
F. Ghassemzadeh
Department of Biology, Faculty of Sciences, Water and Environmental Center,Ferdowsi University of Mashhad, Iran
H. Yousefzadeh
Department of Biology, Faculty of Sciences, Payam-Noor University, Tehran, Iran
M.H. Arbab-Zavar
Department of Chemistry and Environmental Chemistry Center, Ferdowsi University of Mashhad, Iran
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 9 | Page No.: 1668-1675







ABSTRACT
Arsenic (As) and Antimony (Sb) are toxic heavy metals that often associated in contaminated environment. High As concentration is reported in Chelpo, Khorasan province, northeast Iran. This study examined the possibility of As and Sb in rhizofiltration by common reed, Phragmites australis. Plants collected from five sampling sites of Chelpo. As and Sb concentrations in roots and shoots were determined by Atomic absorption spectrometry. About 80% of total As accumulated in roots than shoot system. Maximum As and Sb accumulation in root and rhizomes were 84.5-16.20 and 73-10.20 ppb, respectively. As/Sb ratio of root and rhizome ranged from 35 to 194 and 10.42 to 99.9, respectively. Arsenic contents in roots of contaminated plants were significantly higher than antimony accumulation (p<0.01). There was antagonistic interaction between As, Sb with P contents in the roots of contaminated plants. Common reed establishment may be locally enhanced by significantly decreased phosphorous (1.79 and 1.18 times less than control plant root and rhizome), increased Iron (1.42 and 5.83 times more than control plant root and rhizome) Enzymatic antioxidants (Catalase, Ascorbate peroxidase and Guaicol peroxidase) increased significantly with increment of As uptake in root system. Root As and Sb accumulation with enhanced changes in metabolic changes indicates that P. australis could be used as As, Sb rhizofiltration agent in this region and similar contaminated area.
PDF Abstract XML References Citation
How to cite this article
F. Ghassemzadeh, H. Yousefzadeh and M.H. Arbab-Zavar, 2008. Removing Arsenic and Antimony by Phragmites australis: Rhizofiltration Technology. Journal of Applied Sciences, 8: 1668-1675.
DOI: 10.3923/jas.2008.1668.1675
URL: https://scialert.net/abstract/?doi=jas.2008.1668.1675
DOI: 10.3923/jas.2008.1668.1675
URL: https://scialert.net/abstract/?doi=jas.2008.1668.1675
INTRODUCTION
Heavy metals like Arsenic and Antimony are toxic pollutants released into the surface and ground water as a result of both anthropogenic and natural inputs (Adriano, 2001). Anthropogenic sources include mining and smelting processes besides application of As-based insecticides, herbicides, fungicides, algaecides, sheep dips, wood preservatives, dye stuffs, feed additives and compounds for the eradication of tapeworm in sheep and cattle. Geochemical sources of As-contaminated soils include As-rich parent materials as As easily substitutes for Si, Al or Fe in silicate minerals (Fitz and Wenzel, 2002). Analysis of fresh Platyphyllous cattail acid extracts has demonstrated that As and Sb present a serious threat to human and animal health, as they are capable of entering food chains in large amount (Hozhina et al., 2001). Arsenic and antimony can also co-exist in the environment (Gebel, 2000). These two elements occur as a major constituent in more than 200 minerals on the earth and are derived from ore minerals or alteration products that naturally contaminate the environments (Smedley and Kinniburgh, 2002). Although there are many studies about exposure to arsenic, not many studies address exposure to Sb. At present, a number of technologies can be used to remove As and Sb from contaminated substrates such as filtration, adsorption, chemical precipitation and ion-exchange (EPA, 2003). However, these methods are not efficient in removing low heavy metals concentrations, can be relatively expensive and may fail to achieve legal limits. Therefore, there is an urgent need for the development of innovate processes which can remove low concentrations of heavy metals economically (Volesky, 2000). Phytoremediation is an emerging technology driven by solar energy that uses green plants to remove toxic metals from contaminated aquatic and terrestrial sites (Suresh and Ravishankar, 2004). Phytostabilisation is one branch of phytoremediation that it uses of plants to immobilize contaminants in the soil and ground water through absorption and accumulation by root or precipitation within rhizosphere. This process that also called rhizofiltration, reduces the mobility of contaminated and prevent migration to the ground water and air and reduces the bioavailability for entry into food chain (Suresh and Ravishankar, 2004).
Some wetland plants seem to have developed a tolerance to heavy metals (Wu, 1990). Several emergent species (Typha spp., Canna spp., Colocasia esculenta, Heliconia psittacorum and Thalia dealbato) have been reported to hyper-accumulate arsenic. In 2004 Aksorn and Visootviseh reported that these plants accumulated 8.76-43.9 mg kg–1 arsenic from water. Identification of plant species suitable for phytostabilisation is best done by observation of plants that naturally colonize the contaminated site (Craw et al., 2007). The present study takes this approach and examine common reed (Phragmites. australis) for rhizofiltration. This has recently been considered for remediation of heavy metals such as cadmium (Ederli et al., 2004), copper (Ali et al., 2002), persistent organic pollutants like DDT and PCBS (Chu et al., 2006) and hydrated nickel and copper nitrate from treating urban run off (Lee and Scolz, 2007) and remediation heavy metals such as Cu+2, Ni+2, Cd+2, Zn+2 and Pb+2 (Southichak et al., 2006).
In the present study, we collected P. australis from five contaminated sample collection of the Chelpo area. This area is part of the Kohsorkh mountain range and is located north of Kashmar in the Khorasan Province of Northeastern Iran. Ghassemzadeh et al. (2007) reported the Chelpo River and its tributaries were contaminated with arsenic and antimony. Arsenic and antimony in this naturally polluted area (sampling sites 1, 2, 3, 4, 5) are derived from fault zone where the Paleogene deposits have been cut by faults and the ore deposits formed along these zones. Therefore As and Sb and other elements have formed in this area and are visible in alteration zones along the outcrop belt. The original rocks are mainly fine-grained siliciclastics that have been deposited in a relatively closed basin in central Iran within an intracontinental basin (Mazlomi, 1992). The source of As (210 to 26x104 ppm) in this contaminated area was high (Ghassemzadeh et al., 2003). For this study we selected P. australis for examination of As and Sb hyper-accumulation and root stabilization. The physiological and morphological attributes of the root systems are important for the uptake of metals from soil (Taize and Zeiger, 1998) and specially for root stabilization program (EPA, 2003). Phragmites australis produces vertical and horizontal rhizomes with developing roots that provide a great uptake surface for absorption of heavy metals such as As and Sb (Armstrong, 1979).
The aims of this study were two fold to evaluate As and Sb uptake in P. australis to identify suitable agent for rhizofiltration of As, Sb contaminated sites and to measure phosphorous (P), Iron (Fe) and antioxidant enzyme activities like catalase (CAT) and ascorbate peroxidase (APX) and guaicol peroxidase (GPX) in root system for understanding effects of interaction these two toxic metals on metabolic statues. These results could be helpful to introduce the new hyperaccumulation species for the As, Sb remediation in contaminated Chelpo region and similar areas.
MATERIALS AND METHODS
Site and sampling: The site of study area was 56 km north Kashmar in Khorasan province of Iran (Chelpo). The source of arsenic antimony is from Kohsorkh Mountain, 35°28’ latitude and 58°28’ longitude. Five sampling sites with high geogenic source of arsenic levels were chosen for this study. Control plants collected from non contaminated area. Samples of Phragmites australis and soil close to the roots have been collected with 100 m distances at the end of September 2006.
Preparation plant samples: In the laboratory, Plant samples separated into root, rhizome, stem and leave. Then they washed thoroughly under tap water, rinsed with distilled water 3 times and then dried in an oven at 70°C for 12 h. Air dried samples (0.2 g root and rhizome) were digested with nitric acid (Environmental express, Mt. pleasant, S.C.) using the USEPA method 3050A (http://www.epa.gov/.). The digestion temperature was controlled at approximately 80-100°C to avoid As volatilization (Jinasada et al., 1997).
Arsenic, antimony, iron and phosphorous determination: Determination of total Arsenic, Antimony and Iron (Fe) contents in the randomly triplicate samples from different tissues (root, rhizome, stem and leave) of P. australis was done using GFA-4B graphite furnace atomic absorption spectrometer (Shimadzu A-A-670G). Phosphorous was determined using Molybdate ammonium colorimetric method (American Public Health Association, 1995). Via this method blue complex is made from combination of sample phosphorous and additive Molybdate ammonium that intense of this blue color is depending of phosphorous content of samples. At the end Absorbance was read at 730 nm by UV-Vis spectrophotometer (Diode array WPA S 2000) and using standard curve phosphorous determine (μg mg–1) in plant samples.
Enzyme extraction and measurements: Fresh tissues (1 g) were homogenized at 4°C with 5 mL Triss hydrochloric acid buffer 0.05 M (pH 7.5). The homogenate was centrifuged at (14,000 x g) at 4°C for 20 min (Benavides et al., 2000). This supernatant was used for enzyme determination.
Enzyme determination: Catalase activity was assayed by a UV-Vis spectrophotometer (UV-120U UV-Vis recording spectrophotometer, SHIMADZU, Japan) according to the method of Chen et al. (2000). One unit of enzyme activity was defined as the amount of enzyme that reduced 50% of the H2O2 in 60 sec at 25°C (Kong et al., 1999). Also Guaicol peroxidase (GPX) and Ascorbate peroxidase (APX) activity were assayed UV-Vis spectrophotometer according to the methods of Bergmeyer (1974) and Koricheva et al. (1997), respectively. One unit of enzymes activity was defined as the amount of enzyme that catalyzed one micromole of the H2O2 per minute at 25°C (ε = 26.6 mM–1 cm–1) (Kong et al., 1999).
Statistical analysis: Statistical analysis was performed using SPSS software. A least significant test was employed for comparison of metabolic changes at three probability levels (p≤0.01 and p≤0.05). All results were expressed as mean, followed by corresponding standard deviation where n = 6.
RESULTS
Arsenic and antimony accumulation: Maximum and minimum mean of accumulated As in shoot system were 5.88 and 3.12 μg L–1 while for root were 33.73 and 24.95 μg L–1, respectively. Arsenic accumulation in root is significantly higher than shoot (p<0.05) (Fig. 1). These concentrations of As in the root and rhizomes are 0.5 to 84.5 μg L–1 and for rhizomes ranged 18.90 to 73 μg L–1. Arsenic mean for root and rhizomes are 8.97 and 5.90 μg L–1, respectively (Table 1, 2). Root and rhizomes Sb concentrations ranged 0.2 to 16.20 μg L–1 and 0.2 to 10.20 μg L–1, respectively. Antimony mean and median were 3.77-1.70 and 2.8-1.20 μg L–1, respectively.
Table 1 and 2 showed there was being maximum and minimum As accumulation in roots of first and second sample collection respectively. Also there were maximum and minimum Sb statues in roots of fifth and first sample sites respectively. Also Table 1 and 2 showed that maximum and minimum mean of As and Sb in rhizomes was on second and fist sample collection respectively.
Root and rhizome As/Sb ratio ranged from 35 to 194 and 10.42 to 99.9, respectively. Maximum and minimum As/Sb ratio is 429 in station 5 and 289.5 in station 2, respectively. As/Sb ratio analysis in different sample collection showed on Table 3.
Arsenic antimony interactions
Phosphorous changes: In present study average range of root phosphorous was between 0.14 μg mg–1 in first site to 0.29 μg mg–1 in fifth site. For rhizomes this was between 0.14 μg mg–1 in second site to 0.58 μg mg–1 in first site. Means of root and rhizome of P contents in control plants was 0.43 and 0.44 μg mg–1, respectively. Thus phosphorous contents of contaminated root system (root and rhizomes) decreased significantly in comparison to control plants (p<0.05).
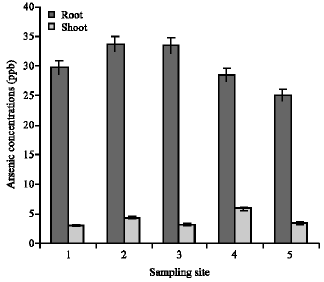 | |
| Fig. 1: | Arsenic concentrations in shoots and roots of P. australis in different sampling sites. Data represents mean±SD (n = 6) |
| Table 1: | Parametric presentation of Arsenic (As) and Antimony (Sb) in the roots of P. australis |
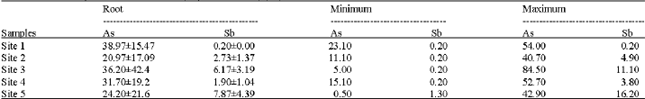 | |
| Mean correspond±SD of six replicates | |
| Table 2: | Parametric presentation of arsenic and antimony in the rhizomes of P. australis |
 | |
| Mean correspond±SD of six replicates | |
| Table 3: | Parametric presentation of arsenic/antimony ratio in the roots and rhizomes of P. australis |
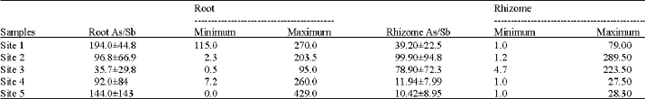 | |
| Mean correspond±SD of six replicates | |
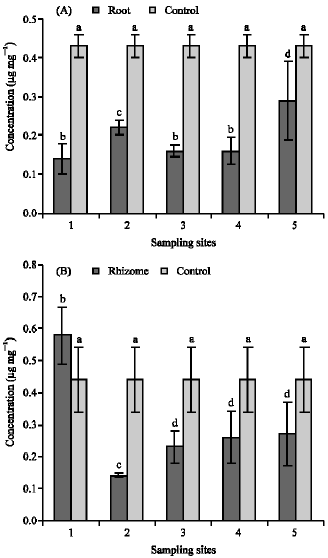 | |
| Fig. 2: | Phosphorous concentrations in root (A) and rhizomes (B) of P. australis in different sampling sites. Bars with the same letter are not significantly different among sampling sites (p<0.05). Data represents mean±SD |
In contaminated rhizomes maximum and minimum of P concentration were in first and second sampling site respectively. Significantly decreasing P in shoots and roots of contaminated plants in comparison to control plants is shown in Fig. 2.
Iron changes: Average range of the Iron contents was between 6.8 ppm in second site to 10.52 ppm in first site.
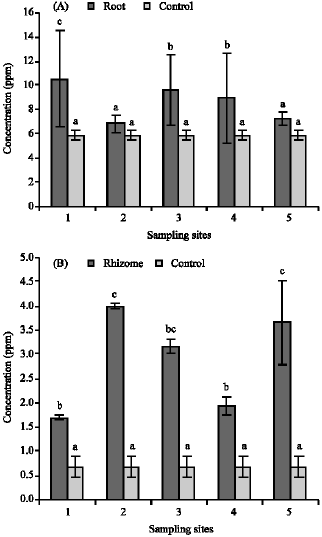 | |
| Fig. 3: | Iron concentrations in root (A) and rhizome (B) of P. australis in different sampling sites. Bars with the same letter are not significantly different among sampling sites (p<0.01). Data represents mean±SD |
While in rhizomes mean range was between 1.7 ppm in first site to 3.64 ppm in fifth site. Means of root and rhizome of Fe contents in control plants was 5.9 and 0.67 ppm, respectively. Thus Fe accumulation in root system (root and rhizomes) of contaminated plants was significantly higher than control plants. Also Fe concentration in reed root was significantly higher than rhizomes (p<0.01). Significantly enhancing Fe levels in roots of contaminated plants in comparison to control plants is shown in Fig. 3.
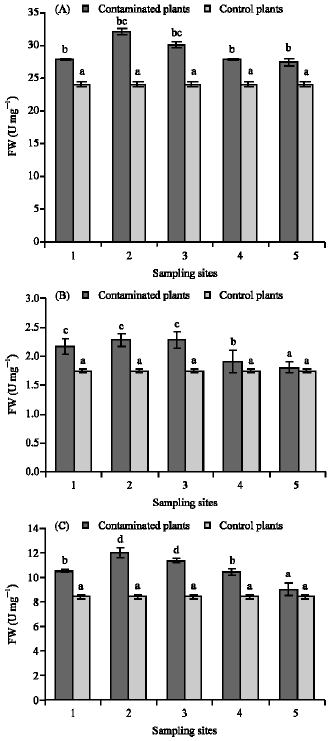 | |
| Fig. 4: | Catalase (A), Ascorbate peroxidase (B) and Guaicol peroxidase (C) roots of P. australis in different sampling sites. Bars with the same letter are not significantly different among sampling sites (p<0.01). Data represents mean±SD |
Enzymatic changes: Average range of CAT activity in contaminated root was between 27.4 in fifth sites to 29.3 U mg–1 FW (enzyme Unit/mg Fresh Weight) in second sampling site while mean value of catalase activity in control plant was 25 U mg–1 FW. Mean value of APX activity in control root was 1.75 U mg–1 FW while average range of APX activity in contaminated roots were 1.81 U mg–1 FW in fifth sampling site to 2.45 U mg–1 FW in second site. Also GPX activity ranges in contaminated roots were 9.71 U mg–1 FW in fifth site to 11.36 U mg–1 FW in second sampling site while mean value of GPX activity in control root was 8.20 U mg–1 FW. The enzymatic activities range in root system (root and rhizome) of contaminated plants was significantly higher than control plants (p<0.05). Significantly enhancing enzymatic responses (CAT, APX and GPX) in roots of contaminated plants in comparison to control plants is shown in Fig. 4.
DISCUSSION
Antimony and arsenic may co-occur in ground water and surface water. However considering their average crustal abundance of 1.8 μg g–1 for As and 0.2 μg g–1 for Sb (Smith and Huyck, 1999). Arsenic also in the study area (sampling site 1, 2, 3, 4 and 5) ranged from 37.4 to 376 μg L–1 and antimony was 11.4 to 127.1 μg L–1 (Ghassemzadeh et al., 2006). In present study we found that in all sampling sites with different amount of arsenic, P. australis consistently accumulated arsenic in the roots more than in the shoots. Only small amount of As translocated to shoot system and rhizofiltration is a process for removing As and Sb by P. australis. In this technique plants transfer only small amount of As into shoot in order to prevent As movement into wild life and food chain (Porter and Peterson, 1975).
Craw et al. (2007) found that Sb contents of all examined plants (shrubs, grasses and reeds) are about one thousandth of that of As. We also found very small As accumulation in reed shoot system, thus for the examination of As and Sb interaction, we only measured Sb content in the root system (root and rhizome). Antimony contents of root and rhizomes are about one 100th and one 48th of As respectively, reflecting the As/Sb ratio of the substrates. This ratio was higher in the roots than rhizomes. Craw et al. (2007) reported that decrease Sb than As accumulation recommended that shrubs and grasses can cope with As phytostabilization of contaminated area.
In all sampling sites among the root systems, the roots accumulated more arsenic and antimony than rhizomes. Greater length and smaller diameter in roots will result in a greater surface area per unit of mass from roots and can increase the uptake of As in plants compared with those plants with a root mass having lower surface area. Another characteristic pivotal to increasing root surface area ensues from root hairs, is tubular extension of the specialized epidermal cells. The root hairs provide an increased capacity for absorption of soil ions and to be a lesser extent, soil water from the rhizopher (Taize and Zeiger, 1998). Thus significant increment of root As and Sb accumulation than rhizomes was reasonable.
Arsenic and Sb are often associated in the environment and so understanding interactions between these two elements with nutrients is necessary. Also there are many studies about exposure to As, not many study address exposure to Sb or the interaction effects of these two heavy metals on nutrients.
In present study P contents of contaminated roots and rhizomes was significantly decreased by increasing Sb and As respectively. Thus we found that there are antagonistic relationship between these two toxic heavy metals and P contents in contaminated root systems. The presence of certain elements and ions such as phosphate (Huysmans and Frankenberger, 1991) and antimony (Andrewes et al., 2000) may interfere to arsenic contents. Adriano (2001) declared that Phosphate ion plays a prominent role in anion-As interactions due to its physiochemical similarities to As. Moreover arsenate is thought to be taken up via the phosphate uptake system and may consequently interact with plant P nutrition (Meharg and Macnair, 1990).
In present study, Fe concentrations in root system significantly increased in all samples (p≤0.01). Porter and Peterson (1975) found a highly significant correlation (p≤0.001) between As and Fe in several As tolerant plants from different mine sites in the United Kingdom. The high concentration of Fe in the reed roots may be due to the presence of an iron plaque around the roots that detoxify pollutant and purify water (St-Cyr and Crowder, 1988; Wang and Peverly, 1996). This plaque may protect the roots from heavy metal toxicity, probably by co-precipitation or absorption of arsenic and antimony.
Common reed establishment may be locally enhanced by significantly decreased root phosphorous 1.79 and 1.18 times less than control root and rhizomes (p<0.05). Also by significantly increased iron contents 1.42 and 5.83 times more than control root and rhizomes (p<0.01). Moreover similar to the Fe and As accumulation model, more P was retained in the roots of the common reed to cope with increased arsenic concentration. The experiments of arsenic tolerant plants revealed that tolerant species had suppressed the phosphate arsenate high affinity uptake system and were less efficient in absorbing phosphate (Meharg et al., 1994). We postulate that the decreased plant P uptake, coupled with high P concentrations in the roots than rhizomes in P. australis demonstrates part of the As detoxification mechanism.
Arsenic is known to induce oxidative stress in plants by generating various Reactive Oxygen Species (ROS) (Tu and Ma, 2003) but there is not enough documents about Sb interaction with antioxidant enzymes. Tolerant species for minimizing the harmful effect of ROS have evolved an effective scavenging system composed of antioxidant molecules and antioxidative enzymes (Meharg, 1994). High activities measured for all four enzymatic components (APX, CAT and GPX) in P. australis upon As and Sb uptake. We evaluate maximum enzyme activities in second sampling site and minimum of its on fifth site. Thus these increments in all sampling site was increased with increments of As and Sb uptake. These enzymes quench ROS to a low level, thus enhancing the continuous accumulation of arsenic levels happened.
Arsenic and antimony accumulation with enhancing As/Sb ratio, significant changes in P and Fe and enzymatic responses in the root system showed that this species is As and Sb tolerant and stabilize these two toxic metals in the roots. Thus this species could be used as rhizofiltration agent on this site and similar contaminated area.
ACKNOWLEDGMENTS
This study was supported by the assistance of scientific research of Ferdowsi University of Mashhad. The first author gratefully acknowledges Professor Jerry Schnoor who reviewed the first draft and made some suggestions to improve the manuscript.
REFERENCES
- Ali, N.A., M.P. Bernal and M. Ater, 2002. Tolerance and bioaccumulation of copper in Phragmites australis roots and Zea mays. Plant Soil, 239: 103-111.
Direct Link - Andrewes, P., W. Cullen and E. Polishchuk, 2000. Arsenic and antimony biomethylation by Scopulariopsis brevicaulis: Interaction of arsenic and antimony compounds. Environ. Sci. Technol., 34: 2249-2253.
Direct Link - Benavides, M.P., P.L. Marconey, S.M. Gallego, M.E. Comba and M.I. Tamaro, 2000. Relationship between antioxidant defence systems and salt tolerance in Solano tobarasum. Aust. J. Plant Physiol., 27: 273-278.
Direct Link - Chen, Y., X.D. Cao, Y. Lu and X.R. Wang, 2000. Effects of rare earth metal ions and their EDTA complexes on antioxidant enzymes of fish liver. Bull. Environ. Contam. Toxicol., 65: 357-365.
CrossRefDirect Link - Chu, W.K., M.H. Wong and J. Zhang, 2006. Accumulation, distribution and transformation DDT. PCBs by Phragmites australis and Oryza sativum. Environ. Geochem., 28: 169-181.
Direct Link - Craw, D., C. Rufat, L. Haffert and L. Paterson, 2007. Plant colonization and Arsenic uptake on high arsenic mine waster, New Zealand. Water Air Soil Pollut., 179: 351-364.
Direct Link - Ederli, L., L. Reale, F. Ferranti and S. Pasqualini, 2004. Responses induced by high concentration of cadmium in Phragmites australis roots. Physiol. Plant, 121: 66-74.
Direct Link - Fitz, W.J. and W.W. Wenzel, 2002. Arsenic transformations in the soil-rhizosphere-plant system: Fundamentals and potential application to phytoremediation. J. Biotechnol., 99: 259-278.
CrossRefPubMedDirect Link - Gebel, T., 2000. Confounding variable in the environmental toxicology of arsenic. Toxicology, 144: 155-162.
Direct Link - Ghassemzadeh, F., M.H. Arbab Zavar and M. Geoffrey, 2006. Arsenic and antimony in drinking water in Kohsorkh area, Northeast Iran possible risks for health. J. Applied Sci., 6: 2705-2714.
Direct Link - Fereshteh, G., B. Yassaman, A.M.M. Reza, A. Zavar and M. Hossein, 2007. Phytoremediation of arsenic by macroalga: Implication in natural contaminated water, Northeast Iran. J. Applied Sci., 7: 1614-1619.
CrossRefDirect Link - Hozhina, E.I., A.A. Khramov, P.A. Gerasimov and A.A. Kumarkov, 2001. Uptake of heavy metals, arsenic and antimony by aquatic plants in the vicinity of ore mining and processing industries. J. Geochem. Explor, 74: 153-162.
Direct Link - Huysmans, K.D. and W.T. Frankenberger, 1991. Evolution of trimethylarsine by a Penicillium sp. Isolated from agricultural evaporation pond water. Sci. Total Environ., 105: 13-28.
CrossRefDirect Link - Jinadasa, K.B.P.N., P.J. Milham, C.A. Hawkins, P.S. Cornish, P.A. Williams, C.J. Kaldor and J.P. Conroy, 1997. Survey of cadmium levels in vegetables and soils of greater Sydney, Australia. J. Environ. Qual., 26: 924-933.
CrossRefDirect Link - Kong, F.X., W. Hu, S.Y. Chao, W.L. Sang and L.S. Wang, 1999. Physiological responses of the lichen Xanthoparmelia mexicana to oxidative stress of SO2. Environ. Exp. Bot., 42: 201-209.
CrossRefDirect Link - Koricheva, J., S. Roy, J.A. Vranjic, E. Haukioja, P.R. Hughes and O. Hanninen, 1997. Antioxidant responses to simulated acid rain and heavy metal deposition in birch seedlings. Environ. Pollut., 95: 249-258.
CrossRefDirect Link - Lee, B.H. and M. Scolz, 2007. What is the role of Phragmites australis in experimental constructed wetland filters treating urban run off. Ecol. Eng., 29: 87-95.
Direct Link - Meharg, A.A. and M.R. Macnair, 1990. An altered phosphate uptake system in arsenate-tolerant Holcus lanatus L. New Phytol., 116: 29-35.
CrossRefDirect Link - Meharg, A.A., 1994. Integrated tolerance mechanisms: Constitutive and adaptive plant responses to elevated metal concentrations in the environment. Plant Cell Environ., 17: 989-993.
CrossRefDirect Link - Meharg, A.A., J. Bailey, K. Breadmore and M.R. Macnair, 1994. Biomass allocation, phosphorous nutrition and vesicular arbuscular mycorrhiza infection in clones of Yorkshire Fog, Holcus lanatus L. (Poaceae) that differ in their phosphate uptake kinetics and tolerance to arsenate. Plant Soil, 160: 11-20.
Direct Link - Porter, E.K. and P.J. Peterson, 1975. Arsenic accumulation by plants on mine waste (United Kingdom). Sci. Total Environ., 4: 365-371.
CrossRefDirect Link - Smedley, P.L. and D.G. Kinniburgh, 2002. A review of the source, behavior and distribution of arsenic in natural waters. Applied Geochem., 17: 517-568.
Direct Link - St-Cyr, L. and A.A. Crowder, 1988. Iron oxide deposits oh the roots of Phragmites australis related to the iron bound to carbonates in the soil. J. Plant Nutr., 11: 1253-1261.
CrossRefDirect Link - Southichak, B., K. Nakano, M. Nomura, N. Chiba and O. Nishimura, 2006. Phragmites australis: A novel biosorbent for the removal of heavy metals from aqueous solution. Water Res., 40: 2295-2302.
Direct Link - Suresh, B. and G.A. Ravishankar, 2004. Phytoremediation-A novel and promising approach for environmental clean-up. Crit. Rev. Biotechnol., 24: 97-124.
CrossRefPubMedDirect Link - Tu, S. and L.Q. Ma, 2003. Interactive effects of pH, arsenic and phosphorus on uptake of as and P and growth of the arsenic hyperaccumulator Pteris vittata L. under hydroponic conditions. Environ. Exp. Bot., 50: 243-251.
Direct Link - Volesky, B., 2001. Detoxification of metal-bearing effluents: Biosorption for the next century. Hydrometallurgy, 59: 203-216.
CrossRefDirect Link - Wang, T. and J.H. Peverly, 1996. Oxidation states and fractionation of plaque iron on roots of common reeds. Soil Sci. Soc. Am. J., 60: 323-329.
CrossRefDirect Link