
Mourad L. Shonouda
Department of Zoology, Faculty of Science, 21511 Mharam Bey, Alexandria University, Egypt
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 9 | Page No.: 1692-1698







ABSTRACT
The antennal response of adult syrphid flies to selected plant volatile chemical compounds was investigated in the present study. The main chemical classes and their chemical compounds were aldehydes (nonanal and benzaldehyde), monoterpene-alcohols (linalool and alpha-terpineol), ketones (6-methyl-5-heptene-2-one and 2-undecanone), hydrocarbons (tetradecane) and benzoids (methyl salicylate). Electroantennogram (EAG) records showed that the syrphid antennae were strongly responded to linalool, 6-methyl-5-heptene-2-one and methyl salicylate even at low concentrations, in addition to the high dose concentration of nonanal comparably to the other chemical compounds. The antennae of old syrphid adults were more responsive and elicited higher levels of responses to all compounds rather than young syrphid adults. The antennal sensitivity may differ from one compound to another according to the sex. The difference in responses could be attributed to the sensitivity of olfactory receptors and/or the characterization of binding protein(s). The quality of biocontrol agent could be improved if the chemical interaction between beneficial natural enemies and the surrounding environment is intensively studied and we clearly understand the chemical ecology of each natural enemy.
PDF Abstract XML References Citation
How to cite this article
Mourad L. Shonouda, 2008. New Approach to Evaluate the Antennal Response of an Adult Predator Insect to Different Volatile Chemical Compounds by using Electroantennogram Technique. Journal of Applied Sciences, 8: 1692-1698.
DOI: 10.3923/jas.2008.1692.1698
URL: https://scialert.net/abstract/?doi=jas.2008.1692.1698
DOI: 10.3923/jas.2008.1692.1698
URL: https://scialert.net/abstract/?doi=jas.2008.1692.1698
INTRODUCTION
Hover flies (diptera: syrphidae) are among the most common species of natural enemies of aphids. The syrphid fly Episyrphus balteatus is of a potential economic importance because larvae are predator of a number of aphids that are serious agriculture pests (Hart and Bale, 1997). Predator and prey come into contact through the behavior of the females that oviposit near suitable larval food source (Hickman et al., 1995). Olfaction and vision are the two main senses which guide many predators to habitats that may harbour their preys (Huffaker and Messenger, 1976). Numerous studies have shown that plant volatiles serve as modifiers of olfactory responses to semiochemicals (Dickens et al., 1993; Das et al., 2007). Particularly, several species of flowers have been shown to be attractive to dipterous insects (Loevei et al., 1993; Alves et al., 2005). Ziziphus flowers are one of the plant species that are attractive to many species of syrphid flies and hymenopterous parasitoids (Shonouda, 2003). Flower extracts of Ziziphus spina christi showed allelochemical effects by increasing the arrestant time, number of visiting and the parasitization rate of female parasitoids as compared with other parts of the plant (Shonouda, 2005). The main identified volatile compounds in flowers of Z. spina christi are belonging to chemical classes aldehydes, monoterpene-alcohols, ketones, hydrocarbons and benzene compounds (Shonouda et al., 2008). Chemical compounds belong to these chemical classes are selected to bioassay their effectiveness and their capacity of inducing electrophysiological activity by establishing dose-response curves of antennae of adult syrphid flies Episyrphus balteatus. Also, a novel concept for differentiation between sexes and ages of the predator insect was investigated. It is imperative to know the chemical interaction between predator syrphid flies and the surrounding semiochemicals to enhance the biological control potential of a natural enemy.
MATERIALS AND METHODS
Rearing of episyrphus balteatus: The pupae of the syrphid fly Episyrphus balteatus (DeGeer) were bought from KATZ BIOTECH Ag Company, Baruth, Germany and kept in large cages until emergency of adults. Each cage provided by pollen, a cotton pad with water and sugar cups. Cages were kept at 26°C, 60% RH in a LD 18:6 h light cycle. Syrphid adults need 14 days for feeding, mating and ovulation (Hart and Bale, 1997). Males and females with two different ages were selected for experiments. The syrphid adults ranged between 6-11 days selected as young adults, while between 16-22 days of males and gravid females were selected as old adults for dose-response experiments. All syrphid adults were selected as naïve adults (have no previous contact with aphids). All experiments were conducted at Georg-August University, Goettingen, Germany during the period from 2006-2007.
Odour stimuli: A range of common as well as specific plant volatile compounds were selected to study the antennal response profile of adult female and male Episyrphus balteatus (Pickett et al., 2005; Das et al., 2007; Shonouda et al., 2008). Chemicals belong to different classes were selected as ketone, aldehyde, monoterpene-alcohol, hydrocarbons and benzoids. The following compounds obtained from commercial suppliers were measured: 6-methyl, 5-haptene-2-one (Fluka), 2-undecanone (Aldrich), nonanal (Acros), benzaldeyde (Acros), linalool (Merck), alpha terpineol (Aldrich), tetradecane (Acros), methyl salicylate (Acros), in addition to paraffin oil (Merck) as control and cis-3-hexen-1-ol (Acros) as a standard control.
Electroantennogram (EAG): For electrophysiological investigations, insect antenna was cut from narcotized female and male syrphid flies and fixed in an antennal holder filled with Ringer solution. Within this holder the ends of the antennae were in touch with electrolyte solution adapted to the insect's hemolymph which provides electrical contact to a pair of Ag/AgCl electrodes. Amplifier setup was used for amplification of potentials resulting in a total amplification of 100 units. The amplified signal was recorded by the Agilent GC ChemStation software using the type 35900E A/D converter (Agilent). The A/D converter was connected to the acquisition system and synchronizes by the start signal of the gas chromatography (GC) (Weissbecker et al., 2004; Johne et al., 2006). Dose-response series of insect antennae were measured by manual puffing of a fixed volume of air (5 mL) from glass syringes which contained pieces of filter paper (2 cm2) drenched with 10¯8, 10¯6, 10¯4 and 10¯2 standard dilutions of stimulus compounds in paraffin oil (μg/μL) in addition to cis-3-hexen-1-ol (10¯3) as a standard control. A minimum five antennae originated from different syrphid flies were tested per compound and resulting signals were amplified and recorded.
Statistical analysis: To compensate for artifacts, EAG values (mV) were corrected by subtracting the control paraffin oil value alone from the obtained values of each stimulus compound. After that, the data were standardized by expressing the corrected mean EAG values as a percentage of the standard stimulus. The data within each individual compound for each sex was analyzed by using one-way ANOVA and between means of different ages and between sexes were determined by Student t-test (Zar, 1984).
RESULTS
The male and female antennae were stimulated with different compounds and showed strong responses to most compounds (Fig. 1, 2).
Differentiation between dose responses within the individual compound: Within the same chemical compound, antennal response increase with the increase of dose concentration of each compound. The response of old antennae was higher and pronounced than young ones. Generally, the difference between mean dose response within each chemical compound of young antennae was insignificant (p>0.05), so the present results focused on the dose response of old antennae to different concentrations within the individual compound according to different chemical classes.
Ketones: The response of female and male antennae to 6-methyl-5-heptene-2-one was high and increased with increase the dose concentrations but the difference between the means within the individual compound was insignificant (p>0.05). However, females and males were highly responded (87.2%, 61.3%) respectively, to the lowest concentration (10¯8) of 6-methyl-5-heptene-2-one in compare to the same lowest concentration of all compounds belong to different chemical classes. Contrarily, the response of female and male antennae to 2-undecanone was low and increased with the increase of doses and was pronounced with the highest concentration (10¯2). The mean dose response at the highest concentration (10¯2) with either females or males was highly significant (F = 31.16, 21.46, p<0.01), respectively, as compared with other low concentrations.
Monoterpene alcohol: Female and male antennae were highly responded to linalool. The difference between means of dose response was significant (F = 57.1, p<0.01 and F = 18.1, p<0.05), respectively. Also, the response of females and males to different concentrations of alpha-terpineol was significantly increased (F = 22.1, 35.7; p<0.05), respectively. For both compounds, the significant difference was in favor to the highest concentration (10¯2). It was also observed that the dose responses of both sexes at all concentrations of linalool were higher than that of other compounds (Fig. 1).
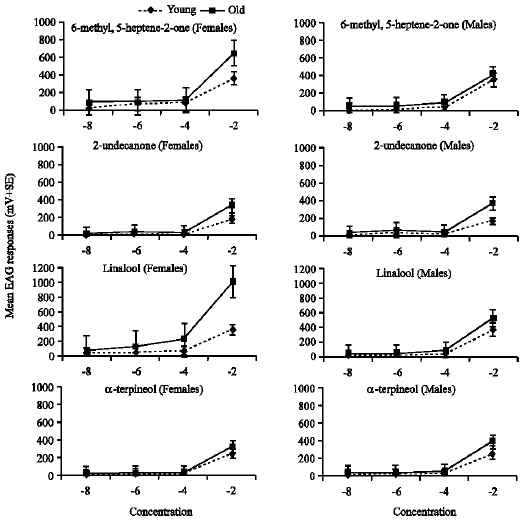 | |
| Fig. 1: | Normalized EAG dose-response profiles for female and male E. balteatus antennae to ketone and monoterpene alcohol volatile compounds |
Aldehydes: The response of female and male antennae was significantly increased with dose concentrations of nonanal (F = 37.9, 217.6; p<0.01), respectively. The mean difference of females was significantly increased between concentrations (10¯6) and (10¯4), while for males between the highest concentration (10¯2) and the other lower concentrations. Concerning benzaldehyde, the response of both sexes were increased gradually but there was no significant difference between means (p>0.05). It was clear that the males were highly responded (77.1%) to the concentration (10¯6) of benzaldehyde comparable to the same dose of all other compounds.
Hydrocarbone: The response of both sexes to different concentrations of tetradecane was more-or-less similar and there was no significant difference between the means (p>0.05). The response of both sexes to tetradecane was lower at doses 10¯6, 10¯4 and 10¯2 as compared with the same doses of other compounds belonging to different chemical classes.
Benzoids: Female and male antennae were gradually responded to different concentrations of methyl salicylate but there was no significant difference between means (p>0.05), however, females responded highly (169.8%) to the concentration (10¯6) of methyl salicylate comparable to the same concentration of all other compounds.
Differentiation between dose responses of different ages: The old adult antennae of both sexes showed higher responses to all compounds than the young antennae. The higher doses gave better results than lower doses for all selected compounds as shown in the following chemical classes:
Ketones: The old antennae responded higher than young ones to the different concentrations of 6-methyl-5-heptene-2-one. The mean difference was significantly higher (t = 2.8, p<0.05) at only the highest concentration (10¯2) of the female while there was no significant difference between mean doses of males. Also, the old antennae respond higher to 2-undecanone than young ones for both sexes.
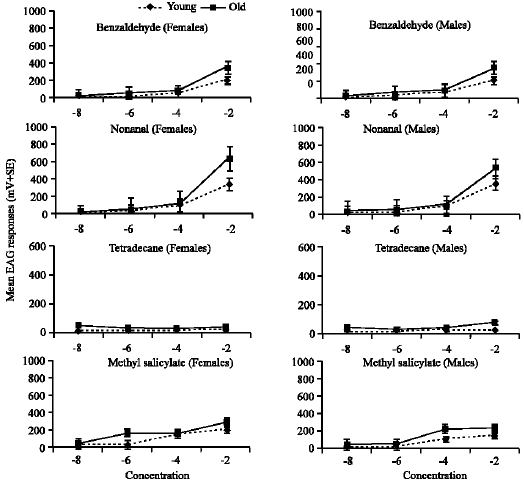 | |
| Fig. 2: | Normalized EAG dose-response profiles for female and male E. balteatus antennae to aldehyde, hydrocarbon and benzoic volatile compounds |
The mean difference was significantly high (t = 3.6, 3.5; p<0.05) for females and males, respectively at the highest concentration (10¯2).
Monoterpene alcohol: Both young and old antennae responded highly to different concentrations of linalool. The mean difference was significantly high (t = 3.4, p<0.05) at the highest concentration but the difference was insignificant for males. Also, the response of both antennae was high with alpha terpineol and there was significant difference (t = 3.1, 2.8; p<0.05) between means at the highest concentration (10¯2) for females and males, respectively.
Aldehydes: Old antennae of both sexes showed high response to nonanal and benzaldehyde in compare to young ones. The mean difference for females and males was significantly high at the highest concentration (10¯2) of nonanal (t = 5.1, 4.7; p<0.01) and of benzaldehyde (t = 4.1, p<0.01 and t = 2.9, p<0.05), respectively.
Hydrocarbons and benzoids: However the old antennae responded highly to the hydrocarbon tetradecane and the benzoid methyl salicylate than young ones but there was no significant difference (p>0.05) between means of different doses for both compounds.
Differentiation between dose responses of both sexes: Comparison of dose responses between females and males syrphid adults of either young or old antennae revealed differences in response for most compounds (Fig. 1, 2).
The young antenna of females responded highly to the range of dose concentrations (10¯8 - 10¯2) of 6-methyl-5-heptene-2-one (30.6-360), linalool (45.6-352.3%), methyl salicylate (29.4-210.9%) and nonanal (8.0-323.9%), while males responded less than females to 6-methyl-5-heptene-2-one (12.9-341%) and methyl salicylate (18-156.8%) and responded slightly high to linalool (7.6-368.4%) and nonanal (16.5-354.4%) especially at the highest concentration. The lowest response was to tetradecane with either females (9.2 to 19.8%) or males (24.7 to 27.7%) as compared with the other compounds.
Concerning the old antennae, the female responded strongly and increased highly to the same range of dose concentrations of linalool (53.1-994.5%), 6-methyl-5-heptene-2-one (87.2-650.5%) and nonanal (18.1-616%) while the response was weak and not increased with tetradecane (42.7-38.2%). On the other hand, the male responded strongly to the same range dose concentrations of linalool (50.5-530.8%), 6-methyl-5-heptene-2-one (61.3-418.5%) and nonanal (36.2-525.5%) but less than the females except with tetradecane (46.7-67.8%). Concerning the other compounds, the old antennae of males responded to benzaldehyde (26.3-352.5%); α-terpineol (40.8-455.4%) and 2-undecanone (42.8-360.9%) slightly higher than females (18.9-340.9%); (25.6-323.6%) and (19.9-334.7%), respectively. The mean difference between sexes was insignificant (p>0.05) for each compound and age.
DISCUSSION
Natural enemies respond to a large array of different plant volatile chemicals. It was previously demonstrated in the field experiment that the plant Z. spina christi attracted a large variety of insects during the flowering season and most of them are natural enemies (Shonouda, 2003). The identified volatile chemicals of Ziziphus flowers were belonging to different chemical classes as ketone, monoterpene alcohol, aldehyde, hydrocarbon and benzoid (Shonouda et al., 2008). In the present study, the electrophysiological response of syrphid antennae to characteristic compounds of floral odorant of Z. spina christi was evaluated by using EAG. The change in potential between the tip of the antenna and its base in response to stimulating by an odour component could be measured by the electroantennogram (EAG). An EAG reflects the electrical responses of the olfactory cell population in the antenna (Das et al., 2007). The female and male antennae of syrphid fly responded strongly to most compounds and the old antennae showed higher responses than the young one for all compounds. The response of old females was more pronounced and stronger than males to linalool, 6-methyl-5-heptene-2-one, nonanal and methyl salicylate especially at the highest concentration (10¯2), where the female antennae being more sensitive to these compounds. On the other hand, males were more responsive and sensitive to benzaldehyde, tetradecane, α-terpineol and 2-undecanone at all concentrations. The weak response of both sexes to tetradecane may be due to the fact that the volatility of hydrocarbon compound is lower than the other volatile compounds.
It is of interest to note that the old antennae of females and males were relatively responded greater to low concentrations of 6-methyl-5-heptene-2-one, linalool and methyl salicylate than the other compounds. The antennal responses were significantly increased with the increase of concentration of linalool and nonanal, while, the response to methyl salicylate and 6-methyl-5-heptene-2-one did not increase significantly (p>0.05) with increasing the concentration. Definitely, the monoterpene linalool showed high EAG activity using E. balteatus antennae of both sexes comparable to the other compounds as found by Verheggen et al. (2005). The pronounced EAG responses of both sexes of syrphid flies to low concentrations of linalool (monoterpene alcohol), 6-methyl-5-heptene-2-one (ketone) and methyl salicylate (benzoid) gave a good indication that the syrphid adults are very sensitive to these compounds even at low concentrations regardless their chemical classes. Accordingly, these compounds may be play significant role in prey or habitat location. The same conclusion was reported by Zhu and Park (2005) who found that methyl salicylate elicited significant electrophysiological responses in the predator C. septempunctata suggesting that the predator may be use methyl salicylate as olfactory cue for prey location. Also, it was known that many plants release minute amount of volatile methyl salicylate and 6-methyl-5-heptene-2-one under attack by pests as external plant stress signaling (Bi et al., 2007). This phenomenon is known as cry for help (Forouhar et al., 2005) to attract great number of species of natural enemies (James and Price 2004; Pickett et al., 2005). Also, monoterpene alcohol compounds including linalool are characteristic compounds of most flowers and their allelochemical effects were proved on different beneficial insects (Du et al., 1998; Georgieva et al., 2005).
The ability of both females and males to detect the same odours is probably due to their similar habitat, which requires the use of the same cues to locate host plants for survival and reproduction. This explains why there was insignificant difference between dose responses of both sexes. However, the variety between antennal response of female and male syrphid adults may be due to the sensitivity of olfactory receptor cells to plant volatiles suggesting adaptation of the set of olfactory receptors on antennae of each sex to particular flower odorants to locate suitable feeding or mating sites in addition to oviposition site for gravid females. In some insect species, the greater response exhibited by females compared with males to plant volatiles have been reported regarding that the female antennae are endowed with a larger population of olfactory receptor cells that are sensitive to plant volatiles (Zhang et al., 1999; Das et al., 2007). Also, other differences found between young and old syrphid flies may be due to the change of their interest relatively to the age. The newly emerged flies take necessary for mate seeking instead of eating pollen while pollen is of great dietary importance for gravid syrphid females (Hickman et al., 1995).
It could be concluded that the antennae of syrphid flies are very sensitive and elicited higher levels of responses to most of the odorant compounds, but there was difference between antennal responses to chemicals even within the same classes. The chemical structure, volatility and concentration of the individual compound in addition to sex and age of E. balteatus adults influence the EAG responses (Park et al., 2001; Das et al., 2007). These variations in dose-response evaluation could be attributed to the sensitivity of the olfactory receptor system of syrphid fly (Das et al., 2007) and the characterization of the odorant binding protein(s), which may be varied between sexes and ages. Finally, gravid syrphid females, which lay their eggs giving rise to predatory larvae, are crucial for biocontrol agent. Habitat manipulation by providing suitable flower volatile chemicals could be used as additive stimulants to increase the density of syrphid flies and this associated with increasing oviposition in target area. The attraction of the predator syrphid flies by applying selective flower allelochemicals in the target area is a management option that is likely to lead to enhance control of pest populations.
ACKNOWLEDGMENTS
I would like to thank the Arab Fund for Economic and Social Development in Kuwait for supporting this research project according to the Academic Distinguished Grant of The Arab Fund Fellowship Program. Also, my thanks are extending to Dr. Stefan Vidal and Dr. Stefan Scuetz, Georg-August University, Goettingen, Germany for providing all facilities to complete the present research project.
REFERENCES
- Alves, R.J.V., A.C. Pinto, D.V.M. Coste and C.M. Rezende, 2005. Ziziphus mauritiana Lam. (Rhamnaceae) and the chemical composition of its floral fecal odor. J. Braz. Chem. Soc., 16: 654-656.
CrossRef - Bi, H.H., R.S. Zeng, L.M. Su, M. An and S.M. Luo, 2007. Rice allelopathy induced by methyl jasmonate and methyl salicylate. J. Chem. Ecol., 33: 1089-1103.
CrossRef - Das, P.D., R. Raina, A.R. Prasad and A. Sen, 2007. Electroantennogram responses of the potato tuber moth, Phthorimaea operculella (Lepidoptera: Gelichiidae) to plant volatiles. J. Biosci., 32: 339-349.
Direct Link - Dickens, J.C., J.W. Smith and D.M. Light, 1993. Green leaf volatiles enhance sex attractant pheromone of the tobacco budworm, Heliothis virescens (Lep.: Noctuidae). Chemoecology, 4: 175-177.
CrossRefDirect Link - Du, Y., G.M. Poppy, W. Powell, J.A. Pickett, L.J. Wadhams and C.M. Woodcock, 1998. Identification of semiochemicals released during aphid feeding that attract parasitoid Aphidius ervi. J. Chem. Ecol., 24: 1355-1368.
CrossRefDirect Link - Georgieva, E., N. Handjieva, S. Popov and L. Evstatieva, 2005. Comparative analysis of the volatiles from flowers and leaves of three Gentiana species. Biochem. Syst. Ecol., 33: 938-947.
Direct Link - Forouhar, F., Y. Yang, D. Kumar, Y. Chen and E. Fridman et al., 2005. Structural and biochemical studies identify tobacco SABP2 as a methyl salicylate esterase and implicate it in plant innate immunity. Proc. Nat. Acad. Sci., 102: 1773-1778.
Direct Link - Hart, A.J. and J.S. Bale, 1997. Cold tolerance of the aphid predator Episyrphus balteatus (DeGeer) (Diptera, Syrphidae). Physiol. Entomol., 22: 332-338.
CrossRefDirect Link - Hickman, J.M., G.L. Loevel and S.D. Wratten, 1995. Pollen feeding by adults of the hoverfly Melanostoma fasciatum (Diptera: Syrphidae). N. Z. J. Zool., 22: 387-392.
CrossRefDirect Link - James, D.G. and T.S. Price, 2004. Field-testing of methyl salicylate for recruitment and retention of beneficial insects in grapes and hops. J. Chem. Ecol., 30: 1613-1628.
Direct Link - Loevei, G.L., D.J. Hodgson, A. Macleod and S.D. Wratten, 1993. Attractiveness of Some Novel Crops for Flower-Visiting Hoverflies (Diptera: Syrphidae): Comparison from Two Continents. In: Pest Control and Sustainable Agriculture, Corey, S.A., D.J. Dall and W.M. Milne (Eds.).Caberra, Caberra, pp: 368-370.
- Park, K.C., J. Zhu, J. Harris, S.A. Ocheing and T.C. Baker, 2001. Electroantennogram responses of a parasitic wasp, Microplitis croceipes, to host-related volatile and anthropogenic compounds. Physiol. Entomol., 26: 69-77.
Direct Link - Shonouda, M.L., S. Angeli, S. Schutz and S. Vidal, 2008. Use of CLSA and SPME- headspace techniques followed by GC-MS analysis to extract and identify the floral odorants. Pak. J. Biol. Sci., 11: 1246-1251.
Direct Link - Weissbecker, B., G. Holighaus and S. Schutz, 2004. Gas chromatography with mass spectrometric and electroantennographic detection: Analysis of wood odorants by direct coupling of insect olfaction and mass spectrometry. J. Chromatogr., 1056: 209-216.
Direct Link - Zar, J.H., 1984. Biostatistical Analysis. 2nd Edn., Prentice-Hall Inc., Englewood Cliffs, New Jersey, USA, Pages: 718.
Direct Link - Zhang, A.J., C. Linn, S. Wright, R. Prokopy, W. Reissig and W. Roelofs, 1999. Identification of a new blend of apple volatiles attractive to the apple maggot, Rhagoletis pomonella. J. Chem. Ecol., 25: 1221-1232.
Direct Link - Zhu, J. and K.C. Park, 2005. Methyl salicylate, a soybean aphid-induced plant volatile attractive to the predator Coccinella septempunctata. J. Chem. Ecol., 31: 1733-1740.
Direct Link - Pickett, J.A., M.A. Birkett, T.J.A. Bruce, K. Chamberlain and R. Gordon-Weeks et al., 2005. CIS-Jasmone as an allelopathic agent through plant defence induction. Proceeding of the 4th World Congress on Allelopathy, Waaga Waaga, Australia, Aug. 21-26. http://www.regional.org.au/au/allelopathy/2005/1/3/2481_pickettja.htm.