
Mahmoud Sanad Soliman
Department of Medicinal Chemistry, National Research Center, Dokki, Cairo, Egypt
Afaf El-Ansary
Department of Biochemistry, King Saud University, P.O. Box 22452, Riyadh,Zip Code 11495, Saudi Arabia
Journal of Applied Sciences
Year: 2007 | Volume: 7 | Issue: 19 | Page No.: 2881-2885







ABSTRACT
Amino acid profiles of control and Solanum nigrum, Ambrosia maritima, Thymelaea hirsute, Sinapis arvensis, Peganum haramala and Callistemon lanceolatus-treated Biomphalaria alexandrina snails were investigated in a trial to correlate the amino acid profile of treated snails to their previously reported molluscicidal and biological effects. Amino acid profiles of the snails were greatly manipulated with the treatment of dry powdered sublethal concentrations of the six studied plant molluscicides. The disturbed amino acid profiles of treated snails were discussed in relation to the decrease in snail's egg laying capacity, reduction of their compatibility for the development of the schistosome larvae and cercarial penetration of mammalian skin.
PDF Abstract XML References Citation
How to cite this article
Mahmoud Sanad Soliman and Afaf El-Ansary, 2007. Induced Changes in the Amino Acid Profile of Biomphalaria alexandrina Molluscan Host to Schistosoma mansoni Using Sublethal Concentrations of Selected Plant Molluscicides. Journal of Applied Sciences, 7: 2881-2885.
DOI: 10.3923/jas.2007.2881.2885
URL: https://scialert.net/abstract/?doi=jas.2007.2881.2885
DOI: 10.3923/jas.2007.2881.2885
URL: https://scialert.net/abstract/?doi=jas.2007.2881.2885
INTRODUCTION
Schistosomiasis, a dreadful disease caused by parasitic trematode worm in both humans as well as in animals is widespread in the world specially in developing countries (Engles et al., 2002). It is considered second only to malaria as a major target disease of the World Health Organization (Xiao et al., 2002).
Schistosomes as digenetic trematods have two hosts, a final mammalian hosts and a molluscan intermediate snail hosts. The freshwater snail Biomphalaria alexandrina (Mollusca; Gastropoda), is widely distributed in Egypt and acts as intermediate host of Schistosoma mansoni. Control of snail is regarded as one of the best preventive measures in controlling schistosomiasis (Madsen and Christiansen, 1992; WHO, 1993; Lardans and Dissous, 1998).
El-Ansary and Qurashy (1994) stated that the ability of the parasite to develop within snail host is correlated to the snail intrinsic biochemical composition rather than any regulatory immune response. Moreover, Thompson et al. (1991) reported that free living stages of schistosomes are completely dependent on the endogenous reserves acquired from their host in the previous parasitic stage. Cercariae for example, live on their endogenous glycogen and fatty acid stores that they build up while inside the snail host (Nabih et al., 1998).
It is well known that amino acids are among the Snail Conditioned Water (SCW) signals needed by schistosome miracidiae to identify their snail host species (Haberl et al., 2000). In fact, amino acid spectra released by freshwater snails varied not only among species, but also within the same species. This may play a critical role in snail-finding mechanisms by the parasite miracidia (Korner and Haas, 1998).
Reduction in free amino acid levels in haemolymph and tissues of mollusks during schistosome infection have been reported which proved their uptake by the developing parasite (Schnell et al., 1985). Certain amino acids (Glu, AsP, Gly and Gln) have stimulatory effects on the development of parasitic helminthes (Hata, 1994) because schistosomes have usual nutritional requirement for essential amino acids. Parasitic absorption and utilization likely explain much of the observed decrease.
Recently El-Ansary et al. (2001) could induce in vivo attenuation of schistosome cercariae using sublethal concentrations of selected plant molluscicides which include Solanum nigrum, Ambrosia maritime, Thymelaea hirsute, Sinapis arvensis, Peganum haramala and Callistemon lanceolatus. Tha pathogenicity of attenuated Schistosoma mansoni cercariae were evaluated (El-Ansary et al., 2003). Although the reduced number of attenuated cercariae released from the treated snails showed normal skin penetration rate, worm burden and egg count in the liver and intestine of mice infected with plant molluscicides-attenuated cercariae were remarkably lower compared to those infected with normal cercariae, showing reduced fecundity of worms developed from attenuated cercariae. Number and size of granulomatous reactions showed significant reduction in attenuated cercariae-infected mice. Moreover, AST, ALT, glycogen level and lipid peroxides were normalized in these mice when compared to control-infected group.
These informations initiated our interest to compare the amino acid profile of control and molluscicides-treated snails in a trial to find out if different amino acids are contributed to the previously reported remarkable reduction in snail compatibility to schistosome parasite (EL-Ansary et al., 2003) which could easily be correlated to the attenuation of cercariae released from molluscicide-treated snails.
MATERIALS AND METHODS
Field collected B. alexandrina (6-8 mm in diameter) were obtained from Abu-Rawash, Giza maintained in the laboratory in glass aerated aquaria, filled with dechlorinated water at 25±2°C, fed fresh lettuce leaves ad lib. and left for 45 days to ensure that they were free from infection. S. nigrum, A. maritime, T. hirsute, S. arvensis, P. haramala and C. lanceolatus are wild herbs. These plants were collected Egyptian country, dried and used as powder. The chemicals used were of analytical quality and purchased from Merck, Germany.
Molluscicide-treatment: B. alexandrina were exposed to LC10 values of the six studied plants which were obtained from the toxicity lines statistically calculated according to the method of Finney (1952). LC10 values were dissolved in dechlorinated water which have the snails for one week (El-Ansary et al., 2001).
Amino acid composition: High performance system Eppdrof-Germany LC 3000 Amino acid analyzer was used for determination of amino acid composition. The analysis was performed in the National Research Center (Unit of central services) Dokki, Cairo, Egypt.
Preparation of samples: Whole snails soft tissue was extracted and homogenized in 5 mL saline solution. Equal volume of 10% TCA added. Protein was precipitated as white amorphous precipitate, collected by centrifugation, washed with 5% TCA solution then with ether and absolute ethanol and dried in a vacuum desiccator. Twente five milligram protein was then hydrolyzed with 6 N HCL at 105°C for 24 h in a sealed tube (Bailey, 1967).
After cooling and filtering, the residue was washed with distilled water and the combined filtrates were completed to 25 mL in a volumetric flask. A portion of the filtrate (5 mL) was evaporated to dryness at room temperature in a desiccator under vacuum. The residue was dissolved in 5 mL buffer (0.2 N sodium citrate pH 2.2) and the solution was filtered through 0.22 μm membrane. Twenty microliters of the final filtrate were injected in the instrument capsule for quantitative determination of the amino acids (Ibrahim and El-Eraqy, 1996). The cationic exchange resins UL-tropac sodium and special programmed buffer system were used (Citrate buffer 0.2 N with three different pH at 3.2, 4.25 and 6.46 at flow rate 0.2 mL min-1). The effluent was met by a stream of ninhydrin reagent. The quantitative estimation of the amino acids depends on the colorimetric determination of the blue colour. Retention time and peak area were determined using computerized system for standard and samples. The relative percentage of each amino acid was calculated.
RESULTS AND DISCUSSION
Percentage changes of different amino acids in Biomphalaria alexandrina snails treated with the six used molluscicides are shown in Table 1 and Fig. 1(a-e). It can easily be noticed that amino acid profiles of the snails were greatly manipulated with molluscicides treatment. Aspartic, glutamic, tyrosine, lysine, arginine and proline are the most affected amino acids.
Amino acids are of critical importance in energy metabolism of mollusks, for example, glutamic acid represents the amino acid nitrogen pool for amino transferases to provide Krebs cycle intermediates. Moreover, amino acids are necessary for trematode-infected Biomphalaria species (Schnell et al., 1985; Nabih et al., 1990).
| Table 1: | The amino acids concentration μg mL–1 changes in Biomphalaria alexandrina snail treated with different plant molluscicides using amino acid analyzer |
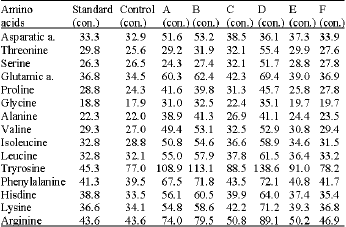 | |
| A- Peganum harmala; B- Callistemon lanceolatus; C- Sinapis arvensis D- Thymelaea hirsute; E- Ambrosia maritime; F- Solanum nigrum, Concentration (con) = μg mL–1 | |
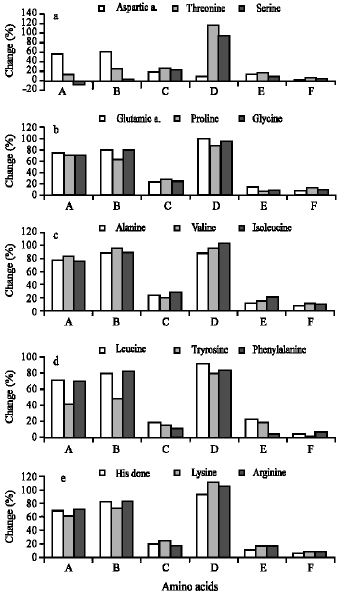 | |
| Fig. 1(a-e): | The percentage changes of different amino acids in Biomphalaria alexandrina snails treated with different plants molluscicides (A-F) relative to untreated snails (control), A- Peganum harmala; B- Callistemon lanceolatus; C- Sinapis arvensis; D- Thymelaea hirsute; E- Ambrosia maritime; F- Solanum nigrum |
The amino acid profile of control B. alexandrina snails reported in the present study is more or less similar to that reported in the Digestive Gland-Gonad complex (DGG) of Biomphalaria glabrata (Ponder et al., 2004). Qualitative analysis revealed the presence of histidine, lysine, serine, alanine, valine and isoleucine or leucine in both samples with non significant decrease in Lysine and valine in Echinostoma caproni- infected Biomphalaria glabrata. Altered amino acid spectra recorded in the present study may lead to abnormal SCW signals which in turn could disturb the snail-finding mechanisms by schistosome miracidiae (Korner and Haas, 1998).
Boehmler et al. (1996) proved that poly L-lysine affecting behaviour of hemocytes of Biomphalaria glabrata on the parasite. The cells showed minimal spreading, moved significantly faster and formed aggregates. In the present study, while S. nigrum and A. maritima show only slight alteration in lysine level, the other four plants P. harmala, S. arvensis, C. lanceolatus and T. hirsuta were more effective with T. hirsuta showed the lowest effect compared to the other three plant molluscicides. Induced lysine levels is of critical importance in inducing parasite killing by hemocytes of molluscicides-treated snails. The effective plants could have immunostimulatory effect through lysine. This could be used to explain the decrease in snail compatibility previously recorded by El-Ansary et al. (2000a, b and 2001) as reduction in the mean total number of cercariae shed/each Biomphalaria alexandrina snail treated with the same molluscicides. This could be easily correlated to the induced levels of aspartic acid which was recorded as effective phagostimulant in Biomphalaria glabrata (Thomas et al., 1989).
Infection by larval trematodes, often causes a cessation of egg production of molluscan intermediate host and this is referred to as parasitic castration. Manager et al. (1996) and Bai et al. (1997) attributed this to the depression of biogenic monoamines (Dopamine) and the inhibition of phenol oxidase activity as an enzyme playing a major role in egg laying capacity of snails. El-Ansary et al. (2003) showed that sublethal concentrations of the plant molluscicides used in the present study were effective in reducing the fecundity of the treated snails. This could find support in the present study, since phenyl alanine as precursor for dopamine was remarkably higher in S. nigrum, A. maritime, T. hirsute, S. arvensis and C. lanceolatus-treated snails which confirmed the impairment of dopamine synthesis. Moreover, in the present study, increase of L-tyrosine (phenol oxidase substrate) levels in tissues of plant-treated snails could be functionally linked to the significant reduction in fecundity of treated snails (El-Ansary et al., 2003). This in turn provides strong support for the crucial role of phenol oxidase in normal egg production. This suggestion could find support in the study of Bai et al. (1997) which assessed the effects of larval infection on L-tyrosine (PO substrate) levels in the albumin gland and ovotestis and revealed a significant increase in the level of L-tyrosine in both organs in castrated infected snails.
Serine was only elevated in S. arvensis-treated snails while alanine was elevated with S. arvensis, C. lanceolatus and Peganum haramala but not with S. nigrum and A. maritime. This could be explained on the basis that the latter two plants reduces the concentration of pyruvate through inhibiting the glycolytic flux. Pyruvate is the precursor of alanine through alanine aminotransferase enzyme (ALT). On the other hand, glutamate as a second product of ALT activity was not elevated in S. nigrum and A. maritima-treated snails. This could be easily attributed to the fact that glutamate is an amino group donor in many other transaminases reactions.
Some amino acids are anaplerotic and contribute to gluconeogenesis because all or part of their carbon skeletons enter the citric acid cycle after deamination or transamination e.g., alanine, cysteine, glycine, serine, threonine for pyruvate, arginine, histidine, glutamine and proline are convertible to α-ketoglutarate via glutamate; isoleucine and valine are converted to succinyle CoA, in addition to tyrosine and phenyl alanine which form fumarate. Elevation of glutamic, aspartic, glycine, valine, proline, isoleucine, histidine and arginine as glucogenic amino acids in Thymelaea hirsute, Sinapis arvensis, Peganum haramala and Callistemon lanceolatus treated snails proved the impairment of gluconeogenesis.
It has previously been postulated that L-arginine emitted by penetrating cercariae serves as an intraspecific signal guiding other cercariae to the penetration site (Ingram et al., 2003). Moreover, Haas et al. (2002) recorded that attachment of Schistosom mansoni cercariae to mammalian skin is specifically stimulated by L- arginine which acts as pheromone which could guide cercariae towards common penetration sites. In addition, Schistosomula (transformed cercariae) could use L- arginine to produce Nitric Oxide (NO) for blood vessel dilation during their migration in the host. Based on these informations, the remarkable increase of arginine in the treated snail's tissues, could lead to the development of intramolluscan parasitic stages and the release of cercariae rich with arginine and hence show unaltered penetration rate to mammalian skin. This could supported the previous results of El-Ansary et al. (2003) who reported that snails treated by sublethal concentrations of the same plant molluscicides showed normal penetration rate in spite of their attenuation and decrease of their pathogenicity to the mammalian hosts.
In conclusion, treatment of Biomphalaria alexandrina snails with sublethal concentration of Solanum nigrum, Ambrosia maritima, Thymelaea hirsute, Sinapis arvensis, Peganum haramala and Callistemon lanceolatus was effective in altering the amino acid profile of this snail species which could be contributed to the impairment of snail egg laying capacity, snail-schistosome miracidiae finding mechanisms and immune response of the molluscan hosts but has no effect on the mammalian skin penetration rate by schistosome cercariae.
REFERENCES
- Bai, G., L.A. Johnston, C.O. Watson and T.P. Yoshino, 1997. Phenoloxidase activity in the reproductive system of Biomphalaria glabrata: Role in egg production and effect of schistosome infection. J. Parasitol., 83: 852-858.
Direct Link - Boehmler, A.M., S.E. Fryer and C.J. Bayne, 1996. Killing of Schistosoma mansoni sporocysts by Biomphalaria glabrata hemolymph in vitro: Alteration of hemocyte behavior after poly-L-lysine treatment of plastic and the kinetics of killing by different host strains. J. Parasitol., 82: 332-335.
Direct Link - El-Ansary, A., E.M. Sammour and A.M. Mohamed, 2000. Susceptibility of Biomphalaria alexandrina to infection with Schistosoma mansoni: Correlation with the activity of certain glycolytic enzymes. J. Egypt. Soc. Parasitol., 30: 547-560.
Direct Link - El-Ansary, A., S. El-Bardicy, M.S. Soliman and N. Zayed, 2000. Sublethal concentration of Ambrosia maritime (Damsissa) affecting compatibility of Biomphalaria alexandrina snails to infection with Schistosoma mansoni through disturbing the glycolytic pathway. J. Egypt Soc. Parasitol., 30: 809-819.
Direct Link - El-Ansary, A., E.M. Sammour, M.S. Soliman and F.A. Gawish, 2001. In vivo, attenuation of schistosome cercarial development and disturbance of egg laying capacity in Biomphalaria alexandrina using sublethal concentrations of plant molluscicides. J. Egypt. Soc. Parasitol., 31: 657-669.
Direct Link - El-Ansary, A., A.M. Mohamed, S.S. Mahmoud and S. El-Bardicy, 2003. On the pathogenicity of attenuated Schistosoma mansoni cercariae released from metabolically disturbed Biomphalaria alexandrina. J. Egypt. Soc. Parasitol., 33: 777-794.
Direct Link - Haberl, B., M. Korner, Y. Spengler, J. Hertel, M. Kalbe and W. Hass, 2000. Host finding in Echinostoma caproni: Miracidia and cercariae use different signals to identify the same snail species. Parasitology, 120: 479-486.
Direct Link - Hass, W., K. Grabe, T. Pash, K. Stoll, M. Fuchs, B. Haberl and C. Loy, 2002. Recognition and invasion of human skin by Schistosoma mansoni cercariae: The key role of L-arginine. Parasitology, 124: 153-167.
Direct Link - Hata, H., 1994. Essential amino acids and other essential components of Angiostronglus costaricensis from third-stage larvae of young adults. J. Parasitol., 80: 518-520.
PubMed - Ingram, R.J., A. Bartlett, M.B. Brown, C. Marriott and P.J. Whitfield, 2003. Penetration of human skin by the cercariae of Schistosoma mansoni: An investigation of the effect of multiple cercarial applications. J. Helminthol., 77: 27-31.
Direct Link - Manager, P., L. Jianyong, B.M. Christensen and T.P. Yoshino, 1996. Biogenic monoamines in the fresh water snail, Biomphalaria glabrata: Influence of infection by the human blood fluke, Schistosoma mansoni. Comp. Biochem. Physiol., 114: 227-234.
Direct Link - Nabih, I., Z. El-Dardiri, A. El-Ansary and M. Rizk, 1990. Measurement of some selected enzymatic activities in infected Biomphalaria alexandrina snails. Cell Mol. Biol., 36: 637-642.
Direct Link - Ponder, E.L., B. Fried and J. Shermal, 2004. Free-pool amino acids in Biomphalaria glabrata infected with Echinostoma caproni as determined by thin-layer chromatography. J. Parasitol., 90: 665-666.
Direct Link - Xiao, S., M. Tanner, E.K. N'Goran, J. Utzinger and J. Chollet et al., 2002. Recent investigation of artmether, a novel agent for the prevention of Schistosoma japonicum, S. mansoni and S. haematobium. Acta Trop., 82: 175-181.
Direct Link - Engels, D., L. Chitsulo, A. Montresor and L. Savioli, 2002. The global epidemiological situation of schistosomiasis and new approaches to control and research. Acta Tropica, 82: 139-146.
CrossRef