
C.K. Yap
Department of Biology, Faculty of Science, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
S.G. Tan
Department of Biology, Faculty of Science, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor, Malaysia
Journal of Applied Sciences
Year: 2007 | Volume: 7 | Issue: 20 | Page No.: 3111-3114







ABSTRACT
It had been documented through electrophoretic studies that allozyme polymorphisms of bivalve populations are affected by heavy metal stress. In the present laboratory study, by using P. viridis as a test organism, the results indicated that changes in the enzymes GOT, EST and ME were due to Zn stress which are complemented by reductions of filtration rate and condition index. However, it is not yet known for sure whether the enzymes GOT, EST and ME are inducing behavioural and other changes in P. viridis. This is because of the possible subtle interactions could occur between different environmental stresses.
PDF Abstract XML References Citation
How to cite this article
C.K. Yap and S.G. Tan, 2007. Changes of Allozymes (GOT, EST and ME) of Perna viridis Subjected to Zinc Stress: A Laboratory Study. Journal of Applied Sciences, 7: 3111-3114.
DOI: 10.3923/jas.2007.3111.3114
URL: https://scialert.net/abstract/?doi=jas.2007.3111.3114
DOI: 10.3923/jas.2007.3111.3114
URL: https://scialert.net/abstract/?doi=jas.2007.3111.3114
INTRODUCTION
Much of the advances in toxicology over the last two decades have been focused on the identification and characterisation of specific proteins expressed in response to toxic metals and xenobiotics (Viarengo, 1989; Lauwerys et al., 1995; Soni and Mehendale, 1998). It had been documented through electrophoretic studies that allozyme polymorphisms and heterogeneities of animal and plant populations and species are affected by environmental stress (Nevo et al., 1986; Ben-Shlomo and Nevo, 1988; Gillespie and Guttman, 1999). Recently, Yap et al. (2004) found a positive relationship between metal levels and allozyme polymorphisms in the soft tissues of the green-lipped mussel Perna viridis collected from contaminated and uncontaminated sites. However, it was argued that not only metals could affect allozyme polymorphisms of mussels since other types of pollutants could cause effects too. Therefore, the objectives of this study are to show if allozyme changes could be caused by a single pollutant stress-Zn contamination after subjecting a mussel population to Zn stress under laboratory conditions for a period of 28 days and to assess enzyme polymorphisms as reliable biomarkers after the mussels were exposed to Zn contamination under laboratory conditions.
MATERIALS AND METHODS
Individuals of P. viridis were collected from an unpolluted area (Pasir Panjang: 02°25’ N, 101°56’ E) of the west coast of Peninsular Malaysia on March 2000. After the shell surfaces were cleaned, all the mussels were transported to the laboratory at the Institute of Bioscience’s hatchery in the campus of Universiti Putra Malaysia in Serdang. During the acclimatization period, seawater was changed daily and the mussels were fed with monocultured alga Isochrysis galbana. Spawning was observed in most of the mussels after 3 days of acclimatization and they were immediately isolated after spawning. Mussels, which had released their spawn, were selected for this experiment. Only relatively similar sized (4-6 cm) mussels were chosen for study. The seawater used was filtered through a plankton net with 200 μm meshes to remove suspended particles (Yap et al., 2002a).
After the acclimatization period (at least 3 days), 36 mussels were exposed to a sublethal concentration of Zn (1.00 mg L–1) in an experimental aquarium while 36 mussels were put into a control aquarium. The concentrations of Zn chosen in this study were based on an earlier study conducted by Yap et al. (2003a). The seawater was changed once every two days and the mussels were fed with I. galbana during the change of seawater and metal solution. After 28 days of metal exposure, mussels exposed to 1.00 mg L–1 Zn had 58% (21 deaths) mortality (Table 1). One control with mussels without metal addition was run simultaneously and about 10% mortality (4 deaths) were observed in the control samples. Only live mussels were taken for metal and allozyme analyses.
The test seawater (10 L) was kept semi-static but constantly aerated and was held at room temperature (25-28°C), salinity at 25-26 ppt, dissolved oxygen at 6.9-7.4 mg L–1 (80-90%), pH 7.55-7.72, during the experimental period.
| Table 1: | Shell length (cm), filtration rate (FR) (L/h/mussel) [and FR reduction] and condition index (CI) (g cm–3) [and CI reduction] of P. viridis and the Zn concentration (μg g–1) in the soft tissues of mussels exposed to Zn (1.00 mg L–1) after 28 days |
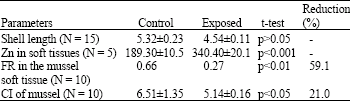 | |
The determinations of Filtration Rate (FR) (and rate of FR reduction) and Condition Index (CI) (and rate of CI reduction), were as described by Yap et al. (2003b) and Yap et al. (2002a), respectively.
For Zn analysis, the total soft tissues of the live mussels were pooled and 5 replicates were analysed; the procedure used was as described by Yap et al. (2002a). For the allozyme analysis, the muscle tissues were dissected and kept at -70°C until electrophoretic study. The horizontal starch gel electrophoresis procedure used followed that described by Yap et al. (2002b). Two buffer systems were used in this study. CA-7 was the buffer system described by Steiner and Joslyn (1979) while the 0.2 M Phosphate buffer system was as described by Shaw and Prasad (1970).
RESULTS AND DISCUSSION
In the mussels exposed to Zn after 28 days, changes in enzyme migrations were found for cathodally migrated glutamate oxaloacetate GOT(-), cathodally migrated esterase EST(-) and anodally migrated malic enzyme ME(+) (Fig. 1). The results were also complemented by reductions of CI (24%) and FR (59%) in the exposed mussels when compared to the control mussels. The Zn concentrations in the exposed mussels were found to be about 2 times higher than those in the control samples.
The mobilities of the 3 enzymes were clearly different between the exposed and the control mussels. This indicates that differential sensitivities between exposed and control mussels may lead to differences in the phenotypes of the two populations. The homeostatic mechanisms included maintenance of energy and acid base state and detoxification of the accumulated end products. Although only 3 enzymes were included in the present study, our results are well supported by reports in the literature. For example, the adaptation process had been suggested to involve changes in allozyme frequencies in mussels when they were exposed to metal stress (Yap et al., 2002a).
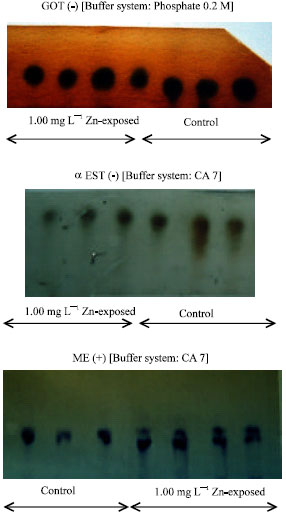 | |
| Fig. 1: | Comparison of the zymogram patterns of the enzymes GOT, EST and ME from control and Zn-exposed mussels after 28 days |
Recently, Yap et al. (2002a) found that the CI value from a metal-contaminated site is significantly (p<0.05) lower than from an uncontaminated one. The lower CI indicated that more stored energy was channeled to homeostatic mechanisms during a metal stress condition (Yap et al., 2002a). Homeostatic mechanisms such as somatic growth and energy devoted to detoxification and tissue repair had been disturbed (Viarengo et al., 1985). Even very low levels of pollutant exposure may be associated with various biological effects such as changes in activities of some enzymes (Lauwerys et al., 1995) and changes in metabolic fuels (Peters and Livingstone, 1996).
Metals are known to be associated with a metal binding protein (metallothionein) which may enter the lysosomes. The higher level of Zn found in the soft tissues of P. viridis after 28 days of exposure was indicative of the above matter. However, the 2-times higher Zn levels in the exposed mussels than in the control samples was low when compared to the 27 and 4 times for Cd and Pb exposures, respectively, after being exposed to 0.5 mg L–1 Cd and 1.50 mg L–1 Pb for 28 days (Yap et al., 2002a). This phenomenon could be due to partial regulation of Zn in the soft tissues of P. viridis which had been reported for this mussel species (Yap et al., 2003a; Phillips, 1985). The lysosomes represent an important cellular compartment for metal detoxification since they are able to accumulate large amounts of heavy metals in non-toxic forms (Viarengo and Nott, 1993). However, excessive concentrations of metals could cause alterations of structure, permeability and integrity of lysosomal membranes when the storage capacity is overloaded. According to Peakall (1992), the behaviour of an organism represents the final integrated result of a diversity of biochemical and physiological processes. In this study, we found behavioral changes in terms of reductions of CI and FR, which were reflected changes of GOT, ME and EST allozymes after Zn stress.
Perhaps, cellular damage occurred in the exposed mussels. Nicholson (1999) reported that P. viridis from contaminated sites in Hong Kong waters had significantly lower CI values than mussels collected from a reference (uncontaminated) site. He also found that the lower CI value in a contaminated site was linked to a dysfunction of the lysosomal system comprising of a digestive diverticula. In present results, the higher reductions of both CI and FR in the exposed mussels than in the control could be linked to a similar dysfunction that was manifested in the three enzymes.
There are thousands of genes that are selectively expressed in individual cells to yield the specific sets of proteins required to make the respective functioning cells. However, while the number of genes in an individual organism is static and fixed, the set of proteins that can be produced by an organism at one time or another throughout its life is not, this varies with age and development, cell and tissue types and in response to environmental stimuli (Moore, 2002). In our case, the environmental stimulus was the stress of Zn imposed onto the experimental mussels.
CONCLUSIONS
It is not yet known for sure whether the enzymes GOT, EST and ME are inducing behavioural and other changes in P. viridis although the present results showed changes in the three enzymes due to Zn stress under laboratory conditions complemented by reductions of FR and CI. However, since other heavy metals such as Cu, Pb, Cd and Hg, pesticides and hydrocarbon were also detected in the mussels, understanding of their effects on the enzyme changes is also needed. It remains an open question whether such effects are actually responsible for the field-observed phenomenon. Although a clear message is that there can be allozyme changes due to metal stress but one must be aware of the possible subtle interactions occurring between different environmental stresses. This question certainly needs much more studies in the future to resolves.
REFERENCES
- Lauwerys, R.P., A.B. Bernard, H. Roels and J.P. Buchet, 1995. Health risk assessment of long-term exposure to non-genotoxic chemicals: Application of biological indices. Toxicol. Lett., 77: 39-44.
Direct Link - Moore, M.N., 2002. Biocomplexity: The post-genome challenge in ecotoxicology. Aqua. Toxicol., 59: 1-15.
Direct Link - Nicholson, S., 1999. Cardiac and lysosomal responses to periodic copper in the mussel Perna viridis (Bivalvia: Mytiloidae). Mar. Pollut. Bull., 38: 1157-1162.
Direct Link - Peters, L.D. and D.R. Livingstone, 1996. Antioxidant enzyme activities in embriologic and early larval stages of turbot. J. Fish Biol., 49: 986-997.
CrossRefDirect Link - Soni, M.G. and H.M. Mehendale, 1998. Role of tissue repair in toxicologic interactions among hepatotoxic organics. Environ. Health Perspect., 106: 1307-1317.
Direct Link - Viarengo, A. and J.A. Nott, 1993. Mechanisms of heavy metal cation homeostasis in marine invertebrates. Comp. Biochem. Physiol., 104C: 355-372.
Direct Link - Yap, C.K., S.G. Tan, A. Ismail and H. Omar, 2002. Genetic variation of the green-lipped mussel Perna viridis (Linnaeus) from the west coast of Peninsular Malaysia. Zoological Studies, 41: 376-387.
Direct Link - Yap, C.K., A. Ismail, H. Omar and S.G. Tan, 2003. Accumulation, depuration and distribution of cadmium and zinc in the green-lipped mussel Perna viridis (Linnaeus) under Laboratory conditions. Hydrobiologia, 498: 151-160.
Direct Link - Yap, C.K., S.G. Tan, A. Ismail and H. Omar, 2004. Allozyme polymorphisms and heavy metal levels in the green-lipped mussel Perna viridis (Linnaeus) collected from contaminated and uncontaminated sites in Malaysia. Environ. Int., 30: 39-46.
CrossRefDirect Link