
Boukare Zeba
Universite de Ouagadougou, UFR/SVT, 03 BP7021 0uagadougou (13 Burkina Faso
Jacques Simpore
Centre medical Saint Camille, 01 BP 364 Ouagadougou Burkina Faso
Odile Germaine Nacoulma
Universite de Ouagadougou, UFR/SVT, 03 BP7021 0uagadougou (13 Burkina Faso
Journal of Applied Sciences
Year: 2007 | Volume: 7 | Issue: 1 | Page No.: 98-102







ABSTRACT
β-lactamase production ways and inhibition patterns were investigated in cell suspensions of clinical isolates. The purpose of this research was prior to investigate the different β-lactamase molecular classes occurring in Burkina Faso owing to the local practice of β-lactam antibiotics. The use of specific inhibitors enabled to draw up an inhibition profile and consequently to assign an enzyme to accurate molecular class of β-lactamase. At the same time, β-lactamase expression ways were explored and correlated to the inhibition profiles. The current results pointed out two main groups of organisms. The first group of isolates secretes β-lactamases mainly by inducible way. Interestingly this bacterial group carries molecular class C of β-lactamase. The second group of isolates uses partial inducible and partial constitutive way for enzyme expression. These bacteria mostly carry molecular class A of β-lactamases. The sole exception encountered during these investigations is an organism, expressing exclusively by the constitutive way an enzyme that is found to belong to molecular class B of β-lactamases.
PDF Abstract XML References Citation
How to cite this article
Boukare Zeba, Jacques Simpore and Odile Germaine Nacoulma, 2007. β-lactamase Molecular Classes Linked to Production Ways from Clinical Isolates in Burkina Faso/West Africa. Journal of Applied Sciences, 7: 98-102.
DOI: 10.3923/jas.2007.98.102
URL: https://scialert.net/abstract/?doi=jas.2007.98.102
DOI: 10.3923/jas.2007.98.102
URL: https://scialert.net/abstract/?doi=jas.2007.98.102
INTRODUCTION
β-lactamases received classification number EC 3.5.2.6 from the international committee of enzymes (Webb, 1984). These enzymes destroy β-lactam antibiotics by hydrolysing them (Abraham and Chain, 1940), thereby rendering the antimicrobial ineffective (Reaction1). Therefore, β-lactamases are the main responsibles for bacterial resistance to β-lactam rings (Frère, 1995; Medeiros, 1997; Massova and Mobashery, 1998). They were grouped into for molecular classes (A, B, C and D) on the basis of amino acid sequences and into subgroups according substrate/inhibitor profiles (Bush et al., 1995).
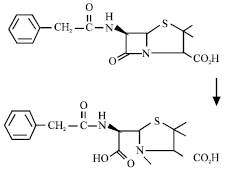 |
| Reaction 1 Hydrolysis of benzylpenicillin by a β-lactamase (Bush, 1988) |
Class A enzymes were formerly designed penicillinase to refer to their preference for penicillins in term of good penicillin-hydrolysing activity. Likewise Class C.
β-lactamases were characterized as cephalosporinases on the basis of their high hydrolysis rates for cephalosporins (Fleming et al., 1963). Class A β-lactamases includes most enzymes being chromosomal or plasmid mediated (Bush, 1989; Boissinot and Levesque, 1990; Perilli et al., 1991). Class C β-lactamases includes chromosomal cephalosporinases generally carried by Gram¯bacteria. Class D β-lactamases were described in the late 1980s (Ouellette et al., 1987; Huovinen et al., 1988). Class D enzymes must be considered as penicillinases able to inactivate Oxacillin with high hydrolysis rates. This efficiency in Oxacillin hydrolysis explains their designation as Oxacillinase. The class B β-lactamases represent the only metallo- β-lactamases that are further step forward in the evolution of antibiotic resistance and are now a source of increasing clinical concern. They are zinc-dependent enzymes that possess the ability to inactivate most β-lactam antibiotics, including the strongest as carbapenems (Bush, 1998; Zeba et al., 2005).The strategy for classification of β-lactamases often used specific inhibitors, because a single inhibitor is not effective for all the different enzymes (Bush, 1998; Bush and Sykes, 1983).
Present study intends to analyse β-lactamase production ways in clinical isolates with particular reference to inhibition profiles related to molecular classes occurring in Burkina Faso. It should be noted that only β-lactamases produced by Gram¬ bacteria were considered.
MATERIALS AND METHODS
acteria and culture conditions: Gram¯bacteria from various sources (faeces, urine, wound, blood) were collected from 2000 to 2005 at Centre Médical Saint Camille (SCMC). SCMC is a central institution for care and medical analysis in the capital city-Ouagadougou. Twenty seven isolates gave good hydrolytic activity on nitrocefin and were retained for further investigations. All the isolates were identified by Api 20 and Vitek2 systems ((BioMerieux France). The identified organisms were grown aerobically by shaking (250 rpm) at 37°C in Luria-Bertani (LB) liquid medium.
β-lactamase characterization: Cultures of 10 mL of each isolate were used to prepare the crude extracts of enzymes. Cell suspensions were harvested by centrifugation at 8546 g and 4°C during 15 mn. The pellet was suspended in 500 μL of sodium phosphate buffer 100 mM pH 7 and the periplasmic content was released by subjecting cells to five cycles of freeze-thaw (Simpson and James, 1982). Then cell debris were discarded again by centrifugation at 14000 g and 4°C for 15 mn. The supernatant generally contains the β-lactamase activity. This latter was stored at 4°C for further use.
Spectrophotometric β-lactamase assays: β-lactamase activity was measured at 25°C in 50 mM sodium phosphate buffer pH 7. The reaction was recorded on Uvikon 923 double beam UV/VIS spectrophotometer at 482 nm to monitor the nitrocefin (substrate) hydrolysis.
β-lactamase basal level determination and induction assays: Overnight preculture (600 μL) was used to inoculate 20 mL of Luria-Bertani liquid medium and the latter was grown for 1 h at 37°C on a rotating shaker. Then the culture was stopped and divided into 2 volumes of 10 mL. The 2 contents were grown an additional 4 h with an inducer in one the mediums, the second medium (without inducer), being a control. It should be noted that imipenem was used as inducer. The β-lactamase crude extracts from the 2 mediums were prepared as indicated above. The β-lactamase activity was tested from the 2 extracts using the chromogenic β-lactam nitrocefine as substrate. If the initial velocity (V0) is the same on the 2 extracts, the β-lactamase expression was considered as being constitutive. When the hydrolytic activity was observed only on induced extract, it was concluded that the activity was 100% inducible. Finally when the 2 extracts were both active but with at least doubled hydrolysis rate from induced assay compared to control, the β-lactamase expression was considered to be partial inducible and partial constitutive.
Inhibition of β-lactamase activities: Inhibition patterns and then, molecular classes characterization were achieved, with the help of specific inhibitors. Inhibition by suicide inactivator as clavulanic acid was used to identify class A β-lactamase, while inhibition by cloxacillin or monobactam aztreonam allowed the class C assignment. The metal chelators such as EDTA (Bush et al., 1995) or dipicolinic acid easily assigned to class B enzymes. Class D enzymes were reported to be inhibited by Chloride ions (Philippon et al., 1983). Therefore, Cl¯ was used to investigate class D enzymes. All the inhibition reactions were performed after 5 mn preincubation at 25°C of ongoing inhibitor with crude active extract. The total volume of reaction mixture was 500 μL containing 5 μL of crude extract, 100 μM of nitrocefine as substrate in final concentration and 1 mM of inhibitor (final concentration). NaCl was used exceptionally at a final concentration of 100 mM.
RESULTS
β-lactamase production: From Gram¯ bacteria being investigated, two important representatives emerged with two major production types. β-lactamases from most isolates as Citrobacter freundii and Enterobacter cloacae were fully inducible. Comparatively Escherichia coli and Klebsiella pneumoniae seemed to have basal concentration of enzymes whose level can be doubled in the presence of an inducer. A single isolate identified to Chryseobacterium indologenes was the exception in that its β-lactamase expression was exclusively constitutive (Table 1).
β-lactamase inhibition : The use of β-lactamase inhibitors enabled to discriminate and to characterize many molecular classes (Table 2). In this experimental conditions, at the indicated concentration, clavulanic acid completely abolished β-lactamase activity in crude extracts of Escherichia coli and Klebsiella pneumoniae. In the same time, the complete inactivation of hydrolytic activity by cloxacillin was observed on crude extract of Citrobacter freundii and Enterobacter cloacae the main representatives of this group.
| Table 1: | β-lactamase production ways |
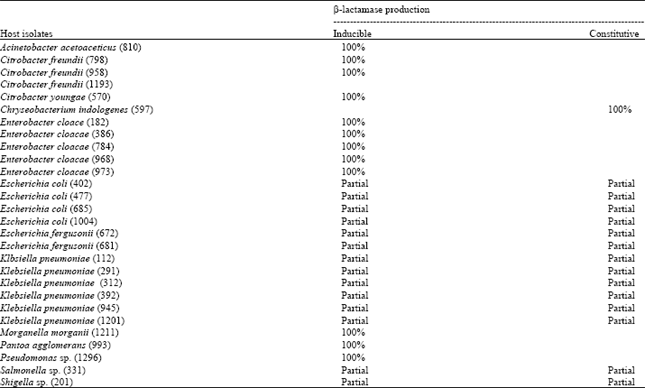 | |
| Table 2: | Inhibition patterns and molecular classes |
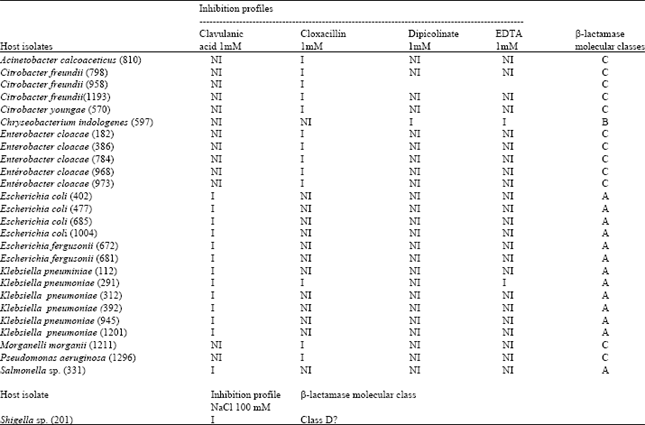 | |
| Legend from Table 2:1 = inhibited, NI = not inhibited | |
Likewise the activity of an extract from the single Chryseobacterium indologenes was strongly affected by EDTA or dipicolinic acid at the indicated concentration.
DISCUSSION
The most frequent clinical isolates are Escherichia species, Klebsiella pneumoniae Enterobacter cloacae and Citrobacter freundii. These clinical β-lactamase producing Gram¯ bacteria were investigated with particular reference to molecular classes occurring in Burkina Faso. Interestingly is the closest relation between expression way and inhibition profile, which suggests the molecular class of an enzyme. The majority of isolates possessing basal activity, whose level undergoes slight enhancement with an inducer, were found to express enzymes susceptible to clavulanate, implying molecular class A β-lactamases. The current results confirmed that Escherichia coli and Klebsiella pneumoniae are classical class A β-lactamase producers (Bush et al., 1995). However, the slight induction observed with these activities seemed to be unprecedented to our knowledge and remains intriguing. On the other hand, Most of the β-lactamases inhibited by cloxacillin and therefore belonging to molecular class C chromosomal enzymes, appeared to be carried by isolates expressing their enzymes exclusively by induction. These results underlined the previously mentioned data on class C β-lactamase in some members of the Enterobacteriaceae (Mahlen et al., 2003). The other case is the Chryseobacterium indologenes enzyme inhibited by metal chelators, thereby being molecular class B assignment. In contrast to the previous cases, this β-lactamase was exclusively expressed by constitutive way. These results are also consistent with reports that no evidence of β-lactamase inducibility was detected for Chryseobacterium indologenes isolates with imipenem or cefoxitin as an inducer (Bellais et al., 2000). On the other hand, no indubitable highlight of class D enzyme was obtained from our investigations. Although the net reduction of activity (50%) in the presence of NaCl 100 mM and its significant oxacillin hydrolysis rate, the Shigella sp. 201 extract failed to be clearly assigned to class D enzymes. Because in contrast to oxacillin, no confirmatory activity on cloxacillin was observed, unless this latter compound acted as inhibitor according previous findings (Matthew, 1978). In summary, this study successfully showed that at least three β-lactamase molecular classes (A, B and C) are occurring in our country. The molecular class D existence remains to be confirmed. Investigations are under way to find this missing link of bacterial β-lactamase-mediated resistance in Burkina Faso.
ACKNOWLEDGMENTS
This study was supported by the Coopération Universitaire au Développement (CUD) from Belgium through a project named Etude des enzymes de la résistance bactérienne aux antibiotiques. We greatly thank this institution. A great thank was also addressed to Dr Pierrette Melin of Service de Bactériologie du CHU de Liège for her useful technical contribution, especially in the bacterial identification.
REFERENCES
- Abraham, E.P. and E. Chain, 1940. An enzyme from bacteria able to destroy penicillin. Nature, 146: 837-837.
CrossRefDirect Link - Bellais, S., L. Poirel, S. Leotard, T. Naas and P. Nordmann, 2000. Genetic diversity of carbapenem-hydrolyzing metallo-β-lactamases from Chryseobacterium (Flavobacterium) indologenes. Antimicrob. Agents Chemother., 44: 3028-3034.
PubMedDirect Link - Boissinot, M. and R. Levesque, 1990. Nucleotide sequence of the PSE-4 carbenicillinase gene and correlations with Staphylococcus aureus PC1 β-lactamase crystal structure. J. Biol. Chem., 265: 1225-1230.
Direct Link - Bush, K., 1988. β-lactamase inhibitors from laboratory to clinic. Clin. Antimicrob. Chemother., 1: 109-123.
CrossRefDirect Link - Bush, K., G.A. Jacoby and A.A. Medeiros, 1995. A functional classification scheme for β-lactamases and its correlation with molecular structure. Antimicrob. Agents Chemother., 39: 1211-1233.
PubMedDirect Link - Fleming, P.C., M. Goldner and D.G. Glass, 1963. Observations on the nature, distribution and significance of cephalosporinase. Lancet, 1: 1399-1401.
CrossRefDirect Link - Frere, J.M., 1995. Beta-lactamases and bacterial resistance to antibiotics. Mol. Microbiol., 16: 385-395.
CrossRefDirect Link - Huovinen, P., S. Huovinen and G.A. Jacoby, 1988. Sequence of PSE-2 β-lactamase. Antimicrob. Agents Chemother., 32: 134-136.
PubMed - Mahlen, S.D., S.S. Morrow, B. Abdalhamid and N.D. Hanson, 2003. Analyses of ampC gene expression in Serratia marcescens reveal new regulatory properties. J. Antimicrob. Chemother., 51: 791-802.
Direct Link - Massova, I. and S. Mobashery, 1998. Kinship and diversification of bacterial penicillin-binding proteins and β-lactamases. Antimicrob. Agents Chemother., 42: 1-17.
Direct Link - Matthew, M., 1978. Properties of the β-lactamase specified by the Pseudomonas plasmid R151. FEMS Microbiol. Lett., 4: 241-244.
Direct Link - Medeiros, A.A., 1997. Evolution and dissemination of β-lactamases accelerated by generations of β-lactam antibiotics. Clin. Inf. Dis., 24: S19-S45.
CrossRefDirect Link - Ouellette, M., L. Bissonnette and P.H. Roy, 1987. Precise insertion of antibiotic resistance determinants into Tn21-like transposons: Nucleotride sequence of the OXA-1 β-lactamase gene. Proc. Natl. Acad. Sci. USA., 84: 7378-7382.
Direct Link - Perilli, M., N. Franceschini, B. Segatore, G. Amicosante and A. Oratore et al., 1991. Cloning and nucleotide sequencing of the gene encoding the β-lactamase from Citrobacter diversus. FEMS Micobiol. Lett., 83: 79-84.
Direct Link - Philippon, A.M., G.C. Paul and G.A. Jacoby, 1983. Properties of PSE-2 beta-lactamase and genetic basis for its production in Pseudomonas aeruginosa. Antimicrob. Agents Chemother., 24: 362-369.
CrossRefDirect Link - Simpson, I.N. and M. James, 1982. Comparison of routine techniques for cell breakage and release of β-lactamase activity. J. Antimicrob. Chemother., 9: 119-123.
CrossRefDirect Link - Zeba, B., J. Simpore, O. Nacoulma and J-M. Frere, 2005. Identification of metallo-β-lactamase from a clinical isolate at saint camille medical center of ouagadoudou, Burkina Faso. Afr. J. Biotechnol., 4: 286-288.
Direct Link