
E. Farshadfar
College of Agriculture, Razi University, Kermanshah, Iran
S.H. Sabaghpour
Dryland Agricultural Research Institute, Sararood, Kermanshah, Iran
N. Khaksar
College of Agriculture, Razi University, Kermanshah, Iran
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 21 | Page No.: 3931-3937







ABSTRACT
In order to study the inheritance and genetic analysis of drought tolerance indicators a six generations of F1, F2, BC1 and BC2 of the cross Hashem cultivar x ICCV96029 were studied using generation mean analysis. Genetic variation was found for grain yield, biological yield, No. of pod/plant, No. of seed/plant, earliness and proline content. High heterosis was observed in the F1 hybrid for grain yield, biological yield, harvest index, No. of pod/plant and No. of seed/plant. Genetic analysis indicated dominance in the inheritance of grain yield, biological yield, harvest index, seed weight and No. of seed/plant, while over dominance gene action in the No. of pod/plant, earliness and proline content. Moderate narrow-sense heritability estimates were observed for biological yield, harvest index, seed weight, No. of seed/plant and proline content. Moderate genetic advance for grain yield and proline content indicated that direct and indirect selection through correlated response could be effective. The joint scaling test revealed additivex dominance = [ j ] for grain yield, biological yield and proline content, while duplicate epistasis ( additive x dominance = [ j ] and dominance x dominance = [ l ] ) were shown for No. of pod/plant and No. of seed/pod. Since several important characters are influenced by dominance and non-allelic gene interaction, it is advisable to delay selection to later generation with increased homozygosity.
PDF Abstract XML References Citation
How to cite this article
E. Farshadfar, S.H. Sabaghpour and N. Khaksar, 2008. Inheritance of Drought Tolerance in Chickpea (Cicer arietinum L.) Using Joint Scaling Test. Journal of Applied Sciences, 8: 3931-3937.
DOI: 10.3923/jas.2008.3931.3937
URL: https://scialert.net/abstract/?doi=jas.2008.3931.3937
DOI: 10.3923/jas.2008.3931.3937
URL: https://scialert.net/abstract/?doi=jas.2008.3931.3937
INTRODUCTION
Legumes are important source of good quality protein in the diets of people and are valuable as animal feed. Chickpea is the third most important grain legume in the world grown in an area of about 10 million ha with a production of 7.8 million tons (Gowda et al., 2005; Rubio et al., 2004). Chickpea is the most important legume in Iran and occupies nearly 64% of the food legume in Iran with 17-24 protein, 41-50.8 carbohydrates and a percentage of other nutrients and has low production cost, high adaptability and applying in crop rotation and nitrogen fixation (Yaghotipoor and Farshadfar, 2007).
As all cultivated plants, the main objective of chickpea growing is to grow high yield and high quality crops. Since genotypic and environmental factors are components determining yield and quality in plants, a primary aid should be the determination of effects of genotypic factors in selection. As the effect of environment on yield and quality in plants is not heritable, effects of genotypic factors on yield and related characters in plant breeding research need to be examined (Güler et al., 2001).
The development of cultivars or varieties, which can be adapted to a wide range of environmental stresses is the ultimate goal of plant breeders in chickpea. Among the environmental stresses drought is the second contributor to yield reduction after disease (Farshadfar et al., 2001, 2003, 2008). Improving drought resistance is, therefore a major objective in plant breeding programs for rainfed agriculture (Ehdaie et al., 1991; Ehdaie and Waines, 1993).
To formulate an efficient breeding program for developing drought-tolerance varieties, it is essential to understand the mode of inheritance, the magnitude of gene effects and their mode of action (Farshadfar et al., 2001, 2008; Iqbal et al., 2007 ). Many workers developed genetic models for the estimation of different genetic effects (Gamil and Saheal, 1986; Kearsey and Pooni, 2004). However, the majority of these genetic models are basically additive-dominance models or simply additive models. The epistatic or non-allelic interaction are largely ignored so as to have a simplified interpretation of genetic variation. But, it has now been established that such inter-allelic interaction are of frequent occurrence in the control of trait-expression for continuous variation. Thus, inferences drawn from additive models are likely to be based to an unknown extent. That is why, Jinks et al. (1969) suggested, it is no longer possible to justify the use of a biometrical genetical analysis which does not have a built-in test for epistasis. To be on safe side, it is, therefore, rather essential to test the presence or absence of non-allelic interaction, i.e., to carry out test of additivity or epistasis (Sharma, 1998; Kearsey and Pooni, 2004; Farshadfar et al., 2008 ).
More than one procedure are available to test the deviation from additive models, i.e., to test the presence of non-allelic interactions. These are: Wr-Vr tests of additivity, the triple test cross test of epistasis, computation of interaction per se and scalling test (Farshadfar et al., 2008; Bakheit et al., 2002; Bhatnagar et al., 2004; Viana, 2005). We know that phenotypic mean is consummated by additive (a), dominance (d) and interaction effects (i) of genes in point. The interaction effect is again of two kinds: (i) complementary (aa) and (ii) duplicate (ad and dd) at digenic level. The analysis of generation means provides the opportunity first to detect the presence or absence of epistasis (by scaling test) and when present, it measures them appropriately. It also determines the components of heterosis in terms of gene-effects and some other statistics, like potence ratio levels of dominance, number of effective factors, etc. (Rebetzke et al., 2006; Farshadfar et al., 2008).
However as all cultivated plants, the main objective of chickpea growing is to grow high yield and high quality crops. Since genotypic and environmental factors are components determining yield and quality in plants, a primary aid should be the determination of effects of genotypic factors in selection. As the effect of environment on yield and quality in plants is not heritable, effects of genotypic factors on yield and related characters in plant breeding research need to be examined (Güler et al., 2001).
The objective of the present investigation was the genetic analysis of quantitative indicators of drought tolerance in chickpea under rainfed condition using generation mean analysis.
MATERIALS AND METHODS
The experimental material consisted of six generations (P1, P2, F1, F2, BC1 and BC2) derived from crosses between Hashem cultivar (late maturing, tall, medium grain size) and the line ICCV96029 (early maturing, short in plant height and small grain size) in the experimental farm of Dryland Agricultural Research Institute, Sararood, Kermanshah, Iran in 2003 and 2004. The parents (P1 and P2), the first (F1) and second (F2) generation hybrids and the first (P1 x F1 = BC1) and second (P2 x F1 = BC2) backcrosses were grown in a randomized complete block design with three replications in the field under rainfed condition in 2005. Parents, F1, BC1 and BC2 were sown in 2 rows with 30 cm row to row and 10 cm plant to plant spacing. The research station ordination on 1351.6 altitude and 47° , 20` longitude and 34° , 20` latitude. Average precipitation was 408 mm and average temperature was 13.8 °. Soil texture of the field was silty-clay loam. The soil was fertilized at the rate of 20 kg P2O5 ha-1 before sowing. Weeds were removed whenever appeared. Heliotis pest was controlled with sevine (3 kg ha-1) and disease ascochyta blight was controlled using the fungicide karbendazim (1.5-2/1000). The characters studied including: Grain Yield (GY), Biological Yield (BY), Harvest Index (HI), No. of pod per plant (NPP), No. of seed per plant (NSP), No. of seed per pod (NSPP), 100 seed weight, earliness and Proline Content (PC).
Assessment of proline in leaves: Proline of leaves was determined by Bates et al. (1973) method. Data were measured at 520 mm by Bausch and Lomb spectro photometer 70. A standard cure; 1.9, 7.8, 15.62, 31.25 and 125 μg of proline were prepared. Proline content of treated extracts was calculated using the standard curve and recorded.
Statistical analysis: Analysis of variance, mean comparison and correlation analysis was done using SPSS and MSTAT-C softwares. Generation mean analysis was performed using the Mather and Jinks method (1982). In this method the mean of each characters is indicated as follows:
Y = m + α [ d ] + β [ h ] + α2 [ i ] +2 α β [ j ] + β2 [l] |
| Y | = | The mean of one generation |
| m | = | The mean of all generation |
| d | = | The sum of additive effects |
| h | = | The sum of dominance effects |
| i | = | The sum of additive x additive interaction (complementary ) |
| l | = | The sum of dominance x dominance interaction (duplicate ) |
| j | = | Sum of additive x dominance and α, β, 2αβ and β2 are the coefficients of genetic parameters. The genetic parameters ( m, [ d ], [ h ], [ I ], [ j ], [ l ] ) were tested for significance using a t-test |
The adequacy of the additive dominance model was determined by χ2-test. Broadsense (H2b) and narrowsense (H2n) heritability were estimated by Warner ( 1952 ) and Allard ( 1960 ) formulas:
Hb2 = [ VF2 – ( VP1 + VP2 + VF1 ) / 3 ] / VF2 |
Hn2 = [ 2VF2 – ( VBC1 + VBC2 ) ] / VF2 |
where, V = variance of P1, P2, F1, F2, BC1 and BC2 generations.
The components of F2 variance were obtained by the following formula (Farshadfar, 1998; Farshadfar et al., 2008):
D = 4VF2 – 2 (VBC1 + VBC2) H = 2VF2 – 2D - 2E E = 1/3 (VP1 + VP2 + VF1) F = VBC1 - VBC2 |
| D | = | Additive genetic variance |
| H | = | Dominance variance |
| E | = | Environmental component of variance |
| F | = | Correlation between D and H over all loci |
RESULTS AND DISCUSSION
The results of analysis of variance (Table 1) revealed that all the generations had highly significant differences for GY, BY, NPP, SW, NSP, earliness and proline content indicating the existence of genetic variation and possibility of selection for drought tolerance.
In fact the development of any plant breeding program is dependent upon the existence of genetic variability. The efficiency of selection and expression of heterosis also largely upon the magnitude of genetic variability present in the plant population (Singh and Narayanan, 1993; Singh and Chaudhary, 1999).
The results of mean comparison (Table 2) showed that Hashem cultivar (P1) was significantly different from ICCV96029 P2) for GY, BY, NPP, SW, NSP, earliness and PC. The F1 hybrid displayed the highest heterosis over the best parent for GY, BY, HI, NPP and NSP.
BC1 also displayed satisfactory heterosis for GY, BY, HI, NPP and NSP over the best parent. The outstanding performance of the F1 hybrid and BC1 could be due to the contribution of Hashem cultivar. The significant genetic distance between Hashem and ICCV96029 and the higher amount of PC in Hashem is an indicator of drought tolerance in Hashem cultivar. Sabokdast and Kheialparast (2003) confirmed that drought tolerant chickpeas had high amount of proline content. Trangressive segregation was observed for GY, NPP and NSP in the F2 generation. The degree of dominance (h/d), broad-sense (Hb) and narrow-sense (Hn) heritabilities, genetic advance (GA) and genetic components of variation are presented in Table 3.
The variation observed between the genotypes for the characters investigated exhibited that selection maybe effective for the improvement of drought tolerance (Umarahan et al., 1997; Farshadfar et al., 2001; Farshadfar et al., 2008), however, the selection efficiency is related to the magnitude of heritability and genetic advance (Jonson et al., 1955; Singh and Narayanan, 1993).
| Table 1: | Analysis of variance for the characters used for the genetic analysis of drought tolerance |
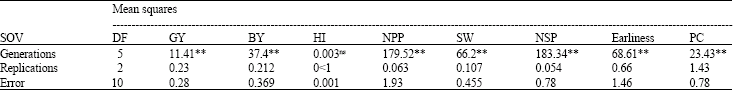 | |
| **, nsSignificant at 0.01 level of probability and non-significant, respectively | |
| Table 2: | Mean comparison of the characters under investigation |
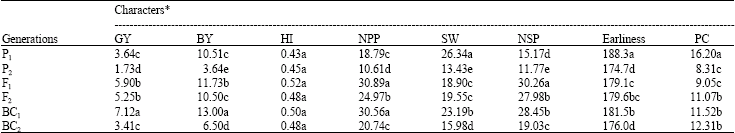 | |
| *Means with common letter(s) have no significant differences at 1% level of probability | |
| Table 3: | Genetic parameters and components of variation for various characters investigated under rainfed condition |
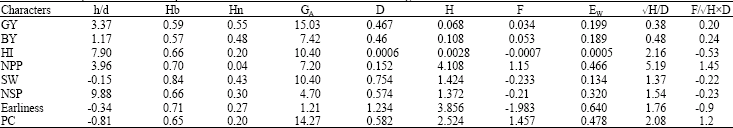 | |
Heritability estimates along with genetic advance are important selection parameters and normally more helpful in predicting the gain under selection than heritability estimates alone. However, heritability estimates are influenced by the type of genetic material, sample size, method of sampling, conduct of experiment, method of calculation and effect of linkage. Genetic advance which refers to the improvement in the mean genotypic value of selected individuals over the parental population is influenced by the genetic variability, heritability and selection intensity (Alza and Martinez, 1997; Sharma, 2003). Low narrow-sense heritability (less than 0.2) was shown for NPP, but HI and PC displayed moderate (0.2-0.5) heritability (Farshadfar, 1998; Stansfield, 2005). The rest of the traits indicated high heritability estimate (greater than 0.5) (Tefra and Peat, 1997). The low Hn in NPP suggested that environmental effects constitute a major portion of the total phenotype variation for this character (Farshadfar et al., 2008). The high heritability estimates for GY, BY, SW, NSP and earliness revealed that effective progress can be made through selection. All the characters showed high Hb.
The difference between Hb and Hn for HI, NPP and PC suggested the involvement of dominance effect in the genetic constitution of these characters which is in agreement with the rate of √H/D in Table 3 (>1). The results of this investigation are confirmed by the report of Omoigii et al. (2006) and Yücel et al. (2006). The rate of genetic advance is connected with heritability (Mather and Jinks, 1982). The genetic advances for GY and PC was moderate (14-40%) indicating the importance of direct selection for these traits and the significance of indirect selection for other traits with low genetic advance (less than 14%) through correlated response with characters having high heritability and genetic advance (Farshadfar et al., 2001; Sharma et al., 1991).
Degree of dominance and variance components: Information about the genetic components of variation helps the breeder in the selection of desirable parents for crossing programs and also in deciding a suitable breeding procedure for the genetic improvement of various quantitative traits (Singh and Narayanan, 1993). Degree of dominance and variance components are presented in Table 3. In Table 3 EW, D and H are environmental, additive and dominance components, respectively.
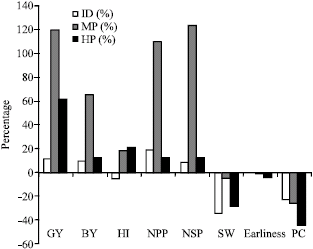 | |
| Fig. 1: | Percentage of heterosis and inbreeding depression for the characters investigated |
F is an indicator of correlation between D and H over all loci. If F is zero it means that dominant genes are in the parent with high performance, while negative F exhibits that dominant genes are in the low performance parent. If the ratio of F/√DxH is equal to or near one confirms that the magnitude and sign of dominance for all the genes monitoring the character is equal, therefore, the ratio √H/D is a good estimator of dominance. If F/√DxH is equal to zero or close to zero, the magnitude and sign of the genes controlling the character is not equal and hence √H/D explains average dominance. The h/d ratio estimates the degree of dominance (Kearsey and Pooni, 2004; Farshadfar, 1998; Sharma, 1998; Singh and Chaudhary, 1999; Farshadfar et al., 2001, 2008). The ratio of √H/D for GY, BY, HI, SW and NSP showed average dominance, while for NPP, earliness and PC displayed over dominance.
Heterosis and inbreeding depression: The estimates of heterosis and inbreeding depression together provide information about type of gene action involved in the expression of various quantitative traits. The percentage of heterosis with regard to High Parent (HP) and Mid-Parent (MP) and Inbreeding Depression (ID) (Fig. 1) exhibited that mid-parent and high parent heterosis were positive for GY, BY, HI, NPP and NSP, GY, NPP and NSP in chickpea while negative for SW, earliness and PC.
| Table 4: | Estimates of the genetic components of the mean for the characters studied |
 | |
| *, **: Significant at 0.05 and 0.01 level of probability, respectively, nsNon Significant | |
Sabaghpour et al. (2005) reported positive heterosis for GY, NPP and NSP in chickpea.
Inbreeding depression was positive for GY, BY, NPP and NSP, whereas, negative for HI, SW, earliness and PC. Positive ID revealed that the value of progenies in the F2 generation in comparison with F1 reduced, while negative ID indicated the increase of F2 in relation to F1 progenies. As GY showed positive heterosis, therefore, it is possible to utilize the phenomenon of heterosis for improvement of GY in chickpea.
Generation mean analysis: The joint scaling test (Mather and Jinks, 1982) was employed to estimate the mean (m), additive effect (d), dominance effect (h), additive x additive (i), additive x dominance (j) and dominance x dominance (l) values. The best dominance additive model was selected using non-significant Chi-square test (χ2) and the lower standard error (Table 4).
The genetic models fitted for GY and BY indicated additive and additivexdominance gene effects. Gowda et al. (2005) reported dominance type of gene action for GY in chickpea. No significant additive effect (d) was found for HI and proline content, indicating that selection is not effective in early generations. The epistatic effect (additive x dominance= [j]) was significant for GY, BY, NPP, NSP and PC, which is not fixable by selection under selfing conditions. Both additive x dominance [j] and dominance x dominance [l] effects were significant for NPP and NSP, supporting the presence of duplicate type of epistasis. This complementary interaction increases the variation between the generation and in the segregating population. The significance of [d] and [h] in the inheritance of NPP, SW and earliness revealed that both types of additive and dominance effects are involved in the genetics of No. of pod per plant and seed weight. Singh et al. (1993), Kidambi et al. (1990) and Girase and Deshmokh (2000) reported additive and non-additive (dominance and epistasis) effects in the inheritance of grain yield in chickpea. Based on the investigation of Tyagi and Srivastava (2001) in chickpea, biological yield is controlled by non-allelic interaction. Singh et al. (1993), Kidambi et al. (1990) and Girase and Desmokh (2000) reported the effect of additive-dominance and epistatic in the genetics of NPP, NSP and earliness.
Epistatic components related to a large number of interacting genes, lead to inefficient selection. But if the number of interacting genes will be reduced, selection will be efficient relied on the magnitude due to dominance and environmental variances. These results indicated epistasis could not be ignored in the genetic models (Farshadfar et al., 2008; Sood et al., 2006). Genotype-environment interactions may mask the expression of epistatic effects (Tefra and Peat, 1997), therefore a series of experiments are required for efficient breeding procedures (Frahsdfar et al., 2008). Since several important characters are influenced by dominance and non-allelic gene interaction, it is advisable to delay selection to later generation with increased homozygosity (Tefra an Peat, 1997).
REFERENCES
- Alza, J.O. and J.M.F. Martinez, 1997. Genetic analysis of yield and related traits in sunflower (Helianthus annus L.) in dryland and irrigated environments. Euphytica, 95: 243-251.
CrossRef - Bakheit, B.R., M.Z. El-Hifny, M.M. Eissa and S.B. Ragheb, 2002. Triple test cross and six population techniques for partitioning the components of genetic variance in faba bean (Vicia faba). J. Agric. Sci., 139: 61-66.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Bhatnagar, S., F.J. Betran and L.W. Rooney, 2004. Combining ability of quality protein maize inbreds Crop Sci., 44: 1997-2005.
Direct Link - Ehdaie, B., A.E. Hall, G.D. Farquhar, H.T. Nguyen and J.G. Waines, 1991. Water use efficiency and carbon isotope discrimination in wheat. Crop Sci., 31: 1282-1288.
Direct Link - Ehdaie, B. and G.G. Waines, 1993. Variation in water-use efficincency and its components in wheat. I. Well-watered pot experiment. Crop Sci., 33: 294-299.
Direct Link - Farshadfar, E., M. Ghandha, M. Zahravi and J. Sutka, 2001. Generation mean analysis of drought tolerance in wheat (Triticum aestivum L.). Acta Agron. Hung, 49: 59-66.
Direct Link - Farshadfar, E., R. Mohammadi, M. Aghaee and J. Sutka, 2003. Identification of QTLs involved in physiological and agronomic indicators of drought tolerance in Rye using multiple selection index. Acta Agron. Hung., 51: 419-428.
CrossRefDirect Link - Farshadfar, E., M. Aghaie, M. Sharifi and A. Yaghotipoor, 2008. Assessment of salt tolerance inheritance in barley via generation mean analysis. J. Biol. Sci., 8: 461-465.
CrossRefDirect Link - Farshadfar, E., S. Mahjouri and M. Aghaee, 2008. Detection of epistasis and estimation of additive and dominant components of genetic variation for drought tolerance in Durum wheat. J. Biol. Sci., 8: 598-603.
CrossRefDirect Link - Gamil, K.H. and Y.A. Saheal, 1986. Estimation of genetic effects for agronomic traits in wheat. Wheat Inform. Service, 62: 36-41.
Direct Link - Gowda, C.L.L., S. Ramesh, S. Chandra and H.D. Upadhyaya, 2005. Genetic basis of pod borer(Helicoverpa armigera) resistance and grain yield in desi and Kuboli chickpea (Cicer arietinum L.) under unprotected conditions. Euphytica, 145: 199-214.
CrossRef - Guler, M., M.S. Adak and H. Ulukan, 2001. Determining relationships among yield and some yield components using path coefficient analysis in chickpea (Cicer arietinum L.). Eur. J. Agron., 14: 161-166.
CrossRefDirect Link - Iqbal, M., A. Navabi, D.F. Salmon, R.C. Young and B.M. Mardoch et al., 2007. Genetic analysis of flowering and maturity time in high latitude spring wheat. Euphytica, 154: 207-218.
CrossRef - Jinks, J.L., J.M. Perkins and E.L. Breese, 1969. A general method of detecting additive, dominance and epistatic variation. II. Application to inbred lines. Heredity, 24: 45-47.
CrossRef - Johnson, H.W., H.F. Robinson and R.E. Comstock, 1955. Estimates of genetic and environmental variability in soybeans. Agron. J., 47: 314-318.
CrossRefDirect Link - Omoigui, L.O., M.F. Ishiyaku, A.Y. Kamara, S.O. Alabi and S.G. Mahammed, 2006. Genetic variability and heritability studies of some reproductive traits in cowpea (Vigna unguiculate (L.) Walp.). Afr. J. Biotechnol., 5: 1191-1195.
Direct Link - Rubio, J., F. Flores, M.T. Moreno, J.I. Cubero and J. Gill, 2004. Effects of the erect/bushy habit, single/double pod and late/early flowering genes on yield and seed size and their stability in chickpea. Field Crop Res., 90: 255-262.
Direct Link - Rebetzke, G.J., R.A. Richards, A.G. Condon and G.D. Farquhar, 2006. Inheritance of carbon isotope discrimination in bread wheat (Triticum aestivum L.). Euphytica, 150: 97-106.
CrossRef - Sabaghpour, S.H., R.S. Malhotra and T. Banai, 2005. Registration of hashem kaboli chickpea. Crop Sci., 45: 2651-2651.
Direct Link - Sharma, S.N., R.S. Sain and R.K. Sharma, 2003. Genetics of spike length in durum wheat. Euphytica, 130: 155-161.
CrossRef - Singh, R.K., B.B. Singh and P.D. Singh, 1993. Analysis of gene effects for yield and certain yield traits in chickpea. Indian J. Genet., 53: 203-207.
Direct Link - Sood, S., N.R. Kalia, S. Bhateria and S. Kumar, 2006. Detection of genetic components of variation for some biometrical traits in Linum usitatssimum L. in sub-montain Himalayan region. Euphytica, 155: 107-115.
CrossRef - Tefra, H. and W.E. Peat, 1997. Genetics of grain yield and other agronomic characters in (Eragrostis tef Zucctrotter ). II the triple test cross. Euphytica, 96: 193-202.
Direct Link - Umarahan, P., R.P. Ariyanayagam and S.Q. Haque, 1997. Genetic analysis of yield and its componenets in vegetable cowpea (Vigna unguiculata L. walp). Euphytica 96: 207-213.
CrossRef - Viana, J.M.S., 2005. Dominance, epistasis, heritability and expected genetic gains. Genet. Mol. Biol., 28: 67-74.
Direct Link - Yaghotipoor, A. and E. Farshadfar, 2007. Non-parametric estimation and component analysis of phenotypic stability in chickpea (Cicer arietinum L.). Pak. J. Biol. Sci., 10: 2646-2652.
CrossRefPubMedDirect Link - Yocel, D.O., A.E. Anlarsal and C. Yocel, 2006. Genetic variability, correlation and path analysis of yield and yield components in chickpea (Cicer arietinum L.). Turk. J. Agric. For., 30: 183-188.
Direct Link