
M. Goudarzi
Department of Crop Production and Plant Breeding,
College of Agriculture, Shiraz University, Shiraz, Iran
H. Pakniyat
Department of Crop Production and Plant Breeding,
College of Agriculture, Shiraz University, Shiraz, Iran
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 12 | Page No.: 2300-2305







ABSTRACT
The effects of salt stress (NaCl and Na2SO4 in 1:1 ratio) on the activity of peroxidase (POD), proline accumulation, protein, Na + and K + contents and K + /Na + ratio were studied on leaves of two maize and wheat genotypes. Wheat cvs., [Kavir (tolerant) and Ghods (sensitive)] and maize cvs. [704 (tolerant) and 666 (semi-tolerant)] were grown under control (ECe = 1.26 dS m 1) and two levels of salinity (ECe = 6.8 and 13.8 dS m 1, respectively) in a greenhouse. A Completely Randomized Design (CRD) with factorial treatments with three replications was used. Salinity stress decreased K + and K + /Na + ratio and increased Na + , proline, protein and POD activity in both genotypes of wheat and maize under both salinity levels. Furthermore the results showed a lower amount of proline, protein and POD activity in wheat than maize cvs. On the other hand, the higher amounts of K + and K + /Na + ratios which were found in wheat than maize resulted in better ion homeostasis in wheat that caused this species to have a higher tolerance than maize. In wheat cvs.; salinity stress resulted in an increase in proline and protein contents, POD activity, K + and K + /Na + ratio in Kavir cultivar than Ghods. However, Ghods cultivar showed a higher Na + content than Kavir. In maize, cultivar 704 showed a higher increase in proline and protein contents, POD activity, K + and K + /Na + ratio and a lower increase in Na + than 666 cultivar. Maize cvs. showed a higher level of proline, protein and POD activity than wheat cvs., but these components may have been a reaction to salt stress in maize and not a plant response associated with tolerance. The results of this study suggest that Na + and K + contents and K + /Na + ratio in maize as a C4 plant and in wheat as a C3 plant may be considered for selecting the tolerant cultivars.
PDF Abstract XML References Citation
How to cite this article
M. Goudarzi and H. Pakniyat, 2008. Comparison Between Salt Tolerance of Various Cultivars of Wheat and Maize. Journal of Applied Sciences, 8: 2300-2305.
DOI: 10.3923/jas.2008.2300.2305
URL: https://scialert.net/abstract/?doi=jas.2008.2300.2305
DOI: 10.3923/jas.2008.2300.2305
URL: https://scialert.net/abstract/?doi=jas.2008.2300.2305
INTRODUCTION
Crops grown in arid and semi-arid regions are often exposed to adverse environmental factors such as drought or high soil salinity. Increased salinization of arable land is expected to have devastating global effects, resulting in 30% land loss within next 25 years and up to 50% by the middle of 21st century (Dionisio-Sese and Tobita, 1998). Plant salt tolerance has generally been studied in relation to regulatory mechanisms of ionic and osmotic homeostasis (Yeo, 1998; Zhu, 2003; Ashraf and Harris, 2004; Chinnusamy et al., 2005). In addition to ionic and osmotic components, salt stress, like other abiotic stresses, also leads to oxidative stress through an increase in Reactive Oxygen Species (ROS) (Mittler, 2002; Neill et al., 2002; Dionisio-Sese and Tobita, 1998). These ROS are highly reactive and can alter normal cellular metabolism through oxidative damage to lipids, proteins and nucleic acids (Alscher et al., 1997; Imlay, 2003). The ability of plants to tolerate salt is determined by multiple biochemical pathways that facilitate retention and/or acquisition of water, protect chloroplast functions and maintain ion homeostasis. Essential pathways include those that lead to synthesis of osmotically active metabolites, specific proteins and certain free radical scavenging enzymes that control ion and water flux and support scavenging of oxygen radicals or chaperones.
These cytotoxic oxygen species are highly reactive and in the absence of any protective mechanism, they can seriously disrupt normal metabolism through oxidative damage to lipids, protein and nucleic acids. Plants defend against the reactive oxygen (by-products of hyperosmotic oxygen species), by induction of activities of certain antioxidative enzymes such as catalase, peroxidase, glutathione reductase and superoxide dismutase, which scavenge reactive oxygen species. There are several reports of increasing in activity of antioxidative enzymes in wheat and more species plants under saline condition (Sairam et al., 2002; Mittler, 2002; Rios Gonzalez et al., 2002; Meneguzzo et al., 1999; Sairam et al., 2002; Hernandez et al., 1999; Manchandia et al., 1999).
It has been reported that soluble protein increases at salinity in many plants and decrease in some else (Parida et al., 2004; Noble and Rogers, 1992). Salinity also inhibits the synthesis of majority of shoot proteins (Agastian et al., 2000; Alamgir and Ali, 1999; Muthukumarasamy et al., 2000; Parida et al., 2004). A number of nitrogen-containing compounds (NCC) accumulated in plants exposed to saline stress (Wang et al., 2003). The most frequently accumulating NCC includes amino acids, amides, imino acids, proteins, quaternary ammonium compounds and polyamines (Parida et al., 2004). Many plants accumulate proline as a nontoxic and protective osmolyte under saline conditions (Lee and Liu, 1999; Khatkar and Kuhad, 2000; Muthukumarasamy et al., 2000; Singh et al., 2000; Jain et al., 2001). Under salt stress, plants maintain high concentrations of K + and low concentrations of Na + in the cytosol. They do this by regulating the expression and activity of K + and Na + transporters and of H + pumps that generate the driving force for transport (Zhu, 2003). There is a negative relationship between Na + and K + concentration in roots and leaves of many plants (Goudarzi and Pakniyat, 2008; Pakniyat and Armion, 2007). The selective uptake of K + as opposed to Na + is considered to be one of the important physiological mechanisms contributing to salt tolerance in many plant species (Poustini and Siosemardeh, 2004).
It is well documented that a greater degree of salt tolerance in plants is associated with a more efficient system for selective uptake of K + over Na + (Noble and Rogers, 1992; Pakniyat et al., 1997). Wheat and maize have C3 and C4 metabolism respectively. The objective of the present investigation is to study the effects of salinity on proline, protein, POD activity, leaf Na + and K + and K + /Na + ratio in both cultivars of wheat and maize and finally evaluating the changes of studied traits and suggesting the best criteria for selection in wheat (as a C3 plant) and maize (as a C4 plant) tolerant cultivars.
MATERIALS AND METHODS
Plant materials, culture and salt treatment application: In a two separate experiment that were conducted during 2006-2007, 10 seeds of two wheat cultivars (Kavir and Ghods) and maize (704 and 666) were sown in 5 kg pots in greenhouse at Agricultural College, Shiraz University, Iran. A Completely Randomized Design (CRD) with factorial treatments in three replications was used. After germination three plants were retained in each pot. Plants were subjected to three conditions: no salt (control) and two salinity levels using NaCl and Na2SO4 in 1:1 ratio. Salt stress treatments were applied to appropriate pots in split and in 4 stages within 4 weeks to final concentrations by irrigation based on soil field capacity and finally concentration of salinity expressed as ECe 1.26 (control) 6.8 and 13.8 dS m –1.
K + , Na + and K + /Na + ratio measurements: Leaves of plants were cut from soil surface and dried at 60°C oven for 48 h and made into fine powder using mortar and pestle. Samples (1 g) were ashed by putting them into crucibles and placed in 600°C electric furnace, for 4 h. Five milliliter of 2N HCl were added to ash samples and mixed with boiling distilled water and filtered by filter paper (Whatman No. 2) into 50 mL volumetric flasks. Na + and K + was measured using standard flame photometer and reported as mg g 1 dry weight.
Protein content: Protein content in the enzyme extracts was determined according to Bradford (1976). Using Bovine Serum Albumin as a standard.
Proline determination: Proline was determined according to the method described by Bates et al. (1973). Approximately 0.5 g of fresh leaf material was homogenized in 10 mL of 3% aqueous sulfosalicyclic acid and then this aqueous solution was filtered through Whatman’s No. 2 filter paper and finally 2 mL of filtrated solution was mixed with 2 mL acid-ninhydrin and 2 mL of glacial acetic acid in a test tube. The mixture was placed in a water bath for 1 h at 100°C. The reaction mixture was extracted with 4 mL toluene and the chromophore containing toluene was aspirated, cooled to room temperature and the absorbance was measured at 520 nm with a spectrophotometer. Appropriate proline standards were included for calculation of proline in the sample.
Peroxidase determination: Leaves (0.5 g) were homogenized in 50 mM sodium phosphate buffer (pH 7.0) containing 1% soluble polyvinyl pyrolidine (PVP). The homogenate was centrifuged at 13000 x g for 15 min at 4°C and the supernatant used for assays of the activities of peroxidase (POD). The activity of POD was assayed by adding aliquot of the tissue extract (100 µL) to 3 mL of assay solution, consisting of 3 mL of reaction mixture containing 13 mM guaiacol, 5 mM H2O2 and 50 mM Na-phosphate (pH 6.5) (Chance and Maehly, 1995). An increase of the optical density at 470 nm for 1 min at 25°C was recorded using spectrophotometer. POD activity was expressed as change in absorbance min 1 mg 1 protein. The increase in A470 was measured for 3 min and activity expressed as A470 mg 1 protein min 1.
Statistical analysis: Data were analyzed using MSTAT-C Software and the mean comparisons were made following Tukey’s Test at p = 0.05. Orthogonal comparisons between maize and wheat for studied traits were made, using SAS Software.
RESULTS
As the data analysis of variance shows, all the traits investigated in this experiment were significantly affected by the salinity (Table 1). The species employed also responded differently to the salinity stress (Table 4).
K + , Na + and K + /Na + ratio: The sodium concentration of all cultivars basically was low (Table 2, 3). With increase in salinity, leaf sodium content of all cultivars was increased. In wheat cultivars; Kavir (tolerant) showed the higher and lower K + and Na + , respectively compared to Ghods (sensitive), resulting in the higher K + /Na + ratio in this cultivar in both salinity levels (Table 2). The same response was found for the 704 hybrid (semi- tolerant) showed the higher K + /Na + and K + and lower Na + compared to 666 hybrid (Table 3). Orthogonal comparison between wheat and maize cvs. showed that K + and K + /Na + ratio had a higher level in wheat when compared with maize. On the other hand maize showed a higher Na + than wheat.
Protein content: The results confirmed that protein content was increased in exposure to salt stress. In wheat, protein content of tolerant cultivar was significantly higher than the sensitive one (Table 2). The same relationship was obtained for maize cultivar (Table 3). Furthermore maize showed a higher protein content than wheat cvs. when compared via orthogonal comparison.
| Table 1: | Mean square values for studied traits in wheat (W) and maize (M) |
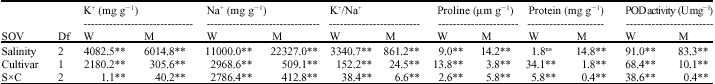 | |
| **: significantly at p< 0.01 ns: non significant | |
| Table 2: | Comparison between wheat cultivars (Kavir and Ghods) for studied traits under salt stress |
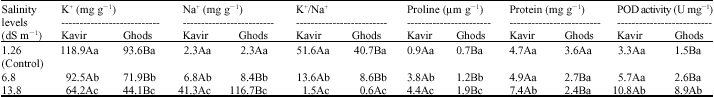 | |
In each column, means followed by the small same letter are not significant at p<0.05 with Tukey’s test. For every studied trait, means followed by the capital same letter, are not significant at p<0.05 with Tukey’s test between two cultivars | |
| Table 3: | Comparison between maize cultivars (704 and 666) for studied traits under salt stress |
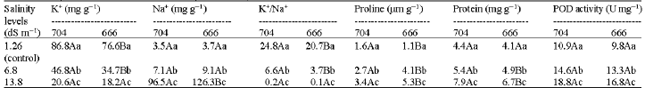 | |
In each column, means followed by the small same letter are not significant at p<0.05 with Tukey’s test. For every studied trait, means followed by the capital same letter, are not significant at p<0.05 with Tukey’s test between two cultivars | |
| Table 4: | Orthogonal comparison between wheat (W) and maize (M) species for studied traits under salt stress |
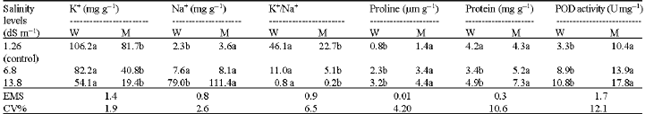 | |
For every studied trait, means followed by the same letter are not significant at corresponding salt levels | |
Proline content: Data on proline measurement indicated that proline accumulation has been the consequence of salinity and different salinity levels had significant effects (Table 1) on leaf proline content of two wheat and maize cultivar. Maize cvs. showed a higher proline content when compared with wheat cvs. Comparing proline content of cultivars it was found that in wheat, the Kavir cultivar accumulated higher proline content compared with Ghods, while in maize the 666 hybrid showed a higher proline concentration under saline conditions when compared with 704 hybrid (Table 2, 3).
Peroxidase activity: POD activity in leaves of all cultivar was remarkably increased as a result of salt stress. In salt stressed Kavir plants, leaf POD activity increased approximately 2.2 and 1.2 fold that of Ghods at 6.8 and 13.8 dS m 1, respectively (Table 2). In maize 704 cultivar showed a higher increase in POD activity than 666 and the activities were 1.09 and 1.2 fold in 6.8 and 13.8 dS m 1 respectively (Table 3). Orthogonal comparison between maize and wheat showed that maize had a higher POD activity in control and both salinity treatments when compared with wheat.
DISCUSSION
It has been demonstrated that both osmotic and ionic effects under salinity can limit photosynthesis and respiration leading to an increase in ROS generation, which are responsible for a secondary oxidative stress that can damage cellular structure and metabolism. It is also known that plant responses to salt stress are multigenic, involving both osmotic and ionic homeostasis, as well as cell detoxification, therefore; salt stress resistance may depend, particularly, on the enhancement of the antioxidative defense system, which includes antioxidant compounds and several antioxidative enzymes such as POD. In tolerant plant genotypes, POD activity was found to be higher, enabling plants to protect themselves against the oxidative stress, whereas such activity was not observed in sensitive plants (Sairam et al., 2002). Accumulation of proline and protein under stress in many plant species has been correlated with stress tolerance and its concentration has been shown to be generally higher in stress-tolerant than in stress-sensitive plants. Despite the presence of a strong correlation between stress tolerance and accumulation of proline and protein in higher plants, this relationship may not be universal (Fougere et al., 1991; Petrusa and Winicov, 1997; Lutts et al., 1999). However, further studies are needed to determine whether the relationship between stress tolerance and accumulation of proline is species-specific or if it can be altered by experimental conditions. Currently, there is more evidence supporting the presence of a positive relationship. Regarding proline content, we have constant argument that this component is a plant response component to stresses. Protein content was not considered in this discussion (De-Lacerda et al., 2003).
The high number of differentially regulated proteins under salt stress may reflect the physiological reactions to a combination of ion intolerance, osmotic adjustment and Na + /K + homeostasis (Liu and Zhu, 1997). It was evident from the present study that maize and wheat varied markedly in their sensitivity towards salt stress. Wheat was able to tolerate higher level of stress and could maintain the growth of its shoots at stress levels that were inhibitory for maize. As reported earlier, the type of osmolytes and capacity to accumulate them during stress may vary between different types of plants and specify their different adaptive mechanisms.
Maize showed inherently higher proline content than wheat during unstressed conditions and also accumulated significantly more proline than wheat during stress. The present study also indicated that wheat appeared to have advantage over maize during salt stress with respect to accumulation of proline and more accumulation of this component in maize than wheat under saline condition is not associated with tolerance and only was a reaction to salt stress thus wheat has a better capacity for osmoregulation. It is widely accepted that competition exists between Na + and K + leading to a reduced level of internal K + at high external NaCl concentration (Botella et al., 1997). This can result in high Na + /K + ratios that reduce plant growth and eventually become toxic (Schachtman and Lio, 1999).
A notable distinction between maize and wheat plants towards salt application was observed in the present study. Wheat showed more tolerance towards salt treatments than maize, as indicated by response of accumulation of lower Na + content, higher K + and K + /Na + ratio. This suggests that maize may lack the ability to express some components to balance the increasing level of Na + and then stabilize the high K + /Na + ratio.
Wheat, a C3 plant, is better adapted to saline conditions due to higher proline and protein accumulation and POD activity than a C4 plants like maize and this pathway in C4 plants is usually argued as conferring greater tolerance to water deficits, high temperature and nitrogen deficiencies while, wheat shows contrasting response to these factors and had a better tolerance to salt and cold stresses (Loomis and Connor, 1992). The present findings indicated that higher proline and protein content could be one of the deciding factors, which offers advantage to wheat over maize for superior performance under saline conditions and suggests use of physiological markers such as Na + , K + and K + /Na + ratio for selecting the tolerant cultivars of maize.
REFERENCES
- Alscher, R.G., J.L. Donahue and C.L. Cramer, 1997. Reactive oxygen species and antioxidants: Relationship in green cells. Physiol. Plant., 100: 224-233.
CrossRefDirect Link - Agastian, P., S.J. Kingsley and M. Vivekanandan, 2000. Effect of salinity on photosynthesis and biochemical characteristics in mulberry genotypes. Photosynthetica, 38: 287-290.
CrossRefDirect Link - Ashraf, M. and P.J.C. Harris, 2004. Potential biochemical indicators of salinity tolerance in plants. Plant Sci., 166: 3-16.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Botella, M.A., V. Martinez, J. Pardines and A. Cerda, 1997. Salinity induced potassium deficiency in maize plants. J. Plant Physiol., 150: 200-205.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Chance, B. and A.C. Maehly, 1955. Assay of catalases and peroxidases. Methods Enzymol., 2: 764-775.
CrossRefDirect Link - Chinnusamy, V., A. Jagendorf and J.K. Zhu, 2005. Understanding and improving salt tolerance in plants. Crop Sci., 45: 437-448.
Direct Link - De Lacerda, C.F., J. Cambraia, M.A. Oliva, H.A. Ruiz and J.T. Prisco, 2003. Solute accumulation and distribution during shoot and leaf development in two sorghum genotypes under salt stress. Environ. Exp. Bot., 49: 107-120.
CrossRefDirect Link - Dionisio-Sese, M.L. and S. Tobita, 1998. Antioxidant responses of rice seedlings to salinity stress. Plant Sci., 135: 1-9.
CrossRefDirect Link - Fougere, F., D. Le Rudulier and J.G. Streeter, 1991. Effects of salt stress on amino acid, organic acid and carbohydrate composition of roots, bacteroids and cytosol of alfalfa (Medicago sativa L.). Plant Physiol., 96: 1228-1236.
PubMed - Goudarzi, M. and H. Pakniyat, 2008. Evaluation of wheat cultivars under salinity stress based on some agronomic and physiological traits. J. Agric. Soc. Sci., 1: 35-38.
Direct Link - Hernandez, J.A., A. Campillo, A. Jimenez, J.J. Alarcon and F. Sevilla, 1999. Response of antioxidant systems and leaf water relations to NaCl stress in pea plants. New Phytol., 141: 241-251.
CrossRefDirect Link - Jain, M., G. Mathur, S. Koul and N.B. Sarin, 2001. Ameliorative effects of proline on salt stress-induced lipid peroxidation in cell lines of groundnut (Arachis hypogaea L.). Plant. Cell Rep., 20: 463-468.
CrossRefDirect Link - Khatkar, D. and M.S. Kuhad, 2000. Short-term salinity induced changes in two wheat cultivars at different growth stages. Biol. Plant., 43: 629-632.
Direct Link - Lee, T.M. and C.H. Liu, 1999. Correlation of decreases calcium contents with proline accumulation in the marine green Macroalga ulva fasciata exposed to elevated NaCI contents in seawater. J. Exp. Bot., 50: 1855-1862.
Direct Link - Liu, J. and J.K. Zhu, 1997. An Arabidopsis mutant that requires increased calcium and potassium nutrition for salt tolerance. Proc. Natl. Acad. Sci. USA., 94: 14960-14964.
PubMed - Lutts, S., V. Majerus and J.M. Kinet, 1999. NaCl effects on proline metabolism in rice (Oryza sativa L.) seedlings. Physiol. Plant., 105: 450-458.
CrossRef - Manchandia, A.M., S.W. Banks, D.R. Gossett, B.A. Bellaire, M.C. Lucas and E.P. Millhollon, 1999. The influence of alpha-amanitin on the NaCl-induced up-regulation of antioxidant enzyme activity in cotton callus tissue. Free Radic. Res., 30: 429-438.
PubMedDirect Link - Meneguzzo, S., F. Navari-Izzo and R. Izzo, 1999. Antioxidative responses of shoots and roots of wheat to increasing NaCl concentrations. J. Plant. Physiol., 155: 274-280.
Direct Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Muthukumarasamy, M., S.D. Gupta and R. Pannerselvam, 2000. Enhancement of peroxidase, polyphenol oxidase and superoxide dismutase activities by triadimefon in NaCl stressed Raphanus sativus L. Biol. Plant., 43: 317-320.
Direct Link - Neill, S., R. Desikan and J. Hancock, 2002. Hydrogen peroxide signalling. Curr. Opin. Plant Biol., 5: 388-395.
CrossRefDirect Link - Noble, C.L. and M.E. Rogers, 1992. Arguments for the use of physiological criteria for improving the salt tolerance in crops. Plant Soil, 146: 99-107.
CrossRefDirect Link - Pakniyat, H., L.L. Handley, W.T.B. Thomas, T. Connolly, M. Macaulay, P.D.S. Caligari and B.P. Forster, 1997. Comparison of shoot dry weight, Na+ content and C13 values of ari-e and other semi-dwarf barley mutants under salt stress. Euphytica, 94: 7-14.
CrossRefDirect Link - Pakniyat, H. and M. Armion, 2007. Sodium and proline accumulation as osmoregulators in tolerance of sugar beet genotypes to salinity. Pak. J. Biol. Sci., 10: 4081-4086.
CrossRefPubMedDirect Link - Parida, A.K., A.B. Das, B. Mittra and P. Mohanty, 2004. Salt-stress induced alterations in protein profile and protease activity in the mangrove (Bruguiera parviflora). Z. Naturforsch, 59: 408-414.
PubMedDirect Link - Petrusa, L.M. and I. Winicov, 1997. Proline status in salt tolerant and salt sensitive alfalfa cell lines and plants in response to NaCl. Plant Physiol. Biochem., 35: 303-310.
Direct Link - Gonzalez, K.R., L. Erdei and S.H. Lips, 2002. The activity of antioxidant enzymes in maize and sunflower seedlings as affected by salinity and different nitrogen sources. Plant Sci., 162: 923-930.
CrossRefDirect Link - Sairam, R.K., K.V. Roa and G.C. Srivastava, 2002. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci., 163: 1037-1046.
CrossRefDirect Link - Schachtman, D. and W. Liu, 1999. Molecular pieces to the puzzle of the interaction between potassium and sodium uptake in plants. Trends. Plant Sci., 4: 281-287.
CrossRef - Singh, S.K., H.C. Sharma, A.M. Goswami, S.P. Datta and S.P. Singh, 2000. In vitro growth and leaf composition of grapevine cultivars as affected by sodium chloride. Biol. Plant., 43: 283-286.
CrossRefDirect Link - Wang, W., B. Vinocur and A. Altman, 2003. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta, 218: 1-14.
CrossRefPubMedDirect Link - Yeo, A., 1998. Molecular biology of salt tolerance in the context of whole-plant physiology. J. Exp. Bot., 49: 915-929.
CrossRefDirect Link - Zhu, J.K., 2003. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol., 6: 441-445.
CrossRefDirect Link