
Md. Sagir Ahmed
Laboratory of Aquatic Resource Management, Department of Zoology, University of Dhaka, Dhaka 1000, Bangladesh
Md. Raknuzzaman
Department of Fisheries, University of Dhaka, Dhaka 1000, Bangladesh
Hafeza Akther
Faculty of Agriculture, Kagoshima University, Kagoshima 890-0065, Japan
Sumaiya Ahmed
Laboratory of Aquatic Resource Management, Department of Zoology, University of Dhaka, Dhaka 1000, Bangladesh
Journal of Applied Sciences
Year: 2007 | Volume: 7 | Issue: 13 | Page No.: 1785-1789







ABSTRACT
A study was conducted on association of Vibrio cholerae with plankton specially emphasis on cyanobacteria in relation to some physico-chemical parameters in the River Buriganga, Dhaka, from January to December 2002. Monthly abundance of phytoplankton and zooplankton varied from 457 to 14166 and from 169 to 1055 individual L-1, respectively. Monthly average of faecal coliform in water, zooplankton and phytoplankton samples were 3.99x109, 4.54x103 and 4.28x102 (CFU L-1), respectively. During epidemics, toxigenic V. cholerae 01 and 0139 were isolated from the patients as well as from the surface water. V. cholerae 01 and 0139 were also isolated from plankton samples. More over, it was observed that ctx (cholera toxic) positive in water and phytoplankton samples of the river. A bloom of Oscillatoria sp. (1.6x104 individual L-1) occurred in the upper reaches of the River Buriganga in May 2002. Methanol-water extract of bloom sample was analyzed by high performance liquid chromatography with UV detection and Mass Spectrum (MS) detected microcystin-RR. Cyanobacteria are abundant in the aquatic environment of Bangladesh and it was established that V. cholerae maintain a symbiotic relationship with these algae particularly mucilaginous cyanobacteria. During epidemics, patients symptoms included diarrhea, vomiting and hemorrhagic enteritis and in severe cases hemorrhagic diarrhea. So, question has arisen that which is responsible, microcystins or cholera for death of cholera/diarrhea patients in Bangladesh. Future research should be directed to isolate microcystins and cholera toxins from the epidemic areas to clarify the fact.
PDF Abstract XML References Citation
How to cite this article
Md. Sagir Ahmed, Md. Raknuzzaman, Hafeza Akther and Sumaiya Ahmed, 2007. The Role of Cyanobacteria Blooms in Cholera Epidemic in Bangladesh. Journal of Applied Sciences, 7: 1785-1789.
DOI: 10.3923/jas.2007.1785.1789
URL: https://scialert.net/abstract/?doi=jas.2007.1785.1789
DOI: 10.3923/jas.2007.1785.1789
URL: https://scialert.net/abstract/?doi=jas.2007.1785.1789
INTRODUCTION
Cholera is a dreadful diarrhoeal disease from the time immemorial and endemic in many parts of the world (Pollitzer, 1959). It is caused by ingestion of food or water containing Vibrio cholerae, serogroups 01 or 0139. Symptoms include acute watery diarrhoea and vomiting which can result in severe dehydration or water loss. When left untreated, death can occur rapidly-sometimes within hours (WHO, 2000). Bangladesh has been considered as the homeland of cholera and every year about 4.5 million people suffered from watery diarrhea, among them a considerable number affected with cholera. In 1991, an epidemic cholera was responsible for and estimated 8,000 deaths in 12 weeks periods (Siddique et al., 1992). Cholera epidemics occur twice a year and have regular seasonal patterns in the epidemic areas of Bangladesh (Khan et al., 1984). During epidemics, Vibrio cholerae 01 and 0139 were isolated from patients as well as from the surface water of pond, lake and river (Islam et al., 1994, 1992; Islam, 1990; Huq et al., 1990) but apparently disappeared from the environment during the inter-epidemic seasons (Khan et al., 1984). V. cholerae can survive under unfavorable environmental conditions in a dormant state, that is, it is viable but nonculturable (Roszak and Colwell, 1987). Viable but nonculturable V. cholerae contribute to the occurrence of seasonal epidemics because V. cholerae can persist for a long time in the aquatic environment, reintroduction of the organism by infected humans is not necessary. Furthermore, V. cholerae is a microbial inhabitant of brackish water and estuarine ecosystems, that is, it is autochthonous (Xu et al., 1982). Evidence collected over the last 20 years indicates a close association of V. cholerae with copepods and algae (Lipp et al., 2002). The association of V. cholerae with zooplankton has proven to be a key factor in deciphering the global nature of cholera epidemics (Colwell, 1996).
About 307 different species of cyanobacteria has been reported from all kinds of water sources (river, canal, ponds, ditches, lakes etc.) in Bangladesh. Among them 13 species frequently form blooms (Islam, 1991). Islam et al. (1992) detected V. cholerae 01 and 0139 in the cyanobacteria from the aquatic environment of Bangladesh. Islam et al. (1994) demonstrated that cyanobacteria can provide micro environment for the survival of V. cholerae in the inter-epidemic period and can act as a reservoir. It was assumed that seasonal multiplication of V. cholerae following algal bloom formation may plays an important role in maintaining the seasonality of cholera in endemic areas of Bangladesh.
The supply of clean and safe drinking water is one of the main challenges of public health care in Bangladesh. Traditionally, surface water is the main source of drinking water and consumed without any treatment or after boiling when fuel is available. Welker et al. (2004) in a study at three different regions in Bangladesh detected microcystins in 39 ponds, mostly together with varying abundance of potentially microcystin-production genera such as Microcystis, Planktothrix and Anabaena. Total microcystin concentrations in their study ranged between <0.1 and up to >1000 μg L–1 and more than half of the positive samples contained high concentrations of more than 10 μg L–1. The results clearly showed that the concentration of microcystins well above the WHO provisional guideline value of 1 μg L–1 MC-LR.
Recently, there has been an increased use of surface water for human consumption due to arsenic contamination in ground waters. In Bangladesh such a practice could amount to replacing one health hazard with another.
The seasonal pattern of cholera epidemics in Bangladesh and the recent occurrence of microcystions from cyanobacteria led us to investigate the role of cyanobacteria blooms in cholera epidemics.
Present study deals with the seasonal abundance of cyanobacteria in relation to some physico-chemical parameters from the river Buriganga and isolation of faecal colifrom and V. cholerae and their relationship with cyanobacteria and other plankton.
MATERIALS AND METHODS
Present investigation was conducted in the river Buriganga that encompasses the southwestern periphery of the Dhaka, the capital city of Bangladesh. It is considered as one of the most polluted urban rivers in the world.
Collection of samples: Water, phytoplankton and zooplankton samples were collected fortnightly intervals from January to December 2002. Samples of water were collected in pre-sterilized 500 mL narrow-mouth plastic bottles. Phytoplankton and zooplankton were colleted using plankton net of mesh size 20 and 60 μm, respectively and kept in 20 mL glass bottle. All field samples were preserved and transported to the laboratory in an insulated igloo box with cool packs and processed within 6 h of collection.
Physico-chemical parameters were recorded by using an ecological HACH fresh water, aquaculture manual, test kit model FF-2. Qualitative and quantitative analysis of plankton were performed using taxonomic keys following by Wards and Whipple (1959), Needham and Needham (1966), Mellenby (1979) and methods by Welch (1952). Meteorological data were collected from the Department of Meteorology, Dhaka, Bangladesh.
Processing of samples: Phytoplankton samples (10 mL) were homogenized in a Teflon-tipped tissue grinder using a Steed Fast stirrer. Ten milliliter of phytoplankton homogenate and 50 μL of the water sample were enriched separately in bile salts peptone broth and incubated overnight at 37°C. All samples were then plated into Thiosulphated Citrate Bile Salts (TCBS) agar and Taurocholate Tellurite Gelatin Agar (TTGA) and incubated at 37°C for 24 h (Mansur, 1961). Suspected Vibrio colonies were further characterized using standard procedures (West and Colwell, 1984).
Faecal coliform counts: Faecal coliform count was taken from the undiluted water, phytoplankton and zooplankton samples (25 μL each) spreading into MFC agar plate. The plates were then incubated at 44°C for 18-24 h. The characteristic blue colonies were counted as faecal coliforms.
Biochemical and serological tests: For confirmation of bacteria, a series of biochemical reactions were performed by standard methods outlined by WHO Manual (1997) which included Kligler Iron Agar (KIA), Motility Iodole Urea (MIU) and Citrate and Oxidize test. Bacterial cells were confirmed finally by serological reactions. Slide agglutination tests were adopted for this purpose.
Direct Fluorescence Antibody (DFA) test: ICDDR, B, Dhaka provided vibrio cholerae 01 and 0139 monoclonal antibodies and flurescein isothiocyanate (FITC) cojugated antimouse was purchased from the local market. The DFA test was performed using the procedure Huq et al. (1990). The slides were examined under epifluorescence microscope and counted according to Kogure et al. (1979).
Amplification of Cholera Toxin (ctx) a gene for PCR: Primers (ctx 1 and ctx 2) were kindly supplied by Dr. Kaiser Ali Talukder of ICDDR, B, Dhaka. The amplification of ctx A gene in DNA was performed in 50 μL of reaction mixture which contained 10 μL of sample.
Electrophoresis analysis of amplified DNA: The standard method was used to separate, identify and purify DNA through electrophoresis described by Keasler and Hall, (1993).
RESULTS AND DISCUSSION
The seasonal fluctuations of physico-chemical parameters of the River Burigonga during the study period are shown in the Table 1.
A total of 59 species of phytoplankton belonging to 35 genera was identified. The highest number of taxa belonged to Chlorophyta followed by Cyanophyta, Euglenophyta and Chrysophyta (Fig. 1). As a single species Microcystis sp. (1.4x104 cells L–1) dominated through out the year followed by Melosira sp. (2779 individual L–1) and Oscillatoria sp. (2520 individual L–1). Plankton diversity index was moderately rich and evenly distributed. A bloom of Oscilatoria sp. (1.6x104 individual L–1) occurred in the upper reaches of the bloom sample showed Microcystin RR, 235 mg L–1 of original water (details will appear elsewhere). Four types of microcystins have been isolated from cyanobacteria bloom occurred in a pond of cholera epidemic area and the quantity of toxins was much higher than recommended level for safety drinking water (Ahmed et al., 2007).
Faecal coliform in the River water, phytoplankton and zooplankton were counted separately (Fig. 2a and b). The monthly average faecal coliform in the water, zooplankton and phytoplankton samples of Burigonga were 3.99x109, 4.54x103 and 4.28x102 (CFU L–1), respectively. V. cholerae 01 and V. cholerae 0139 were also isolated from water and plankton samples. Agarose gel electrophoresis showed cholera toxin (ctx) positive both in water and phytoplankton samples those confirmed association of V. cholerae with phytoplankton (Fig. 3). This highly polluted water is a threat for aquatic lives and as well as for human being who are using it for domestic and other purposes.
The main cholera epidemics reach its peak during the cooler months of year and a secondary smaller one occurs during the hot season. The reason for maintaining infelicity probably lies in the mechanisms by which V. cholerae survive during inter-endemic periods in these endemic areas. It has been proved that aquatic flora; mainly cyanobacteria act as a reservoir of cholera (Islam et al., 1990).
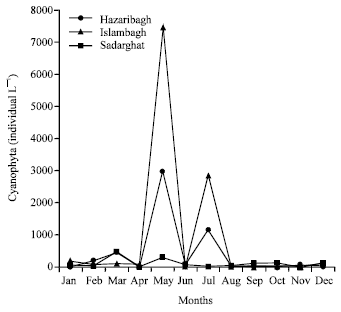 | |
| Fig. 1: | Monthly abundance of cyanophyta (individual L–1) at three different sampling stations in the River Buriganga, from January to December 2002 |
| Table 1: | Monthly variations of physico-chemical parameters in the River Buriganga from January to December 2002 |
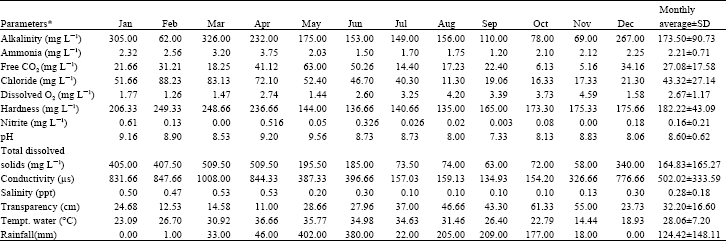 | |
| * These values are from average from three sampling stations | |
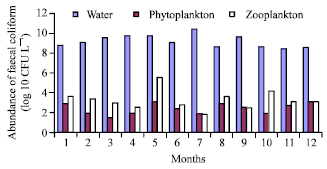 | |
| Fig. 2a: | Monthly abundance of total coliform (Log10 CFU L–1) in water, phytoplankton and zooplankton in Hazaribagh (station A) of the Buriganga (station 3) River from January to December, 2002 |
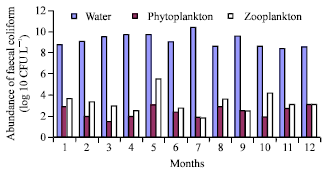 | |
| Fig. 2b: | Monthly abundance of total coliform (Log10 CFU L–1) in water, phytoplankton and zooplankton in Sadarghat (station B) of the Buriganga River from January to December, 2002 |
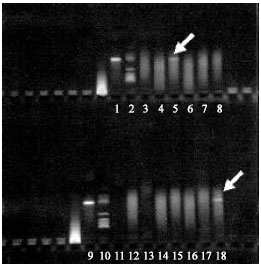 | |
| Fig. 3: | Agarogel electrophoresis showed cholera toxin (ctx) in both water (lane 18) and phytoplankton sample (lane 5) of the river Burigonga. Lane 1 and 9 are positive control |
V. cholerae was detected in the mucilaginous sheath of cyanobacteria, Anabaena variabilis from the eutrophic ponds in Bangladesh (Islam et al., 1990). It was also observed that V. cholerae did not lose their ability to produce cholera toxin during survival in association with cyanobacteria. Large ponds, canals, rivers and estuaries are considered as permanent reservoir of V. cholerae. Cyanobacteria are very common in these water bodies and form blooms. In winter continuous evaporation and human use the water volume is substantially decreased, thereby increasing the concentrations of dissolved organic nutrients. Therefore, increased nutrients, more sunshine and favorable temperatures altogether initiate cyanobacteria bloom formation. After a peak of algal bloom, the vast amount of algae may die and disintegrate due to increased demand of food and the V. cholerae, which had survived inside the mucilaginous sheath, are released into favorable bio-physico-chemical conditions for multiplications. Under all favorable conditions, the rapid multiplication of Vibrios takes place and water becomes heavily contaminated with V. cholerae. Finally, the inhabitants who live around the pond or rivers and use the water for various households and domestic purposes may become infected.
V. cholerae has long been known as a fecal-oral pathogen and indeed, infection rates are significantly greater in areas with poor sanitation; however, the evidence showing that V. cholerae is naturally present in warm, brackish environments is overwhelming. When communities rely on untreated environmental water sources for bathing, cooking and drinking water, the incidental ingestion of copepods, which carry a high dose of V. cholerae, can initiate an infection. Likewise, the likelihood of consuming an infectious dose is higher when a bloom of planktons occurs in the water (Lipp et al., 2002). So, the households which filter their pond water through sari cloth before drinking reduce their risk of cholera infections by approximately 50% (Huq et al., 2001).
In regions of cholera endemicity, the disease does not disappear after an epidemic peak and returns in successive waves (Torres, 2001). Of particular interest and relevance to identifying environmental or climate factors that may promote epidemics is the understanding of dynamics of the disease in areas of endemicity. For example, what causes periodic oscillations in cholera outbreaks and why are some areas more prone to endemism.
CONCLUSIONS
Cyanobacteria are abundant in the aquatic environment of Bangladesh and it was established that V. cholerae maintain a symbiotic relationship with these algae particularly mucilaginous cyanobacteria. Rapid multiplication of V. cholerae takes place following the cyanobacteria bloom formation in ponds, lakes and rivers. Seasonal multiplication of V. cholerae 01 and 0139 following algal bloom formation plays an important role in maintaining the seasonality of cholera in endemic areas of Bangladesh. Four types of microcystins have been isolated from cyanobacteria bloom occurred in ponds and rivers of epidemic area. Although there is no official record of animal or human intoxication induced by cyanobacteria, the role of microcystins in deaths associated with cholera/diarrhea should be clarified.
REFERENCES
- Huq, A., R.R. Colwell, R. Rahaman, A. Ali and M.A.R. Chowdhury et al., 1990. Detection of Vibrio cholerae 01 in the aquatic environment by fluorescent-monoclonal antibody and culture methods. Applied Environ. Microbiol., 56: 2370-2373.
Direct Link - Islam, M.S., M.J. Alam and P.K.B. Neogi, 1992. Seasonality and toxigenicity of Vibrio cholerae non 01 isolated from different components of pond ecosystems of Dhaka city, Bangladesh. World J. Microbiol. Biotechnol., 8: 160-163.
Direct Link - Islam, M.S., B.S. Drasar and R.B. Sack, 1994. The aquatic flora and fauna as reservoirs of Vibrio cholerae. A review. J. Diarrhoea Dis. Res., 12: 87-96.
Direct Link - Lipp, E.K., A. Huq and R.R. Colwell, 2002. Effects of global climate on infectious disease: The cholera model. Clin. Microbiol. Rev., 15: 757-770.
Direct Link - Roszak, D.B. and R.R. Colwell, 1987. Survival strategies of bacteria in the natural environment. Microbiol. Rev., 51: 365-379.
PubMedDirect Link