.J. Pritzlaff and L. Wideman, 2000. Exercise dependent growth hormone release is link to markers of heightened central adrenergic outflow. J. Applied physiol., 89: 629-635.
Direct Link - Wideman, L., J.Y. Weltman, J.T. Patrie, C.Y. Bowers, N. Shah, S. Story, J.D. Veldhuis and A. Weltman, 2000. Synergy of L-arginine and GHRP-2 stimulation of growth hormone in men and women: modulation by exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol., 279: R1467-R1477.
Direct Link - Zaccaria, M., M. Varnier, P. Piazza, D. Noventa and A. Ermolao, 1999. Blunted growth hormone response to maximal exercise in middle-aged versus young subjects and no effect of endurance training. J. Clin. Endocrinol. Metab., 84: 2303-2307.
CrossRefDirect Link