
Sadeq Khalid Nessr Alhag
Key Laboratory of Pesticide and Chemical Biology, Institute of Entomology,Huazhong Normal University, Ministry of Education, Wuhan 430079, China
Peng Jian Xin
Key Laboratory of Pesticide and Chemical Biology, Institute of Entomology,Huazhong Normal University, Ministry of Education, Wuhan 430079, China
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 9 | Page No.: 1756-1760







ABSTRACT
Six insect cell lines were tested for susceptibility to Syngrapha falcifera multiple nucleocapsid nucleopolyhedrovirus-D (SfaMNPV-D) infection by use of a typical endpoint assay procedure. Cell lines from Trichoplusia ni (Tn5B1-4), (L105-clone), Spodoptera litura (SL-ZSU-1), Spodoptera frugiperda (IPLB-SF-21), Pieris rapaeb (Pr-E-HNU9) and Helicoverpa zea (BCIRL-HZ-AM1) in 96-well tissue culture plates were infected with dilutions of extra cellular virus suspensions of (SfaMNPV-D). Each cell/virus combination was incubated at temperatures 27°C and wells were scored for positive infection at 2 to 4 day intervals. The resulting data were analyzed by Reed and Muench method, providing virus titers for each combination of virus, cell line. The results were categorized by accuracy and by rapidity of maximum titer. Virus titer of Tn5B-4 was higher than other cell lines TCID50 8.7x108, the lowest level detected in infected was in (Pr-E-HNU9) cells TCID50 2.4x108. No Virions or polyhedral inclusion bodies were detected in infected SL-ZSU-1 cells.
PDF Abstract XML References Citation
How to cite this article
Sadeq Khalid Nessr Alhag and Peng Jian Xin, 2008. Replication of Syngrapha falcifera Multiple-Nuclear Polyhedrosis Virus-D in Different Insect Cells. Journal of Applied Sciences, 8: 1756-1760.
DOI: 10.3923/jas.2008.1756.1760
URL: https://scialert.net/abstract/?doi=jas.2008.1756.1760
DOI: 10.3923/jas.2008.1756.1760
URL: https://scialert.net/abstract/?doi=jas.2008.1756.1760
INTRODUCTION
Nuclear Polyhedrosis Viruses (NPV) are members of the Baculovirus genus (Wildy, 1971) and have been isolated from insects in the orders Diptera, Hymenoptera, Lepidoptera and Orthoptera. The virion is bacilliform (40 to 70 by 250 to 400 nm) and contains covalently closed, supercoiled, double-stranded DNA with a molecular weight of approximately 108 (Summers and Anderson, 1972). The virus replicates in the nucleus of the insect cell and as the infection proceeds, a substantial proportion of the progeny are enveloped and subsequently occluded by protein into polyhedral, crystalline matrixes (0.1 to 10, um in diameter). The inclusions or polyhedra are readily detectable in the nuclei of infected insect cells and present a cytopathology that is characteristic of infections initiated by viruses of this genus (Harrap, 1972). Polyhedra represent the primary vector by which virus infections are transmitted in nature to insects. When larvae ingest foliage that is contaminated with polyhedra, the alkaline environment and enzymatic activity associated with the larval gut solubilized the protein matrix of the polyhedron, releasing infectious virions (Harrap, 1972). Polyhedra are not infectious for insect cell cultures (Ignoffo et al., 1971), but cultures do support the replication of these viruses when alternative sources of inoculum are used. For example, cultures have been infected with hemolymph of NPV-infected insects (Faulkner and Henderson, 1972), with purified NPV DNA (Vail et al., 1973), with NPV-infected cell culture extracts and with cell-free extracts of NPV-infected larvae. There is, however, only one report where the virus was purified from infected cell cultures and shown to be infectious (Henderson et al., 1974). Infectivity assays of polyhedra have relied primarily on a mean lethal dose fed to larvae. An end-point dilution. Method using a primary insect cell culture (Vaughn and Stanley, 1970) and a quantal response method using a continuous cell culture has also been used to assay infectious material other than polyhedra. More recently, the first plaque method using 0.6% methylcellulose as an overlay has been reported (Hink and Vail, 1973). In the current study six insect cell lines were tested for susceptibility to SfaMNPV-D infection by use of a typical endpoint assay procedure. Infectivity assay procedures for an end-point dilution method described and compared with particle count data and the particle-infectious.
MATERIALS AND METHODS
Cell lines: Insect cell lines used in this study are shown in Table 1. All the cell lines were maintained at 28°C and routinely subcultured every 2-3 day by seeding a 25 cm2 flask with 2×105 cells. The cell lines were generally grown in grace’s medium plus 10% Foetal Bovine serum (FBS) (Cynthil et al., 2001) cell lines were obtained from Laboratory of Pesticide and Chemical in Huazhong Normal University, China in 2004.
Spodoptera exigua larvae and virus: This virus was prepared by per as infection of third instar Spodoptera exigua larvae on an artificial diet (Bell et al., 1981). Infected larvae were surface disinfected by immersion in 70% ethanol for 10 min, air dried and then bled through a cut proleg into cold TC-100 medium (on ice) supplemented with 50 μg mL-1 gentamicin sulfate (Sigma) and 80 μg mL-1 reduced glutathione (Sigma). One milliliter of medium was used per larvae (infected larvae were approximately 2.5-3 cm long). The hemolymph-containing medium was centrifuged (15 min at 350 g) and the supernatant was passed through a 0.45 μm Uniflo filter. This filtrate was used to initiate infection.
Infection of cell lines: Viable cell counts were made by using trypan blue and a hemocytometer. After counting, cells were suspended by flushing with medium from a transfer pipet and diluted in their respective medium to 2x105 cells mL-1. Cells (10,000 in 0.15 mL) were distributed into each well of a 96-well plate The titer of viral suspension was determined by the end point dilution method as 50% tissue culture infective does (TCID50). The virus samples were diluted through 10-fold serial dilutions in modified TC-100 to 10-9 of the original. Aliquots (50 μL) of each dilution were inoculated onto each of 12 wells of the respective cell lines. The plates were sealed in airtight plastic boxes with moistened towels to maintain humidity and placed in incubators at 27°C. Cells were examined for cytopathic effect (the presence of occlusion bodies in the nuclei) at 2 to 4 day intervals up to 4 weeks post infection. The number of positive vs. negative wells for each dilution was noted and the TCID50 was calculated according to Reed and Muench (1938).
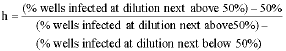 |
where, h is an interpolated log10 value of a dilution step. This value is added to the step above the 50% value.
RESULTS AND DISCUSSION
The quantitative bioassay to express the SfaMNPV-D: The quantitative bioassay to express the SfaMNPV-D titer as tissue culture infective dose 50% (TCID50) unit mL-1 ten- fold dilution of cells were inoculated into 8 wells of 96 wells plates and 3 and 5 days later, the plates were observed. The virus titer was calculated using the method of Reed and Muench (1938). Cell lines were compared to select the most suitable one for virus titration, the virus titer determined with Tn5B1-4. The result was confirmed by estimating the percentage of infected cells in each cell line following their infection at a high multiplicity of infection (5MOI/cell) and there wasn’t clear infection for S. litura (SL-ZSU-1) detected (Table 1).
Contact time between cell lines and viral inoculums: Preliminary experiments it was noticed that the efficiency of infection was improved by increasing the time of contact of the virus inoculums with the cells. In order to verify and quantify this observation, a series of titrations were performed by removing inoculums at regular intervals post-infection as shown in Fig. 1. In Tn5B1-4 line, virus titer reached only 3.5x108 TCID50 mL-1 after 12 h the virus titer increased with increasing time of contact up to a plateau 8.7×108 TCID50 mL-1 when the inoculum was maintained 72 h suggested this line was one of the most susceptible to SfaMNPV-D while the (L105) line reach the maximum titer (6.5×108 TCID50 mL-1) at 96 h post infection (Fig. 1a, d, respectively). IPLB-SF-21 cells reached to 4.7×108 mL-1 when the inoculum was maintained 96 h. The BCIRL-HZ-AM1 and Pr-E-HNU9 cells were found to have the lowest infection with inoculum for 72 h, it reached to 3.6×108 mL-1 and 2.4×108 TCID50 mL-1, respectively (Fig. 2) suggesting these two lines were less susceptible to SfaMNPV-D than the other cells used in this study.
| Table 1: | Cell lines used |
 | |
 | |
| Fig. 1: | Micrographs of cells infected with SfaMNPV-D. (a) Tn5B1-4 cells, (b) IPLB-SF-21 cells, (c) Pr-E-HNU9 cells (d) L105-clone cells, (e) BCIRL-HZ-AM1 cells and (f) S. litura (SL-ZSU-1) uninfected cells |
The comparisons of SfaMNPV-D in the six cell lines revealed the virus showed the highest titer in each line when incubated 96 h with the exception of SL-ZSU-1 cell line no virions or polyhedral inclusion were detected during this time.
The growth cycle data demonstrates that infectious material was released from infected cells 12 h post infection, reaching a maximal titer 96 h post infection. Cytolysis did occur to a limited extent 2 to 3 days post infection and perhaps the adsorption of virus to newly exposed receptor sites accounts for the drop in the interval titer. It is interesting to note that lateral transmission of infectivity occurred 1 to 2 days before any lysis was observed. Virus, however, has been detected in the nuclei of infected cells 12 h post infection and has been observed in the process of budding from the cell surfaces. Several conditions were established for the optimal replication of SfaMNPV-D in a continuous cells culture. The temperature optimum for the growth of the cells of 27°C was also the optimal temperature for the replication of the virus.
Therefore, the differences infection between differences cell lines must be due to differences in the early stages of the infection process to the cell membrane, penetration and transport to the nucleus. The virus envelope fuses with the resulting (Hefferon et al., 1999) acidified endocytic vesicle, releasing the nucleocapsid into the cell cytoplasm followed by its transportation to the nuclear membrane. This cell lines appear to have a good potential for studying baculoviruses and, with some further optimization of the growth condition they may also be useful in production scheme for this virus and this results indicate that Tn5B1-4 and L105-clone cells display cytopathic response following infection with SfaMNPV-D (Fig. 1).
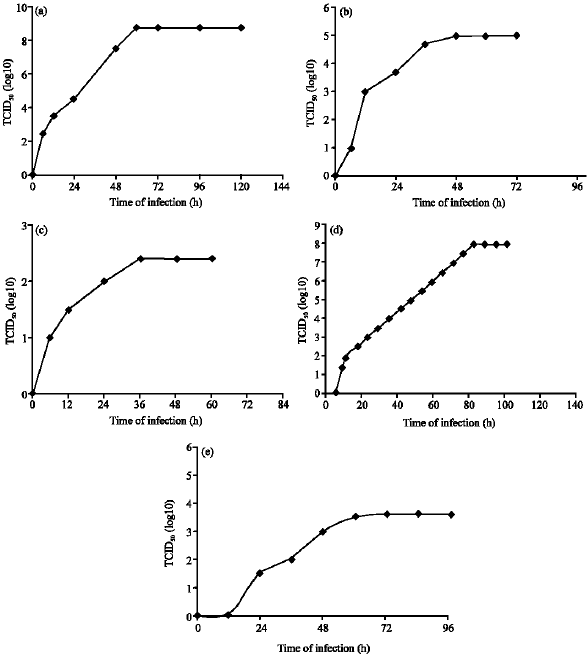 | |
| Fig. 2: | Dynamic curves of SfaMNPV-D propagation in different cell lines. (a) Tn5B1-4 cells, (b) IPLB-SF-21 cells, (c) Pr-E-HNU9 cells, (d) L105-clon cells and (e) BCIRL-HZ-AM1 cells |
Thus Tn5B1-4 and L105-clone cells provide an excellent system for understanding the molecular mechanisms of NPV-cell interactions. No virions or polyhedral inclusion bodies were detected in infected SL-ZSU-1 cells.
REFERENCES
- Bell, R.A., C.D. Owens, M. Shapiro, J.G.R. Tardif, C.C. Doane and M.L. McManus, 1981. Development of mass-rearing technology. The gypsymoth: research toward integrated pest management and development program. USDA Tech. Bull., 1584: 599-633.
Direct Link - Goodman, C.L., G.N. El-Sayed, A.H. Mcintosh and J.J. Grasela, 2001. Establishment and characterization of insect cell lines from 10 Lepidopteran species. In vitro Cell. Dev. Boil. Anim., 37: 367-373.
Direct Link - Faulkner, P. and J.F. Henderson, 1972. Serial passage of a nuclear polyhedrosis disease virus of the cabbage looper (Trichoplusia ni) in a continuous tissue culture cell line. Virology, 50: 920-924.
CrossRefDirect Link - Harrap, K.A., 1972. The structure of nuclear polyhedrosis viruses: III. Virus assembly. Virology, 50: 133-139.
CrossRefDirect Link - Hefferon, K.L., A.G.P. Oomens, S.A. Monsma, C.M. Finnerty and G.W. Blissard, 1999. Host cell receptor binding by baculovirus GP64 and kinetics of virion entry. Virology, 258: 455-468.
CrossRefDirect Link - Henderson, J.F., P. Faulkner and E.A. MacKinnon, 1974. Some biophysical properties of virus present in tissue cultures infected with the nuclear polyhedrosis virus of Trichoplusia ni. J. Gen. Virol., 22: 143-146.
CrossRefDirect Link - Hink, W.F. and P.V. Vail, 1973. A plaque assay for titration of alfalfa looper nuclear polyhedrosis virus in a cabbage looper (TN-368) cell line. J. Invertebr. Pathol., 22: 168-174.
CrossRefDirect Link - Ignoffo, C.M., M. Shapiro and W.F. Hink, 1971. Replication and serial passage of infectious Heliothis nucleopolyhedrosis virus in an established line of Heliothis zea cells. J. Invertebr. Pathol., 18: 131-134.
Direct Link - Mitsuhash, J., S. Hayasaka and S. Imanishi, 2003. Continuous cell lines from the common white, Pieris rapae crucivora boisduval. In vitro Cell. Dev. Biol. Anim., 39: 114-116.
Direct Link - Reed, L.J. and H. Muench, 1938. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol., 27: 493-497.
CrossRefDirect Link - Summers, M.D. and D.L. Anderson, 1972. Characterization of deoxyribonucleic acid isolated from the granulosis viruses of the cabbage looper, Trichoplusia ni and the fall armyworm, Spodoptera frugiperda. Virology, 50: 459-471.
CrossRefDirect Link - Vail, P.V., D.L. Jay and W.F. Hink, 1973. Replication and infectivity of the nuclear polyhedrosis virus of the alfalfa looper, Autographa californica, produced in cells grown in vitro. J. Invertebr. Pathol., 22: 231-237.
CrossRefDirect Link - Vaughn, J.L. and M.S. Stanley, 1970. A micromethod for the assay of insect viruses in primary cultures of insect tissue. J. Invertebr. Pathol., 16: 357-362.
CrossRefDirect Link - Vaughn, J.L., R.G. Goodwin, G.J. Tompkins and P. McCawiey, 1977. The establishment of two cell lines from the insect Spodoptera frugiperda (lepidoptera; noctuidae). In vitro, 13: 213-217.
CrossRefDirect Link