
J. Goma-Tchimbakala
Institut de Developpement Rural, Universite Marien Ngouabi, BP 13502 Brazzaville, Congo
S. Makosso
Institut de Developpement Rural, Universite Marien Ngouabi, BP 13502 Brazzaville, Congo
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 23 | Page No.: 4346-4353







ABSTRACT
A comparative study was carried out in Mayombe, between the soil of natural forest and the soil under four Terminalia superba plantations of 7, 12, 32 and 48 year old. In each plantation type and natural forest composite soil samples were taken in 0-10, 10-20, 20-30, 30-40 and 40-50 cm layers. The goal was to assess the impact of reforestation on soil organic matter and its biological characteristics. Statistically differences between the sites were tested using the Analysis of Variance (ANOVA). The results showed that there was a general decrease of soil organic matter in the profile of all sites except in the 48 year old plantation for total nitrogen. In the surface soil the carbon content and total nitrogen were respectively, 22.2 mg g-1 and 1.56 μg g-1 in the forest. The carbon content was between 14.9 and 23.5 mg g-1 while total nitrogen was between 1.31 and 2.24 μg g-1 in the plantations. The microbial biomass carbon (Cmic) was 732 μg g-1 soil in the forest, it varied between 461 and 740 μg g-1 in the plantations. The metabolic quotient (qCO2) was between 3.6 mg C-CO2 mg-1 Cmic day-1 and 4.8 mg C-CO2 mg-1 Cmic day-1 in plantations. The qCO2 in forest was 3.6 mg C-CO2 mg-1 Cmic day-1.
PDF Abstract XML References Citation
How to cite this article
J. Goma-Tchimbakala and S. Makosso, 2008. Soil Organic Matter and Biological Changes in a Natural-to Planted
Forest Succession: Terminalia superba Plantations Grown on Deforested
Plots in Congo. Journal of Applied Sciences, 8: 4346-4353.
DOI: 10.3923/jas.2008.4346.4353
URL: https://scialert.net/abstract/?doi=jas.2008.4346.4353
DOI: 10.3923/jas.2008.4346.4353
URL: https://scialert.net/abstract/?doi=jas.2008.4346.4353
INTRODUCTION
Tree plantations cover an area of more than 50,000 ha in the littoral and Mayombe regions in the South of Republic of Congo. They are planted with both exotic and indigenous species and are generally monospecific (Goma-Tchimbakala and Bernhard-Reversat, 2006). Terminalia superba (T. superba) is the main native species used for reforestation in the Mayombe. Studies in the tropics have shown significant changes in soil organic matter following conversion of natural forest into cultivation and these changes have been shown to affect soil fertility (Dominy et al., 2002; Yimer et al., 2007). The changes that occurred after deforestation and subsequent cropping are decreases in plant available nutrients (Lu et al., 2002), decreases in microbial activity (Sahani and Behera, 2001), increases in bulk density, soil erosion and runoff (Rasiah and Kay, 1995) and decreases in porosity, infiltration and water-holding capacity (Lu et al., 2002; Sahani and Behera, 2001). However, when some of these degraded soil were brought under well managed pasture, significant improvements in degraded properties occurred after the introduction of pasture (Rasiah and Kay, 1995; Rasiah et al., 2004). In south of Republic of Congo, it is not known if similar improvements would occur under T. superba plantations which were grown on deforested soil. In agreement with Rasiah et al. (2004) such information is necessary for several reasons. First, if natural forest regrowth is poor after abandonment for substantially long period of time, then the information on soil properties may help, at least partially, to identify the factor (s) controlling regrowth or reestablishment. Second, this information may be useful to assess the resiliency of soils in different tropical environment to recover from man induced changes and finally to determine the time required for substantial recovery to occur. The latter information is crucial if monetary benefits are considered for abandonment and subsequent reforestation. Long-term impacts of afforestation and reforestation on the distribution of soil organic matter and biological activities have not been reported for the South of Republic of Congo. Research regarding the effects of reforestation on soil organic matter dynamics and biological properties with T. superba plantation aging will help to determine the potential of recovery of these soils for carbon sequestration and biological properties.
The purpose of this study was to assess changes in soil organic matter and biological properties due to reforestation with T. superba in Mayombe in comparison with natural forest soil characteristics.
MATERIALS AND METHODS
Site description: The sites were previously described by Goma-Tchimbakala and Bernhard-Reversat (2006). Briefly they are located at Bilala in the Mayombe at 4°31`S, 12°4`E and above mean sea level 300 m altitude. Mean annual rainfall is 1250 mm. Relative humidity is high all over the year close to 85%. The temperature ranges from 28°C in the rainy season to 18°C in the dry season. The soil is desaturated ferralitic and lies on sandy-clayey or clayey-sandy material resulting from the weathering of cretaceous green rock and dolomitic limestone. In this study the clay content has narrow range from 27 to 29% in the surface soil (Table 1). T. superba plantations were established by the National Reforestation Service (SNR) after clear cutting the forest and burning all plants residues. The tree spacing is 10x10 m. Four plantations of 7, 12, 32 and 48 years old were chosen for this study (Fig. 1). The undergrowth vegetation was well developed in the planted plots and the Fabaceae, Annonaceae, Euphorbiaceae, Moraceae, Marantaceae and Menispe-rmaceae were the most numerous plant families.
| Table 1: | Average particle size distribution of the surface soil under T. superba plantations and natural forest |
 | |
The nearby natural forest was used for comparison with the planted plots. The most represented tree families in the natural forest are the Cesalpiniaceae, followed by the Rubiaceae and the Euphorbiaceae. All the stems larger than 10 cm Diameter at Breast Height (DBH) were listed together with the plants of the understory (Goma-Tchimbakala and Bernhard-Reversat, 2006).
Soil sampling: Soil samples were collected from ten systematically located squares (1 m2 for each square) in each plantation type and natural forest. In each square two composite soil samples were taken in 0-10, 10-20, 20-30, 30-40 and 40-50 cm layers. Each composite soil sample was combined from four cores from a layer. Samples for chemical analysis were air-dried and then were passed through a 2 mm mesh soil sieve.
Microbial activity and microbial biomass: For the measurement of basal respiration, 100 g of a moist soil sample was weighed into 1000 mL jar, adjusted to 55% of its water-holding capacity and incubated during 7 days at 28°C in the dark. The CO2 produced was absorbed in 50 mL 0.2 M NaOH solution and determined by titration of the excess NaOH with 0.2 M HCl. The CO2 respired was analysed periodically at 2, 4 and 7 days.
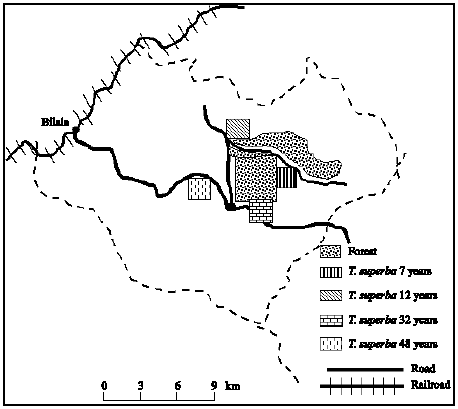 | |
| Fig. 1: | Situation of the plantations and natural forest (the plantation plots are not drawn at the scale) |
Potential Basal Respiration (BR), expressed in mg CO2-C g-1 soil day-1, was calculated when microbial respiration had stabilized, i.e., during the last 3 days of incubation. After 7 days` incubation, 4 soil sub-samples (25 g each) were used to determine the microbial biomass. Microbial biomass carbon (Cmic) and microbial biomass nitrogen (Nmic) were estimated by the chloroform fumigation-extraction method (Vance et al., 1987). Two portions were fumigated with ethanol-free chloroform during 24 h. After removal of CHCl3, C was extracted from fumigated and unfumigated soil samples with 0.5M K2SO4 with a soil: extractant ratio of 1:5 (w:v) for 30 min on a rotating shaker. After shaking and centrifugation at 5100 rpm during 10 min, the soil suspension was filtered with a 0.22 μm filter (Millipore Corporation, Belford, MA 01730) and the filtrate was collected. Dissolved Organic Carbon (DOC) was determined on both the fumigated and unfumigated soil extracts. The Cmic was calculated according to the equation: Cmic = EC/KEC (Vance et al., 1987), where Ec was the difference between extractable C from fumigated samples and unfumigated samples and KEC = 0.38. The Nmic was calculated as Nmic = EN/KEN, where EN was the difference between extractable N from fumigated and unfumigated samples and KEN = 0.54 (Brookes et al., 1985; Joergensen and Mueller, 1996). The metabolic quotient (qCO2) is the ratio of basal respiration to Cmic of soil, expressed in mg CO2-C(mgCmic)-1 day-1 . The microbial quotient (qCmic), which is the ratio of Cmic to Corg of a soil, is expressed in mg Cmic(g(Corg))-1.
Chemical analysis: The analysis were carried out in the laboratory of IRD (Institute of Research for Development) at Pointe-Noire with standard methods. Carbon was analyzed by the modified Walkey and Black method. Total nitrogen was determined using the Kjeldahl procedure. The extractable C for microbial biomass (DOC) was analyzed by the HachTM methods (Jirka and Carter, 1975): Two millimeters of filtered DOC were heated for 2 h with potassium dichromate, the organic matter reducing the dichromate ion (CrO7-2) to green chromic ion (Cr3+). The amount of Cr3+ produced was determined with colorimetric method using DR/890 colorimeter. The extractable N was measured after mineralization of the N to NH4+. Two milliliters of concentrated H2SO4 and 9 drops of H2O2 were added to 2 mL of the filtered soil extracts and heated at 200°C during 24 h. After the mineralization process, the N was analyzed by the Nessler method.
Data analysis: We assessed changes in soils under plantations by computing the difference between mean values of individual soil properties under different T. superba plantations and the values of same soil properties under natural forest and expressed as a percentage of the value under the natural forest. These percent changes were used as references of soil degradation or improvement across the investigated layers.
Statistically significant differences were tested using the Analysis of Variance (ANOVA). The Protected Least Significant Difference (PLSD) Fisher test was used for mean separation when the analysis of variance showed statistically differences. All statistical analysis were performed using Stat View 5.0 (SAS Inst. Inc., 1998).
RESULTS
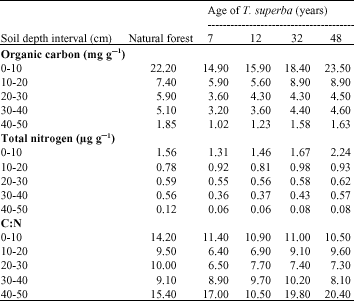
Organic carbon and total nitrogen in the profile: The results showed that deforestation affected contents of soil organic carbon and total nitrogen significantly (p<0.001; Table 2, 3). The soil surface in the 7 year old plantation had the weakest soil organic content than all other plantations (Table 2). Soil organic carbon content increased with the aging of plantations and reached 23.5 mg g-1 under the mature plantation (48 years old). Comparison of mean differences among sites revealed that the 48 year old plantation had similar soil organic carbon with the natural forest (Table 4).
| Table 2: | Depth distribution of soil organic carbon, total nitrogen and C:N ratio under T. superba plantations and natural forest |
 | |
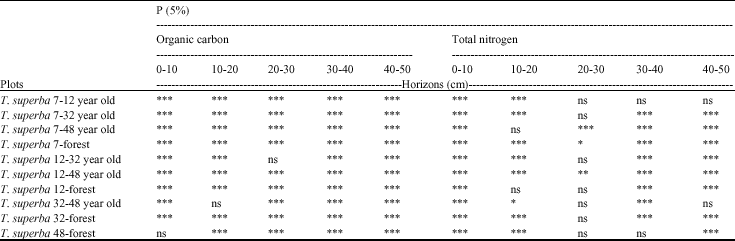
| Table 3: | ANOVA of soil organic carbon and total nitrogen |
 | |
| *** p<0.0001; ** p<0.001; * p<0.05 | |
Soil organic content in the 0-10 cm layer in these two sites was higher than in the plantations between 7 and 38 year old (Table 2). Total nitrogen showed the same trend as soil organic content in the plantations. The 7 year old plantation had significantly low total nitrogen content while the 48 year old plantation had the highest content. However comparison between the sites showed that the soil of natural forest had low nitrogen content than plantation of 32 and 48 year old. However total nitrogen in natural forest was not different with the 12 year old plantation. The low total nitrogen content recorded under natural forest induced a higher C/N ratio than in plantations (Table 2).
The vertical distribution of organic carbon and total nitrogen appeared to differ in soils under plantations and between the soil under plantations and natural forest (Table 2). In the depth 10-20 cm the mean of soil organic carbon were significantly higher (p < 0.05) in the 32 and 48 year old plantations than in the 7-12 year old plantations and the natural forest. On other hand differences were not significant between the 7 and the 12 year old plantation. After, organic carbon decreased regularly in all other layers until 50 cm in depth. Comparison between the sites showed that the 7 year old plantation had the lower organic carbon content and the higher content was in natural forest.
The distribution of total nitrogen in the profile was more complex (Table 2). Total nitrogen was similar under plantations in the l0-20 cm layer and the content was significantly high than in natural forest (Table 3). The same trend was observed in 20-30 cm concerning the plantations of 7, 12, 32 years old and natural forest, whereas the 48 year old plantation had highest total nitrogen content (Table 4). In the two last layers the natural forest and the 48 year old plantation had high total nitrogen content than all other plantations. The C:N ratio decreased in general from soil surface to 30-40 cm in depth. This decline was followed by an increase in 40-50 cm layer. However C:N ratios did not differ significantly in plantations and forest.
Microbial biomass and microbial activity: The microbial biomass carbon (Cmic) varied between 461 and 740 μg g-1 soil whereas the microbial biomass nitrogen (Nmic) was between 87 and 114 μg g-1 soil (Table 5). Comparable Cmic and Nmic were recorded in natural forest, semi mature and mature plantations (Table 5). On the other hand the 7 year old plantation and the 12 year old plantation had similar and significantly lower Cmic and Nmic than in the 48 year old plantation and the natural forest (p<0.0001, Table 5, 6). The lower Cmic to Nmic ratio was in the 7 year old plantation while the higher ratio was observed in the 48 year old plantation.
| Table 4: | Comparison of mean differences between the sites |
 | |
| ***p<0.0001; **p<0.001; *p<0.05; ns: non significant | |
| Table 5: | Microbial activity, microbial biomass and microbial indices under T. superba plantations and natural forest soils (mean and standard deviation) |
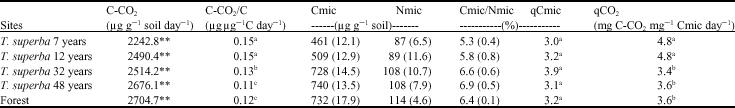 | |
| Cmic: microbial biomass carbon; Nmic: microbial biomass nitrogen; C: organic carbon; The number with the same letter in the column were not different at level 0.05 | |
There were significant differences between all the other plantations (p<0.0001, Table 5). The Cmic expressed versus soil organic C (qCmic) showed no clear trend and varied in narrow range of 3.0-3.9% (Table 5). The mineralized C was higher in the natural forest and the 48 year old plantation than in the 7-12 year old plantations, (p<0.0001, Table 5). Expressed as percent of soil organic C, the mineralized C was significantly higher in the soils under the 7-12 year old plantations than under the natural forest and the 48 year old plantation (Table 5).
Percent changes under T. superba plantations: Table 7 showed that loss of organic carbon was between 20.2 and 44.8% in the 7-12 year old plantations. It was between 13.7 and 27.1% in the 32 year old plantation. In these 3 plantations the losses occurred in all layers, except in the 10-20 cm layer in the 32 year old plantation where the increase was observed. In the 48 year old plantation there was an increase of organic carbon in the 0-10 and 10-20 cm layers while the loss taken place in the 20-30 cm, 30-40 cm and 40-50 cm layers. Comparison between plantation types revealed that loss of organic carbon decrease with the increase of the plantation age. Apart in the 10-20 cm layers, the loss of total nitrogen occurred in the soil profile in the 7-12 year old plantations (Table 7). In these 2 plantations the strong loss was between 33.9 and 50% in the 30-40 and 40-50 cm in depth. The surface soil presented positive changes (7.1 to 25.6%) in the 32 year old plantations while in the deep layers loss of total nitrogen varied between 1.7 and 33.3%.
| Table 6: | ANOVA of microbial biomass |
 | |
| *** p<0.0001; ** p<0.001; * p<0.05; ns: non significant | |
| Table 7: | Percent changes in soil carbon and nitrogen under T. superba plantations (differences between mean of soil carbon and nitrogen under plantations and value under forest expressed as a percentage of value under forest) |
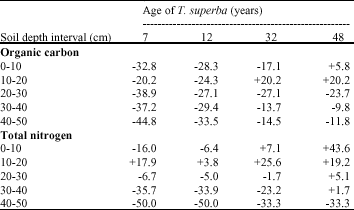 | |
| + Increase; -Loss | |
Except in the layer 40-50 cm there was a positive change in the profile of the 48 year old plantation, the gain varied between 1.8 and 43.6%. Comparison between plantation types showed that loss of nitrogen in the surface soil occurred only in the 7-12 year old plantations while in the 32-48 year old plantations nitrogen content increased.
DISCUSSION
Soil organic carbon and total nitrogen: The findings indicated that surface soil under the 48 year old plantation had significantly higher organic carbon and total N than in soils under the 32, 7 and 12 year old plantations, which might be a result of higher organic matter accumulation due to increased of litter fall and root biomass and reduced litter decomposition rates (Goma-Tchimbakala and Bernhard-Reversat, 2006; Saik et al., 1998; Yimer et al., 2007). Apart in the soil under the 48 year old plantation, the amount of organic matter under plantations was lower than in the natural forest due to the lack of litter return, high mineralization of organic matter following deforestation, losses of organic matter by water erosion (Paul et al., 2002; Jaiyeoba, 2003; Yimer et al., 2007). Deforestation accelerates soil organic matter loss by changing microclimate and exposure to microbial decomposition (Harmand and Pity, 2001). The amount of organic carbon in the surface soil under young and semi-mature plantations was between 17.1 and 32.8% less important compared to original amount in natural forest. The depletion of total N was in the range between 6.4 and 16.0%. In these two cases the loss of soil organic matter was more pronounced under the 7 year old plantation. The same phenomena had observed by others during cultivation or reforestation (Kotto Same et al., 1997; Saik et al., 1998; Xu et al., 2000). In the studies of these authors the loss reached respectively, 31% of organic carbon and 11% of total nitrogen. In ferralitic soils of Sierra Leone, two years after deforestation and planting Brams (1971) noted a loss of 40% of total nitrogen. Harmand (1997) observed, under planted cassia and eucalypts, that fast loss of carbon and nitrogen occurred only up to 40 cm in depth and that variations of organic matter became very small beyond this limit. In the depth, the distribution of soil organic carbon and total N differed between the plantation ages. Soil organic carbon and total N decreased with each layer from 0 to 50 cm in two plantations 7 and 12 years old. The most pronounced decrease occurred for 20-30 cm layer in the 7 year old plantation and for 10-20 cm layer in the 12 year old plantation. In the semi-mature and mature plantations there was improve of carbon content in the subsurface due to the increase of root biomass and decrease of oxidation. The C:N ratio in the 0-10 cm layers was almost similar in all the planted stands. This constancy compared to natural forest may indicate that the soil organic matter quality was more affected during the succession than the organic mater quantity. Finally the level of organic matter reached that of the natural forest after several decades. The low nitrogen content of organic matter in the forest soil could result from its higher mineralization (Goma-Tchimbakala, 2003).
Microbial biomass and organic matter quality: Changes due to reforestation also concerned organic matter quality (Joergensen et al., 1995). The results of microbial biomass fall in the range of some tropical forests 0.27-0.79 mg Cmic g-1 soil. The disturbance caused by the reforestation disappeared with the aging and the microbial indices reached the same level as natural forest. This may due to in part by changes in soil organic matter induced by litter diversity input of mature plantation and natural forest than in young plantations with lower litter diversity. This is in agreement with the finding of Bardgett and Shine (1999). The microbial quotient (qCmic) gives an indication of the stability of an ecosystem. It provides early warning of soil quality deterioration or environmental change (Anderson, 2003; Yan et al., 2003; Freschet et al., 2008). In this study, qCmic values indicated that the microbial biomass of the 32 and 48 year old plantations was closely to of the forest and was related to the amount of soil organic carbon. This finding was supported by the high microbial biomass founded under forest, semi mature and mature plantations where the soil organic content was high. No significant differences in qCmic were observed between the plots, which may indicate that there was little variation in soil organic matter quality with the age of the organic matter input (Freschet et al., 2008). The qCmic expressed as a percentage were in the range, between 1 and 5%, reported in numerous studies for tropical soil (Mao et al., 1992; Priess and Folster, 2001; Salamanca et al, 2002). The CO2 production expressed the microbial activities. It was followed the same evolution as the microbial biomass for similar reasons. The metabolic quotient (qCO2) has been used as bioindicator for ecosystem succession (Insam and Haselwander, 1989) and environmental stress (Anderson and Domsch, 1993; Joergensen et al., 1995; Priess and Fölster, 2001). The metabolic quotient reflects the efficiency of the use of organic carbon by microorganisms (Knoepp et al., 2000). Many researchers had hypothezed that environmental stress like low soil organic matter quality or nutrient deficiency reduces organic carbon efficiency use increasing the mineralized carbon needed for the ecosystem (Wolters and Joergensen, 1991; Kassim et al., 1992; Agnelli et al., 2001). The decrease of qCO2 with the age indicated the presence of microbial populations with more capability to use carbon compounds. The low qCO2 recorded in the 48 year old plantation and forest suggested less adverse environmental conditions and higher efficiency use of the organic resources than in the 7-12 year old plantations. According with Agnelli et al. (2001), a more efficient microbial community evolves in a more complex system with the heterogeneous organic matter input. According to Anderson and Domsch (1985) it may indicate that microbial biomass has recover after disturbance and become more stable in mature plantation.
CONCLUSION
The results of this research show that a decrease in soil organic matter occurred more in the surface soil. The qualitative and quantitative changes in organic matter and microbial characteristics with aging led to conditions close to the initial forest. The general trends suggested that Terminalia superba plantations might not degrade and rather improve soil fertility for further cultivation or plantation if they are logged when 48 year old as was planed. Plant diversity in the undergrowth, multistrate structure, high nutrient turn over and high productivity could be involved in restoring fertility after forest clearing.
ACKNOWLEDGMENTS
The International Foundation of Science (IFS) provided financial support for this research in form of research grant (D/2542-1). The National Reforestation Service (Service National de Reboisement, SNR) is acknowledged for welcoming the fieldwork in their plantations. Kokolo A. and A.N.S. Mboussou Kimbangou M. Ndoundou-Hockemba are acknowledged for their assistance during the soils sampling. Soil analysis were carried out at the analysis laboratory of the Institute of Research for Development of Pointe-Noire. We thank Dr. Victor Mamonekene who checked the English of the first manuscript. We also thank the four anonymous reviewers which helped us to improve the manuscript.
REFERENCES
- Agnelli, A., F.C. Ugolini, G. Corti and G. Pietramellara, 2001. Microbial biomass C and basal respiration of fine earth and highly altered rock fragments of two forest soils. Soil Biol. Biochem., 33: 613-620.
CrossRef - Anderson, T.H., 2003. Microbial eco-physiological indicators to assess soil quality. Agric. Ecosyst. Environ., 98: 285-293.
CrossRef - Anderson, T.H. and K.H. Domsch, 1985. Determination of ecophysiological maintenance carbon requirements of soil micro-organisms in a dormant state. Biol. Fertil. Soils, 1: 81-89.
CrossRef - Anderson, T.H. and K.H. Domsch, 1993. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils. Soil Biol. Biochem., 25: 393-395.
CrossRefDirect Link - Bardgett, R.D. and A. Shine, 1999. Linkages between plant litter diversity, soil microbial biomass and ecosystem function in temperate grasslands. Soil Biol. Biochem., 31: 317-321.
CrossRefDirect Link - Brams, E.A., 1971. Continuous cultivation of West Africa soils: Organic matter diminution and effects of applied line and phosphorus. Plant Soil, 35: 401-414.
CrossRef - Brookes, P.C., A. Landman, G. Pruden and D.S. Jenkinson, 1985. Chloroform fumigation and release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem., 17: 837-842.
CrossRef - Dominy, C.S., R.J. Haynes and R. Van Antwerpen, 2002. Loss of soil organic matter and related soil properties under long-term sugarcane production on two contrasting soils. Biol. Fert. Soils, 36: 350-356.
CrossRef - Freschet, G.T., D. Masse, E. Hien, S. Sall and J.L. Chotte, 2008. Long-term changes in organic matter and microbial properties resulting from manuring practices in an arid cultivated soil in Burkina Faso. Agric. Ecosyst. Environ., 123: 175-184.
CrossRef - Goma-Tchimbakala, J., 2003. Organic matter and nitrogen dynamics in plantation of limba (Terminalia superba Engler et Diels) in Congo: Comparison with a natural forest. Univ. Pierre Marie Curie Paris VI, 1: 180-180.
Direct Link - Goma-Tchimbakala, J. and F. Bernhard-Reversat, 2006. Comparison of litter dynamics in three plantations of an indigenous timber-tree species (Terminalia superba) and a natural tropical forest in Mayombe, Congo. For. Ecol. Manage., 229: 304-313.
CrossRefDirect Link - Harmand, J.M., 1997. Rôle des espèces ligneuses à croissance rapide dans le fonctionnement biogéochimique de la jachère. Effets sur la restauration de la fertilité des sols ferrigineux tropicaux (Bassin de la Benoué au Nord Cameroun). These, Université Paris, 6: 246-246.
Direct Link - Insam, H. and K. Haselwandter, 1989. Metabolic quotient of the soil microflora in relation to plant succession. Oecologia, 79: 174-178.
CrossRef - Jirka, A.M. and M.J. Carter, 1975. Micro semi-automated analysis of surface and wastewaters for chemical oxygen demand. Anal. Chem., 47: 1397-1402.
PubMed - Jaiyeoba, I.A., 1998. Changes in soil properties related to conversion of savannah woodland into pine and eucalyptus plantations, Northern Nigeria. Land Degrad. Dev., 9: 207-215.
CrossRef - Jaiyeoba, I.A., 2003. Changes in soil properties due to continuous cultivation in Nigerian semiarid Savannah. Soil Tillage Res., 70: 91-98.
CrossRefDirect Link - Joergensen, R.G., T.H. Anderson and V. Wolters, 1995. Carbon and nitrogen relationships in the microbial biomass in soils in beech (Fagus sylvatica L.) forests. Biol. Fertil. Soils, 19: 141-147.
CrossRef - Joergensen, R.G. and T. Mueller, 1996. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the KEN value. Soil Biol. Biochem., 28: 33-37.
CrossRef - Kassim, G., J.P. Martin and K. Haider, 1992. Incorporation of a wide variety of organic substrate carbons into soil biomass as estimated by fumigation procedure. Soil Sci. Soc. Am. J., 45: 1106-1112.
Direct Link - Kotto-Same, J., P.L. Woomer, A. Moukam and L. Zapfack, 1997. Carbon dynamics in slash and burn agriculture and land use alternatives of the humid forest zone in Cameroon. Agric. Ecosyst. Environ., 65: 245-256.
CrossRef - Knoepp, J.D., D.C. Coleman, D.A. Crossley and J.S. Clark, 2000. Biological indices of soil quality: An ecosystem case study of their use. For. Ecol. Manage., 138: 357-368.
CrossRef - Lu, D., E. Moran and P. Mausel, 2002. Linking Amazonian secondary succession forest growth to soil properties. Land Degradation Dev., 13: 331-343.
CrossRefDirect Link - Mao, D.M., Y.W. Min, L.L Yu, R. Martens and H. Insam, 1992. Effect of afforestation on microbial biomass and activity in soils of tropical China. Soil Biol. Biochem., 24: 865-872.
CrossRef - Paul, K.I., P.J. Polglase, J.G. Nyakuengama and P.K. Khanna, 2002. Change in soil carbon following afforestation. For. Ecol. Manage., 168: 241-257.
CrossRef - Priess, J.A. and H. Fölster, 2001. Microbial properties and soil respiration in submontane forest of Venezuelian Guyana: Charateristics and response to fertilizer treatments. Soil Biol. Biochem., 33: 503-509.
CrossRef - Rasiah, V. and B.D. Kay, 1995. Runoff and soil loss as influenced by selected stability parameters and cropping and tillage practices. Geoderma, 68: 321-329.
CrossRef - Rasiah, V., S.K. Florentine, B.L. Williams and M.E. Westbrooke, 2004. The impact of deforestation and pasture abandonment on soil properties in the west tropics of Australia. Geoderma, 120: 35-45.
CrossRef - Sahani, U. and N. Behera, 2001. Impact of deforestation on soil physicochemical characteristics, microbial biomass and microbial activity of tropical soil. Land Degrad. Dev., 12: 93-105.
CrossRef - Salamanca, E.F., M. Raubuch and R.G. Joergensen, 2002. Relationships between soil microbial indices in secondary tropical forest soils. Applied Soil Ecol., 21: 211-229.
CrossRef - Saik, H., C. Varadachari and K. Gosh, 1998. Changes in carbon, nitrogen and phosphorus levels due to deforestation and cultivation: A case study in Simlipal National Park, India. Plant Soil, 198: 137-145.
CrossRef - Vance, E.D., P.C. Brookes and D.S. Jenkinson, 1987. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem., 19: 703-707.
CrossRefDirect Link - Wolters, V. and R.G. Joergensen, 1991. Microbial carbon turnover in beech forest soilat different stages of acidification. Soil Biol. Biochem., 23: 897-902.
CrossRef - Xu, D.P., Z.J. Yang, B. Dell and M. Gong, 2000. Effects of Site Management in Eucalyptus Urophylla Plantations in Guangdong Provinces, China. In: Site Management and Productivity in Tropical Plantation Forests, Nambiar, E.K.S., A. Tiarks, C. Cossalter and J. Ranger, (Eds.). Kerrela, India, pp: 31-40.
Direct Link - Yan, T.M., L.Z. Yang and C.D. Campbell, 2003. Microbial biomass and metabolic quotient of soils under different land use in the Three Gorges Reservoir area. Geoderma 115: 129-138.
CrossRef - Yimer, F., S. Ledin and A. Abdelkadir, 2007. Changes in soil organic carbon and total nitrogen contents in three adjacent land use types in the Bale Mountains, South-Eastern highlands of Ethiopia. For. Ecol. Manage., 242: 337-342.
CrossRef