
H. Boubaker
Laboratoire de Biotechnologie et de Valorisation des Ressources Naturelles, Faculte des sciences, B.P. 8106, Universite Ibn Zohr, Agadir, Maroc
B. Saadi
Laboratoire de Biotechnologie et de Valorisation des Ressources Naturelles, Faculte des sciences, B.P. 8106, Universite Ibn Zohr, Agadir, Maroc
E.H. Boudyach
Laboratoire de Biotechnologie et de Valorisation des Ressources Naturelles, Faculte des sciences, B.P. 8106, Universite Ibn Zohr, Agadir, Maroc
A. Ait Benaoumar
Laboratoire de Biotechnologie et de Valorisation des Ressources Naturelles, Faculte des sciences, B.P. 8106, Universite Ibn Zohr, Agadir, Maroc
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 21 | Page No.: 3903-3909







ABSTRACT
In response to growers report of reduced efficacy of benzimidazole fungicides for control of cigar end-rot, a survey was conducted from 2002 to 2005 in four banana-growing locations, with various histories of benzimidazole use, in the Souss-Massa-Drâa Valley, Agadir to determine the proportion of isolates that were in vitro resistant to benomyl and thiophanate-methyl. Of the 274 isolates, collected in Biougra, Belfâa and Ouled-teima locations with more than 10 years of benzimidazole use, 65% (180/274) were resistant to benomyl, 67% (184/274) were resistant to thiophanate-methyl and 65% were resistant to both fungicides tested at a discriminatory concentration of 10 μg mL-1. Only 1.5% of isolates exhibited a differential reaction to the two fungicides. No resistance to benomyl and thiophanate-methyl were detected in isolates collected from banana plantations in the Tamri location which has no known history of benzimidazole use. The mean effective concentrations that reduced growth by 50% (EC50) for resistant-isolates of V. theobromae were between 80 and 97 μg mL-1 for benomyl and between 194 to 233 μg mL-1 for thiophanate-methyl. In contrast, wild-type isolates exhibited mean EC50 values for benomyl and thiophanate-methyl of 0.47 and 0.91 μg mL-1, respectively. All 305 isolates from the four locations sampled in this study were sensitive in vitro to chlorothalonil at 10 μg mL-1. Conidial germination of sensitive-isolates collected from banana plantations never exposed to benzimidazole fungicides were completely inhibited by 1000 μg mL-1 of benomyl or thiophanate-methyl. However, conidial germination of resistant-isolates was not affected by both fungicides tested at 1000 μg mL-1.
PDF Abstract XML References Citation
How to cite this article
H. Boubaker, B. Saadi, E.H. Boudyach and A. Ait Benaoumar, 2008. Resistance of Verticillium theobromae to Benzimidazole Fungicides in Morocco. Journal of Applied Sciences, 8: 3903-3909.
DOI: 10.3923/jas.2008.3903.3909
URL: https://scialert.net/abstract/?doi=jas.2008.3903.3909
DOI: 10.3923/jas.2008.3903.3909
URL: https://scialert.net/abstract/?doi=jas.2008.3903.3909
INTRODUCTION
In Morocco, the greenhouse culture of bananas (Musa spp.) began in the 1980s and soon became one of the fastest-growing segments of agriculture, especially after a government-imposed ban on foreign banana imports went into effect in 1978. Currently, Morocco and Spain are the world`s largest greenhouse banana-producing countries (Galan Sauco et al., 2004). In 2006, banana cultivated area in Morocco spanned some 4400 ha, of which more than 98% was represented by plastic greenhouse plantations, with the Souss-Massa-Drâa (SMD) Valley as the main area of production (60 to 70% of total production).
In Morocco, the culture of bananas in plastic greenhouses is being hindered by nematodes and fungal pathogens, as the main causes of disease of this crop (Janick and Ait-Oubahou, 1989; Guedira et al., 2004). Among fungal diseases, cigar-end rot is one of the most serious infections in greenhouses banana cultures. This disease is caused by two fungi, Verticillium theobromae (Turc.) Mason and Hughes and Trachysphaera fructigena Tabor and Bunting. T. fructigena, is known to cause a destructive rot in the banana plantations of West and Central Africa (Snowdon, 1990), whereas, V. theobromae is more widespread, occurring in most banana-growing regions (Bhangale and Patil, 1983; Janick and Ait-Oubahou, 1989). Unlike others soil-borne Verticillium species that colonize the vascular tissues of plants, V. theobromae is mainly a banana fruit-rotting fungus. This fungus is also a member of a pathogenic complex which causes a rotting of the crowns during shipment and transit of boxed bananas (Snowdon, 1990; Alvindia et al., 2006).
In the SMD Valley, cigar-end rot is controlled primarily by multiple applications of benomyl and/or thiophanate-methyl, because non chemical alternatives, such as the manual removal of the pistil and perianth, do not represent commercially-viable means of control. Both benomyl and thiophanate-methyl belong to the benzimidazole class of fungicides known as a multiplication inhibitor during fungal mitosis that share a similar site specific mode of action (Ma and Michailides, 2005). Most growers in the SMD manage the disease by spraying systemic fungicides (e.g., benomyl, thiophanate-methyl) or combinations of systemic and protectant fungicides (e.g., chlorothalonil, mancozeb). The fungicide regime commonly used includes two to eight benzimidazole sprays per season of either benomyl or thiophanate-methyl at rates of 0.25 g to 1.5 g a.i. L-1 and it is not uncommon for a grower to use the same fungicide for the entire production cycle. In recent years unsatisfactory control of cigar-end rot has been observed in some banana greenhouses, with a previous history of benzimidazole fungicides use. The growers commonly attribute this unsatisfactory control to application method, or frequency and rate of application. However, this failure to achieve control of cigar-end rot in banana greenhouses treated with benomyl and/or thiophanate-methyl may be the result of acquired resistance to benzimidazole among populations of V. theobromae. Several reports (Hewitt, 1998; Staub, 1991) indicated that when populations of fungal pathogens are repeatedly exposed to site-specific fungicides, resistant strains can be readily selected. The result of this selection has been widely documented in cases in which benzimidazole fungicides were used extensively in controlling fungal pathogens during field as well as greenhouse production (Johnson et al., 1994; Murray, 1996; Hanson et al., 1996; van de Graaf et al., 2003). However, little is know about the benzimidazole fungicides sensitivity of V. theobromae populations in banana greenhouses in Morocco.
Therefore, the objectives of this study were to determine if benzimidazole fungicides resistance exists in SMD banana greenhouses and to determine the levels of sensitivity of V. theobromae isolates collected from four banana-growing locations of the SMD Valley, with various histories of benzimidazole use, to both benomyl and thiophanate-methyl.
MATERIALS AND METHODS
Sampling locations: Samples of banana fruits exhibiting cigar-end rot symptoms were collected from four geographically-isolated locations in SMD Valley, Agadir between 2002 and 2005. Benzimidazole fungicides use varied among sampling locations, with three of the locations, namely, Biougra, Belfâa and Ouled-teima totalling more than 10 years of benzimidazole fungicides use and representing the main commercial banana plastic greenhouse production areas. The fourth location, Tamri, is isolated from the above-mentioned areas and consists of open field banana plantations, with no history of benzimidazole use. This fourth location was used to collect baseline (wild-type) isolates of V. theobromae. A representative sample of at least 30 fruits was collected from each location. Fruits from individual bunches were picked, placed in separate polyethylene bags and transported to the laboratory.
Pathogen isolation: Infected fruits were surface disinfected with a solution of 0.5% sodium hypochlorite (NaClO), rinsed with sterile water and then allowed to air dry. Small pieces of fruit tissue were aseptically excised from the advancing edge of the rot and placed on Water-Agar (WA, 2%) containing 50 μg mL-1 of rifampicin. Cultures were incubated at 23°C for 2 to 5 days. When fungal growth from the tissue became visible, the fungi were subcultured on PDA and reincubated as above. Identification of V. theobromae was verified by examination under a compound microscope. A total of 305 V. theobromae isolates were collected from fruits sampled at the four studied locations, with each isolate originating from a different fruit. All isolates were single-spored and maintained on PDA at 5°C.
Fungicides: The fungicides used in this study were benomyl (50%, Benlate 50WP, AMAROC, Maroc), thiophanate-methyl (70%, Pelt 44WP, BAYER, Maroc) and chlorothalonil (75%, Daconil 75WP, CPCM, Maroc). The chlorothalonil was included in this study for comparison. Fungicide solutions were first prepared by dissolving each commercially-formulated fungicide in acetone. An aqueous suspension of each fungicide was then prepared and added aseptically to molten (50°C) sterile Potato Dextrose Agar (PDA). Control dishes contained PDA and acetone and the final concentration of acetone in the medium did not exceed 1% (v/v). All concentrations were expressed as active ingredient (a.i.).
Fungicide sensitivity tests: The isolates of V. theobromae were in vitro-tested for sensitivity to benomyl, thiophanate-methyl and chlorothalonil at a discriminatory concentration of 10 μg mL-1 of medium. Agar plugs (5 mm diameter) were cut from the periphery of actively growing colonies and transferred, mycelium down, to three replicate Petri dishes containing PDA medium supplemented with fungicide. After a ten days incubation period at 23°C, the isolates were considered either resistant if growth was observed on fungicide-amended medium or sensitive if no growth has occurred.
Determination of fungicide sensitivity (concentration producing 50% growth inhibition EC50 values): For each location, ten V. theobromae isolates were used to determine the fungicide concentration producing about a 50% growth inhibition (EC50). These isolates were arbitrary chosen and represent different locations of bananas production (Biougra, Belfâa, Ouled-teima and Tamri).
A 5 mm diameter disk was taken from the margin of a seven day-old culture of each isolate of V. theobromae and placed, upside down, at the center of a PDA dish. The PDA medium was amended with the following concentration of benomyl or thiophante-methyl: 0, 0.1, 1, 10, 100 and 1,000 μg mL-1 for the benzimidazole-resistant isolates, collected from Biougra, Belfâa and Ouled-Teima locations and 0, 0.01, 0.1, 0.5, 1 and 5 μg mL-1 for the benzimidazole-sensitive isolates of V. theobromae collected from Tamri location. Three Petri dishes were used for each fungicide concentration. Colony diameters were measured after seven days at 25°C. Percent growth inhibition at each fungicide concentration was calculated according to the following formula:
Growth inhibition (%) = [(unamendedfungicideamended)/unamended] x100 |
EC50 values were calculated for each isolate and each fungicide by regression analysis of the percent inhibition of fungal growth versus the log of the fungicide concentration (μg mL-1). The experiment was performed twice.
Effect of fungicides on conidial germination: The inhibitory effect of fungicides on spore germination was tested on 2% Water-Agar (WA) amended with 0, 100 and 1,000 μg mL-1 of benomyl, thiophanate-methyl or chlorothalonil. A spore-suspension from a mixture of either four resistant-isolates (Biougra, Belfâa and O-Teima locations) or four sensitive-isolates (Tamri location) was obtained from one-week-old cultures by flooding the cultures with sterile distilled water containing 0.05% (v/v) Tween 80 and filtered through two layers of sterile cheesecloth to remove hyphal fragments. The spore concentration of these suspensions was adjusted to 1x105 conidia mL-1 with the aid of a haemocytometer. Aliquots (100 μL) of spore suspensions were aseptically-spread in triplicate onto WA dishes. After a 20 h incubation at 23°C, germination was determined by observing at least 100 conidia for each concentration under a light microscope. A spore was scored as germinated if the germ tube length was equal to at least one time that of the conidium. Each treatment was replicated twice.
Statistical analysis: The data were subjected to statistical Analysis of Variance (ANOVA) using the STATISTICA software, version 6, StatSoft, 2001, France. Mean separation was performed following the method of Newman and Keuls test at p = 0.05.
RESULTS
Fungicide sensitivity tests: In this study, we collected 95, 88, 91 and 31 V. theobromae isolates from banana plantations located in Biougra, belfâa, Ouled-teima and Tamri, respectively.
| Table 1: | Sensitivity of Verticillium theobromae isolates to benomyl, thiophanate-methyl and chlorothalonil collected from four locations in the Souss-Massa-Drâa Valley, Morocco |
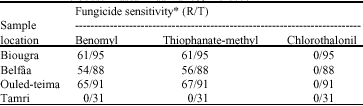 | |
R: No. of isolates resistant to fungicide, T: No. of isolates tested, *Sensitivity of isolates to fungicide was tested on PDA medium amended with 10 μg mL-1. Isolates which failed to grow on the amended medium were considered sensitive | |
All isolates were tested in vitro for resistance to either benomyl or thiophanate-methyl at a discriminatory concentration of 10 μg mL-1. Results of Table 1 show that 64, 61 and 71% of V. theobromae isolates from Biougra, Belfâa and Ouled-teima locations, respectively, were resistant to benomyl, whereas isolates from the Tamri location were sensitive to benomyl. The percentage of thiophanate-methyl resistant isolates from Biougra, Belfâa and Ouled-teima locations ranged from 64 to 74%. Whereas all 31 field-collected V. theobromae isolates from the Tamri location were sensitive to thiophanate-methyl tested at a discriminatory concentration of 10 μg mL-1 (Table 1). It has often been observed in other plant-pathogenic fungi that isolates resistant to one benzimidazole fungicide showed reduced sensitivity to the other benzimidazole fungicides (Keinath and Zitter, 1998). In this study, the majority of V. theobromae isolates exhibited cross-resistance to both fungicides. Of 305 single-conidium isolates tested, 59% were resistant to both benomyl and thiophanate-methyl, while 40% of these isolates were sensitive to both compounds. Only, four V. theobromae isolates exhibited a differential sensitivity to the two fungicides: resistance to thiophanate-methyl but sensitivity to benomyl. We also found that all 305 V. theobromae isolates were sensitive to the chlorothalonil when tested at a discriminatory concentration of 10 μg mL-1 (Table 1).
Level of resistance: As shown in Table 2, the mean EC50 values of 10 benzimidazole-sensitive isolates of V. theobromae collected from the Tamri location on benomyl-or thiophanate-methyl amended PDA were 0.47 and 0.91 μg mL-1, respectively. These values were significantly lower than those observed for isolates from the other locations. The Tamri population was therefore considered as being indicative of a wild-type population (Table 2). The mean EC50 values of V. theobromae isolates sampled from commercial banana greenhouses at Biougra (97 μg mL-1), Belfâa (90 μg mL-1) and Ouled-teima (80 μg mL-1) to benomyl were not significantly different (p = 0.05).
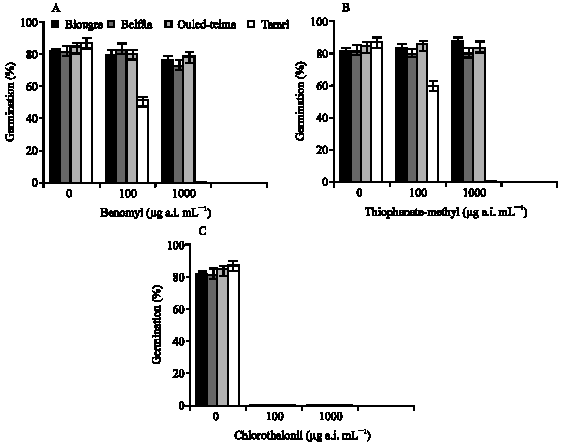 | |
| Fig. 1: | Effect in vitro of fungicides on conidial germination of V. theobromae isolates collected from Biougra, Belfâa, Ouled-teima and Tamri locations. The fungicides tested were benomyl (A), thiophanate-methyl (B) and chlorothalonil (C) at the following concentrations: 0, 100 and 1,000 μg mL-1. Vertical bars represent standard deviations of the mean |
| Table 2: | Sensitivity (EC50) to benzimidazole fungicides of Verticillium theobromae isolates collected in banana plantations at four locations in Souss-Massa-Drâa Valley, Morocco |
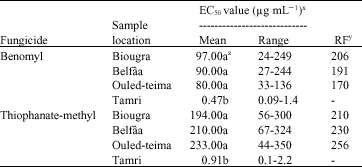 | |
xEC50: Effective concentration of fungicide in the culture medium needed to inhibit 50% of mycelial growth, yResistance Factor (RF) = mean EC50 resistant-isolates/mean EC50 sensitive-isolates, zValues within columns followed by the same letter are not significantly different (p = 0.05). Mean values were determined for 10 isolates per sampling location | |
For thiophanate-methyl, the mean EC50 values for isolates collected from the three locations ranged between 194 and 233 μg mL-1 and no significant difference was apparent between these locations (Table 2). Compared to benomyl sensitivity, isolates of V. theobromae collected in the same banana grown locations were less sensitive to thiophanate-methyl than to benomyl. Resistance factors for both fungicides were higher and ranged between 170 and 206 for benomyl and from 210 and 256 for thiophanate-methyl, indicating the presence of V. theobromae isolates that were highly insensitive to both fungicides.
Effect of fungicides on conidial germination: A spore-suspension mixture of four resistant-isolates to benomyl or thiophanate-methyl collected from Biougra, Belfâa and Ouled-Teima and four sensitive-isolates collected in Tamri, were used to analyze the effects of fungicides on conidial germination of V. theobromae. The results obtained (Fig. 1B, C) showed that benomyl and thiophanate-methyl did not prevent spore germination of isolates resistant to both fungicides, even at the highest concentration tested (1000 μg mL-1). In contrast, conidial germination of V. theobromae isolates sensitive to benomyl and thiophanate-methyl was totally inhibited at 1000 μg mL-1. For both fungicides, conidium germination rates for sensitive isolates were only slightly decreased in the presence of 100 μg mL-1 as compared to resistant isolates and were above 50%. The chlorothalonil at 100 μg mL-1 completely inhibited spore germination of both benzimidazole-resistant and benzimidazole-sensitive isolates of V. theobromae collected from the four locations studied (Fig. 1A).
DISCUSSION
The benzimidazole fungicides have been used quite extensively during the last 20 years to control cigar-end rot in the SMD Valley, Agadir, Morocco. Benzimidazole are systemic fungicides that acts as a multiplication inhibitor during fungal mitosis (Ma and Michailides, 2005) and resistance to these compounds has been detected in many fungal species. The present study showed that all V. theobromae wild-type isolates collected from an open-field banana plantation in the Tamri region were sensitive, in vitro, to both benomyl and thiophanate-methyl tested at the discriminatory concentration of 10 μg mL-1. These results suggest that naturally occurring benzimidazole-resistant strains of V. theobromae are absent or present at very low frequency from locations without any prior history of benzimidazole exposure. However, among 274 V. theobromae isolates collected from banana locations with a history of prolonged benzimidazole use, 65% (180/274) and 67% (184/274) were resistant to benomyl and thiophanate-methyl, respectively. Considering the extensive and heavy use of both fungicides in commercial banana greenhouses, it is not surprising that resistance to benomyl and thiophanate-methyl was so prevalent in V. theobromae populations sampled from SMD, Valley. The high incidence of resistance to both fungicides was similar to that reported for others fungal pathogens (Malathrakis and Vakalounakis, 1983; Romero and Sutton, 1998; Keinath and Zitter, 1998).
It has been observed in other plant-pathogenic fungi that isolates resistant to one benzimidazole also exhibit reduced sensitivity to the other benzimidazole fungicide (Bus et al., 1991; Kawchuk et al., 1994). In this study, we found that all isolates resistant to benomyl were also resistant to thiophanate-methyl. Present results indicate that, in all likelihood, the unsatisfactory control of cigar-end rot in the SMD, Valley banana greenhouses stems from the emergence and the spread of V. theobromae strains highly resistant to benzimidazole fungicides. This is in agreement with previous reports indicating that decreased sensitivity of fungal pathogens to benzimidazole is directly correlated with reduced performance of the fungicides in the field and in the greenhouse (Moorman and Lease, 1992; De Lapeyre et al., 1997; Romero and Sutton, 1998; Errampalli et al., 2001).
The majority of V. theobromae resistant-isolates in our collection exhibited high levels of resistance to both thiophanate-methyl and benomyl. The average EC50 of isolates resistant to benomyl was between 80 and 97 μg mL-1 and between 194 and 233 μg mL-1 for thiophanate-methyl. The median EC50 values for benzimidazole fungicides tested did not differ significantly (at p = 0.05) among locations with previous history of benzimidazole use. In contrast, the range of EC50 values for V. theobromae sensitive-isolates was between 0.09 and 1.4 μg mL-1 for benomyl and between 0.1 and 2.2 μg mL-1 for thiophanate-methyl. The sensitivity of V. theobromae isolates, collected from the Tamri location, to benomyl was of the same order as that reported by Igeleke and Ayanru (2007). In another study, among eight isolates of V. theobromae isolated from rotted banana crowns, three were less sensitive to the benzimidazole fungicide thiabendazole, with EC50 values higher than 10 μg mL-1 (Johanson and Blazquez, 1992). The EC50 values for benomyl and thiophanate-methyl were consistent with other reports. For example, benomyl and thiophanate-methyl EC50 ranged from 27 to 251 μg mL-1 and from 27 to more than 1,000 μg mL-1, respectively, in isolates of Botrytis cinerea (LaMondia and Douglas, 1997). The different levels of resistance to benomyl among V. theobromae populations may be due to the presence of a different allele for benomyl resistance as observed with Venturia inaequalis populations (Koenraadt et al., 1992).
The resistance factors, calculated by dividing mean EC50 values of resistant-isolates collected in Biougra, Belfâa and Ouled-Teima by the mean EC50 values of sensitive-isolates collected in Tamri location, were between 170 and 206 for benomyl and between 210 and 256 for thiophanate-methyl. The difference in resistance factors reflects greater variation in sensitivity to benomyl and thiophanate-methyl within populations of V. theobromae and indicates differences in sensitivity to benzimidazole fungicides between the wild-type population and the exposed populations of V. theobromae.
With regard to the effect of fungicides on conidial germination, the data presented here show that the benomyl and the thiophanate-methyl tested at a concentration of 1000 μg mL-1 were without effect on conidial germination of V. theobromae resistant-isolates collected from locations with a history of benzimidazole use. However, these two fungicides totally inhibited conidial germination of sensitive-isolates at a concentration of 1000 μg mL-1.
In the present study, at 10 and 100 μg mL-1, the multi-site fungicide chlorothalonil completely inhibited mycelial growth and germination of conidia, respectively, of both benzimidazole-sensitive and benzimidazole-resistant isolates of V. theobromae, collected from the four locations sampled. Therefore, an appropriate strategy for limiting the spread of V. theobromae resistance to benomyl and thiophanate-methyl in the SMD Valley banana greenhouses would involve the use of mixtures of benzimidazole fungicides and contact fungicides such as chlorothalonil. Fungicide insensitivity is generally not a concern with protectant fungicides such as chlorothalonil because of their multi-site mode of action (Vincent and Sisler, 1968).
In conclusion, the data presented in this study reveal, to our knowledge for the first time, that a majority of V. theobromae isolates collected in commercial banana-greenhouses in SMD Valley between 2003 and 2005 were resistant to benomyl and thiophanate-methyl. This high proportion of isolates with reduced sensitivity to benzimidazole fungicides may account for the unsatisfactory control of cigar-end disease in the SMD. On the basis of these data, the use of benzimidazole fungicides for the control of cigar-end rot should be seriously questioned. Moreover, these results emphasize the need for new control strategies involving lower pesticide use, which would be in line with consumer requirements.
REFERENCES
- Alvindia, D.G., T. Kobayashi and K.T. Natsuaki, 2006. The aerial and fruit surface populations of fungi in nonchemical banana production in the Philippines. J. General Plant Pathol., 72: 257-260.
CrossRef - Bus, V.G., A.J. Bongers and L. Risse, 1991. Occurrence of Penicillium digitatum and P. italicum resistant to benomyl, thiabendazole and imazalil on citrus from different geographic origins. Plant Dis., 75: 1098-1100.
Direct Link - De Lapeyre, L. de Bellaire and C. Dubois, 1997. Distribution of thiabendazole-resistant Colletitrichum musae isolates from Guadeloupe banana plantations. Plant Dis., 81: 1378-1383.
CrossRefDirect Link - Errampalli, D., J.M. Saunders and J.D. Holley, 2001. Emergence of silver scurf (Helminthosporium solani) as an economically important disease of potato. Plant Pathol., 50: 141-153.
CrossRef - Guedira, A., A. Rammah, Z. A. Triqui, H. Chlyah, B.Chlyah and R. Haïcour, 2004. Evaluation of the resistance to two nematodes: Radopholus similes and Meloidogyne spp. in four banana genotypes in Morocco. C. R. Biologies, 327: 745-751.
PubMedDirect Link - Igeleke, C.L. and D.K.G. Ayanru, 2007. Evaluation of fungicides on growth and conidial germination of Verticillium theobromae isolated from plantain. J. Applied Sci., 7: 531-535.
CrossRefDirect Link - Johanson, A. and B. Blazquez, 1992. Fungi associated with banana crown rot on field-packed fruit from the Windward Islands and assessment of their sensitivity to the fungicides thiabendazole, prochloraz and imazalil. Crop Prot., 11: 79-83.
CrossRef - Johnson, K.B., T.L. Sawyer and M.L. Powelson, 1994. Frequency of benzimidazole- and dicarboximide-resistant strains of Botrytis cinerea in Western Oregon small fruit and snap bean plantings. Plant Dis., 78: 572-577.
CrossRefDirect Link - Kawchuk, L.M., J.D. Holley, D.R. Lynch and R.M. Clear, 1994. Resistance to thiabendazole and thiophanate-methyl in canadian isolates of Fusarium sambucinum and Helminthosporium solani. Am. Potato J., 71: 185-192.
CrossRef - Keinath, A.P. and T.A. Zitter, 1998. Resistance to benomyl and thiophanate-methyl in Didymella bryoniae from South Carolina and New York. Plant Dis., 82: 479-484.
CrossRefDirect Link - LaMondia, J.A. and S.M. Douglas, 1997. Sensitivity of Botrytis cinerea from connecticut greenhouses to benzimidazole and dicarboximide fungicides. Plant Dis., 81: 729-732.
CrossRefDirect Link - Ma, Z. and T.J. Michailides, 2005. Advances in understanding molecular mechanisms of fungicide resistance and molecular detection of resistant genotypes in phytopathogenic fungi. Crop Prot., 24: 853-863.
CrossRef - Malathrakis, N.E. and D.J. Vakalounakis, 1983. Resistance to benzimidazole fungicides in the gummy stem blight pathogen Didymella bryoniae on cucurbits. Plant Pathol., 32: 395-399.
CrossRefDirect Link - Moorman, G.W. and R.J. Lease, 1992. Benzimidazole- and dicarboximide-resistant Botrytis cinerea from pennsylvania greenhouses. Plant Dis., 76: 477-480.
CrossRef - Murray, T.D., 1996. Resistance to benzimidazole fungicides in the cereal eyespot pathogen, Pseudocercosporella herpotrichoides, in the Pacific Northwest 1984 to 1990. Plant Dis., 80: 19-23.
CrossRefDirect Link - Romero, R.A. and T.B. Sutton, 1998. Characterization of benomyl resistance in Mycosphaerella fijiensis, cause of black Sigatoka of banana, in Costa Rica. Plant Dis., 82: 931-934.
CrossRefDirect Link - Staub, T., 1991. Fungicide resistance: Practical experience with antiresistance strategies and the role of integrated use. Annu. Rev. Phytopathol., 29: 421-442.
CrossRefDirect Link - Van de Graaf, P., T.M. ONeill, J.M. Chartier-Hollis and M.E. Joseph, 2003. Aspects of the biology and control of benzimidazole resistant isolates of Phoma clematidina, cause of leaf spot and wilt in clematis. J. Phytopathol., 151: 442-450.
CrossRefDirect Link - Vincent, P.G. and H.D. Sisler, 1968. Mechanism of antifungal action of 2,4,5,6-tetrachloroisophthalonitrile. Physiol. Plant., 21: 1249-1264.
CrossRefDirect Link