
Gamal M.A. Lashin
Department of Botany, Faculty of Sciences, Zagazig University, Zagazig, Egypt
Journal of Applied Sciences
Year: 2007 | Volume: 7 | Issue: 9 | Page No.: 1304-1313







ABSTRACT
Palynostratigraphic studies of the Lower Cretaceous sediments, Alamein Formation, Bahrein-1 well in the northern part of the Western Desert, Egypt, have been carried out from an evolutionary and geological viewpoint. The palynoflora principally consists of pteridophytes, gymnosperms and some rare angiosperm pollen. The microflora shows that the palaeoenvironments were favorable for different kinds of plants. The recovered microflora, therefore, indicates an Early Aptian age and can be extended to the Early Albian. The palynological data revealed that the North Western Desert (N.W.D.) of Egypt belongs to the North Gondwana phytogeoprovince in the Early Cretaceous.
PDF Abstract XML References Citation
How to cite this article
Gamal M.A. Lashin, 2007. Palynostratigraphic Studies of the Lower Cretaceous Sediments,Alamein Formation, Bahrein-1Well in the Northern Part of the Western Desert, Egypt. Journal of Applied Sciences, 7: 1304-1313.
DOI: 10.3923/jas.2007.1304.1313
URL: https://scialert.net/abstract/?doi=jas.2007.1304.1313
DOI: 10.3923/jas.2007.1304.1313
URL: https://scialert.net/abstract/?doi=jas.2007.1304.1313
INTRODUCTION
Recently palynology has contributed significantly to our knowledge of the evolution and the origin of angiosperms. Palynological studies started almost hundred years ago in Egypt. The main interest is the Lower-Cretaceous microflora and its correlation with the biostratigraphy of the same time in different localities.
In the past few decades, the area of Egypt in general and the Western Desert in particular have been subject to more palynological studies, which threw the light on the microflora characteristics of the Lower-Cretaceous. The studied section of the Bahrein-1 borehole is generally barren of diagnostic macrofauna and consequently the delineation of the Lower Cretaceous stratigraphic boundaries is rather a questionable problem (Sultan, 1986).
Most of the previous studies of the seventies were concentrated on the subsurface stratigraphy of the Western Desert especially Jurassic and Cretaceous rocks. Noteworthy, the works of Saad (1974 and 1978), Soliman (1976), Sultan (1978), Saad and Ghazaly (1976), Sultan and Soliman (1976) and Aboul Ela (1978 and 1979).
During the eighties and nineties decades the Cretaceous palynological studies were covered most of the area of Egypt. We refer in this context to the works of, Abdel Malik et al. (1981), Schrank (1987), Abdel Mohsen (1987 and 1992), Sultan (1986 and 1987), Sultan and Ali (1986), El-Beialy (1993, 1994 and 1995), El-Beialy et al. (1990), EL-Shamma (1988 and 1991), El Shamma and Arafa (1988), Omran (1989) and Ismail and Soliman (1997). The palynomorphs and palynozonation especially in Western Desert have been discussed by many authors e.g., Aboul Ela et al. (1989), Omran et al. (1990), Abd El-Shafy and Abd El-Moneim (1991), Soliman et al. (1991), Bassiuni et al. (1992), El-Shamma and Baioumi (1992), Ibrahim (1996), Ibrahim et al. (1995), Schrank and Ibraheim (1995), Ibrahim and Schrank (1996), Schrank and Mahmoud (1998), Lashin (1999), Mahmoud and Moawad (2000) and Schrank and Mahmoud (2002). The present research deals with the palynostratigraphy of undifferentiated Lower Cretaceous clastic rock sequence intersected in Bahrein-1 well. The research aims also to study the Aptian palynology from evolutionary viewpoint, evaluate the palaeoenvironments based on the recovered palynomorphs and precise or correct age determination.
MATERIALS AND METHODS
The study based on the investigation of ten rock samples obtained from Bahrein-1 well in the Western Desert (Fig. 1). According to the geological composite well logs of Petroleum companies (Composite well log of the studied well was kindly provided by the Egyptian General Petroleum Authorities), the represented Cretaceous age is Aptian time (Alamein formation). The ages from which we managed to obtain the ten samples are those shown in Fig. 2.
Geological setting:
Bahrein-1 well (B-1): Was drilled by General Petroleum Co. in 1970 and is located at Lat.: 28° 48` North and Long: 26° 33`15`` East (Fig. 1).
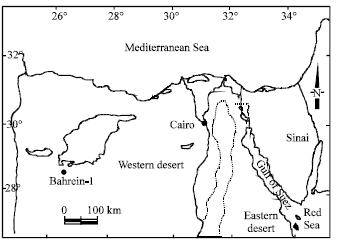 | |
| Fig. 1: | The locality of the studied Bahrein-1 borehole ● (After Ibrahiem, 1996) |
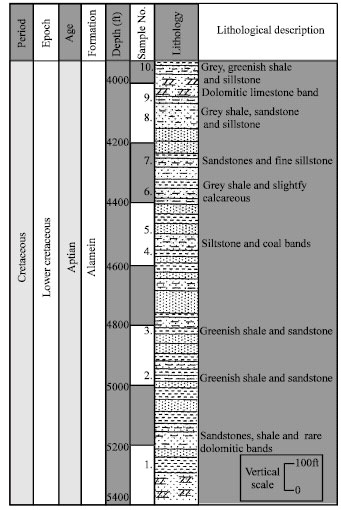 | |
| Fig. 2: | Part of the geological section of Bahrein-1 well, showing depths of studied samples and their lithological description |
The well traversed strata belonging to Eocene, Cretaceous and Jurassic and was bottomed in the Paleozoic sediments with a total depth of 10022 ft. (Norton, 1967). The sample depths and their lithological description are as shown in Fig. 2. Thus according to the composite well logs the studied samples are mainly Cretaceous particularly Lower Cretaceous (Aptian) and belonging to Alamein Formation.
The ten samples investigated palynologically from that well. The samples lithology consists mainly of alternating shale, silt shale and carbonates. The technical procedure for preparation and palynological investigation of the selected samples was that adopted by Herngreen (1983). Permanent slides were prepared (5 slides from every sample) using glycerin jelly as a mounting medium. The slides were investigated for palynology, facies and deposited environments using the light microscope. Some of these samples especially those of the Upper Cretaceous sediments (mainly carbonates) were found to be palynologically barren. The palynomorphs assemblages are examined and identified. The representative forms illustrated in Fig. 3-23 and their palynogram presented in Fig. 24. The materials were deposited in the Department of Botany, Faculty of Sciences, Zagazig University, Egypt.
 | |
| Fig. 3: | Triplanosporites sp. |
 | |
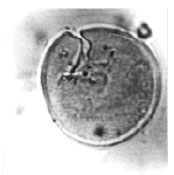
| Fig. 4: | Todisporites minor Couper, 1953 |
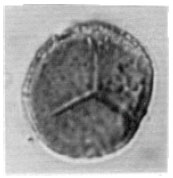 | |
| Fig. 5: | Trilete spore |
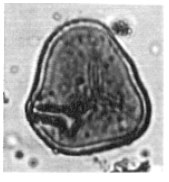 | |
| Fig. 6: | Cyathidites minor Couper, 1953 |
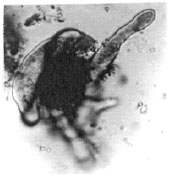 | |
| Fig. 7: | Elaterosporites klaszii (Jardine nd Magloire) Jardine, 1967 |
 | |
| Fig. 8: | Spheripollenites sp. |
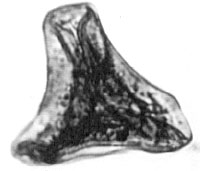 | |
| Fig. 9: | Gleicheniidites cf. senonicus Ross, 1949 |
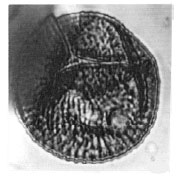 | |
| Fig. 10: | Cicatricosisporites cf. venustus Deak, 1963 |
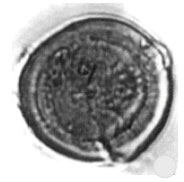 | |
| Fig. 11: | Classopollis sp./cf. C. torosus Reissinger) Couper, 1958 |
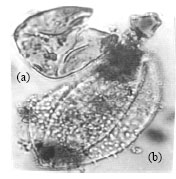 | |
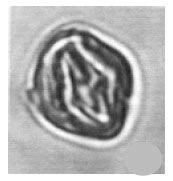
| Fig. 12: | (a) Cyathidites australis Couper, 1953 and (b) Stellatopollis sp. |
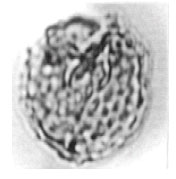 | |
| Fig. 13: | Brenneripollis sp. |
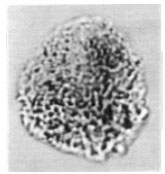 | |
| Fig. 14: | Afropollis operculatus Doyle et al., 1982 |
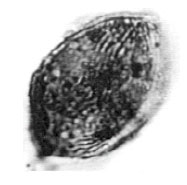 | |
| Fig. 15: | Classopollis sp. |
 | |
| Fig. 16: | Tricolporopollenites sp. x 2000 |
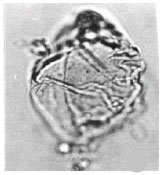 | |
| Fig. 17: | Dinoflagellates |
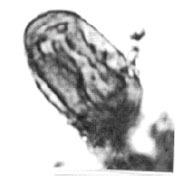 | |
| Fig. 18: | Eucommiidites sp. |
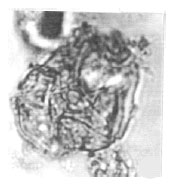 | |
| Fig. 19: | Cretacaeiporites polygonalis (Jardine and Magloire) Herngreen, 1973 |
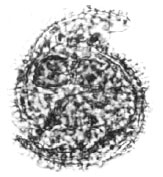 | |
| Fig. 20: | Chorate dinoflagellates |
 | |
| Fig. 21: | Ephedripites sp. |
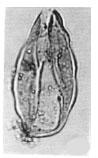 | |
| Fig. 22: | Cycadopites sp. |
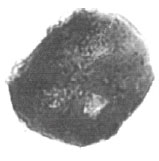 | |
| Fig. 23: | Cretacaeiporites scabratus Herngreen, 1973 |
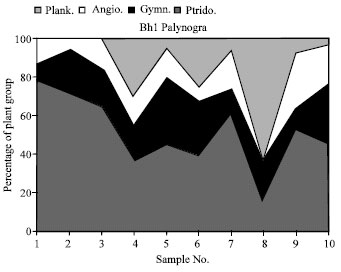 | |
| Fig. 24: | Palynogram of the plant groups distribution throughout the studied section (Ptreid. = Ptreidophytes, Gymn. = Gymnosperms, Angio = Angiosperms, plank. = Phytoplanktons |
RESULTS
The samples from Bahrein-1 well, covering an interval (interval from 5296 to 3969 ft.) of Lower Cretaceous sediments (Alamein Fm.) are palynologically studied. The important index sporomorphs, which recovered from the studied well illustrated in Fig. 25, 3-23 and represented by palynogram (Fig. 24).
The palynomorphs taxa comprise more than of 28 genera and 29 species (Fig. 24, 25 and 3-23). The taxa characterized by the dominance of pteridophyte spores with the percentages of 60% contribute 13 genera. The Gymnosperms about 20% and contribute 6 genera and 6 species. The angiosperms about 10% contribute 7 genera and 8 species and microphytoplanktons about 10%, contribute 2 assemblages.
The lower part (sample No. 1-4) reveals a less diversity. The pteridophyte sporomorphs characterizing the Early Cretaceous time are recorded and represented mainly by Cyathidites, Todisporites, Concavissimisporites variverrucosus, C. punctatus, Perotrilites sp., Cicatricosisporites sp., Matonisporites sp., Gleicheniidites, Triplanosporites sp. and Dictophyllidites sp.
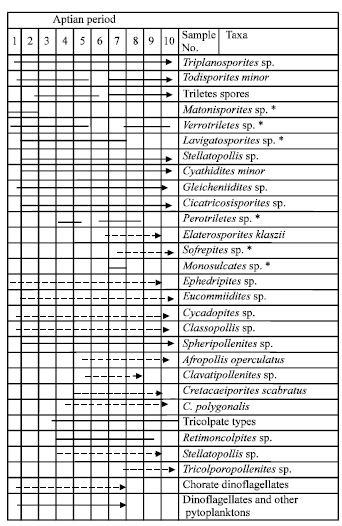 | |
| Fig. 25: | The representative palynomorphs of the studied sample (*) Not illustrated in the plate (⟶) Common (- - -), rare (⤑) very rare |
The gymnosperm pollen as Ephedripites sp., Monosulcates, Eucommiidites sp., Cycadopites and Classopollis sp., were recognized. The angiosperms were rare and represented only by some tricolpate, retimonocolpate types and some ill preserved Afropollis pollen. A few scattered occurrences of some chorate dinoflagellates. Other phytomicroplankton as Chomotriletes sp. also represented.
The uppermost part of the studied interval (sample No. 5-10) was characterized by different varieties of sporomorphs. The pteridophyte sporomorphs characterizing the Early Cretaceous were recorded and represented mainly by Cyathidites, Cicatricosisporites sp., Todisporites, Lycopodiumsporites, Gleicheniidites Concavissimisporites variverrucosus, C. punctatus and Perotrilites sp. The gymnosperms pollen of the genera Classopollis, Araucariacites, Spheripollenites, Ephedripites, Eucommiidites, Monosulcites and Callialasporites are recorded. Angiosperms are common in this interval and represented by Tricolpites, Clavatipollenites, Liliacidites, Cretacaeiporites, Afropollis, Brenneripollis, Stellatopollis sp. and Tricolporopollenites sp. Elaterates as Elaterosporites klaszii are present. Dinoflagellates are few and scattered occurrences.
DISCUSSION
The palynomorphs assemblages of Alamein formation are diverse and possess numerous elements which have been described from Lower Cretaceous and could also be continued upwards from Aptian to Albian. The lack of diversification in the pollen and spores in the studied samples may be related to poor preservation under increasingly sub aerial continental conditions.
At the depth of 5399 ft., the occurrence of Cicatricosisporites sp., Matonisporites equiexinus, Todisporites minor, Gleicheniidites cf. senonicus and Chomotriletes sp. (phytoplankton) may attribute to the Lower Cretaceous flora (Bassiouni et al., 1992). Aboul Ela et al. (1989) reported that Cicatricosisporites sp. was common and important during Barrisian-Hautervian rocks of N.W.D. of Egypt. These taxa are closest to palynofloras previously described from the late Barremian of North and Western Africa (Schrank, 1992; Herngreen et al., 1996, Jardene, 1967).
At the depth 4999-4840 ft., the main of pteridophyte spores are Triplanosporites sp., Lavigatosporites, Cyathidites and Lycopodiumsporites. Cyathidites were recorded in Lower Jurassic-Early Cretaceous by Saad and Ghazaly (1976). The gymnosperm pollen as Ephedripites sp., Monosulcates, Eucommiidites sp., Cycadopites and Classopollis sp., were also recognized. Ibraheim et al. (1995) stated that the Barremian in NWD is characterized by Ephedripites sp. and primitive angiosperms Liliacidites and fixed that Ephedripites are dominant in Albian of Western Desert microflora. Herngreen and Chlonova (1981) reported the Classopollis sp., in Neocomian time. The aquatic fern spores appear at first in the Neocomian-Barremian (Abdel Malike et al., 1981) and were know to appear at the first in Lower Cenomanian and continued downward into Albian where they disappeared (Herngreen, 1975; Batten and Uwins, 1985; Thusu and Der Eem, 1985). Angiosperms were very rare. The oldest angiosperm pollen so far was known from Egypt during the Late Barremian (Penny, 1991; Schrank, 1992). Sporadic marine influence in this interval represents by a few of chorate dinoflagellates. These may indicate the regression and transgression of marine shore line or the impact of the terrestrial elements by the river into the sea. Related assemblages have been recorded from different wells in the Western desert by Saad and Ghazaly (1976), Abdel-Malik et al. (1981), Schrank (1987, 1992), Sultan (1986) and El-Shamma and Baioumi (1992) and attributed to the Lower Cretaceous (Neocomian-Barremian age).
At the depth 4500-4460 ft., the main of pteridophyte spores were Lavigatosporites, Cyathidites, Lycopodiumsporites, Gleicheniidites and some Perotrilites. The gymnosperm pollen as Ephedripites sp., Cycadopites, Spheripollenites sp. and Classopollis sp., were recognized. Angiosperms were rare and represented only by some tricolpate, retimonocolpate and some ill preserved Afropollis sp., Pollen. Few of chorate dinoflagellates and some aquatic fern spores of perotriletes were recorded. These may indicate the regression and transgression of marine shore line or the in put of the terrestrial elements by the river into the sea. Similar assemblages have been recorded from different wells in the Western Desert by Saad and Ghazaly (1976), Sultan (1986), Penny (1991), Schrank (1992) and El-Shamma and Baioumi (1992) and attributed to the Lower Cretaceous (Barremian age).
At the depth 4270-4370 ft., the pteridophyte spores as Perotrilites were common. The gymnosperm pollen as Monosulcates, were recognized. The angiosperms were rare and represented by tricolpate, retimonocolpate, Stellatopollis sp. and Elaterosporites klaszii pollen. The marine influence in this interval represented by a few scattered occurrences of dinoflagellates and some aquatic fern spores. Related assemblages have been recorded from different wells in the Western Desert by Saad and Ghazaly (1976), Schrank (1992), Sultan (1986) and El-Shamma and Baioumi (1992) and are attributed to the Lower Cretaceous (Barremian-Aptian).
At the depth of 4040-4098 ft., the assemblages are Murospora sp. (pad preserved), Afropollis operculatus and Clavatipollenites, Cretacaeiporites, Tricolpites, Stellatopollis and Elaterosporites klaszii were reported and indicate age not older than Aptian. In a similar stratigraphic interval a number of angiosperm pollen, has been recorded by Saad (1978), El-Shamma and Baioumi (1992) and Lashin (1999) (from the Salam oil field-Bahariya formation) in the North Western Desert. Moreover, these assemblages could be due to the transition between Aptian and Albian age.
At the depth 3960 ft., the miospores spectra of this interval are characterized by the presence of pteridophyte spores as Lavigatosporites, Cyathidites and Gleicheniidites. The gymnosperm pollen as Ephedripites sp., Cycadopites, Eucommiidites sp. and Classopollis sp., are recognized. But angiosperms are represented by tricolpates, retimonocolpates, Elaterosporites, Tricolporopollenites sp. and Cretacaeiporites polygonalis pollen. The oldest angiosperm Tricolporopollenites sp. pollen so far known from Egypt and north Africa during Late Aptian or Albian (Penny, 1991; Schrank, 1992; Uwins and Batten, 1985; Warr and Doyle, 1990; Lashin, 1999). The influence of marine is absent in this interval. These may indicate that the terrestrial elements carried by the river fare from the sea. The related assemblages had been recorded from different wells in the Western Desert by Saad and Ghazaly (1976), Abdel-Malik et al. (1981), Schrank (1992), Sultan (1986) and El-Shamma and Baioumi (1992) and are attributed to the Lower Cretaceous (Barremian-Albian age).
Age correlation: It is well known that, the definition of the Lowe Cretaceous according to the microfloras is controversial especially in sections of non-diagnostic macrofauna or in terrestrial facies. Also the biostratigraphic value of many taxa is still uncertain. However, the age assignment of the described assemblage could be achieved by comparison with stratigraphically well defined assemblages in other regions.
Huges and Mc-dougall (1987) mentioned that the first appearance of Ephedripites is in Barremian-Albian age. El-Saadawi and Kedves (1991) recovered the Ephedripites in Early Cretaceous rocks. Cycadopites and Eucommiidites described by Schrank (1987) during the Lower Cretaceous from the Sudan and NWD of Egypt. Saad (1978) also described Cycadopites and Eucommiidites from UmBarka well during Barremian time. Araucariacites, Eucommiidites and Ephedripites characterize the Pre-Albian of West Africa South America Province (WASA) of Herngreen and Chlonova (1981), Uwins and Batten (1985) and Ward and Doyle (1990).
However, the Lower Cretaceous is widely viewed as the starting point of angiosperms evolution (Cronquist, 1988). The most important record is the presence of some Elaterates and ill preserved angiosperms especially Afropollis sp., tricolpate and Tricolporopollenites sp. These palynomorphs are important in palynostratigraphy for Upper Aptian and Lower Albian ages in the Northern Egypt and Africa (Jardine, 1967; Sultan, 1978; Saad, 1978; Penny, 1986; El-Shamma, 1991; El-Beialy, 1994). The first appearance of tricolpate pollen indicate strongly the beginning of the Albian as argued by Couper and Huges (1963). They described the first tricolpate type in Lower Aptian-Early Albian. Saad (1978) and Doyle et al. (1982) considered that the first appearance of Afropollis jardinus at higher stratigraphic horizon is a reliable indicator for Aptian age. These are also well known from Aptian to Cenomanian strata of Brazil (Herngreen, 1973, 1974; Doyle et al., 1977). The later recognized the A. jardinus and considered the first appearance of this angiosperm type in equatorial Africa as a stratigraphic marker for Early Aptian age. The tricolporate angiosperms and elater bearing elements are present in the Early Albian-Early Cenomanian (Herngreen, 1975; Hochuli, 1981; Srivastava, 1984; Herngreen and Jimenez, 1990; Méon et al., 1996).
Palaeoenvironments: The microphytoplanktons are present in some samples in the middle of this interval. These could indicate the mixed freshwater elements with the marine shoreline or indication of near facies during the deposition or sedimentation time (Schrank and Mahmoud, 1998). The microfloras show that the palaeoenvironments was favorable for different kinds of plants (Fig. 24). From the Bh-1 palynogram, the high percentage were the pteridophytes sporomorphs (80%) in the lower part of the profile (sample No. 1-3), also the angiosperms and gymnosperms about 20%. These indicate that the terrestrial wet land and regression of the marine more common in that time. In the middle of the interval (sample No. 4-7), the pollen flora are common and we suggest that the shore line of lagoon or brackish water are dominant and dry period were long (Schrank and Mahmoud, 1998). In the upper part of the interval (sample No. 8), the transgression is more effective and the phytoplankton facies are common (El-Shamma and Arafa, 1988; Schrank, 1987). The lack of the pteridophytes was influence by the abundance of terrestrial pollen bearing plants (Sultan, 1986).
CONCLUSIONS
The recovered microflora, therefore, indicates an Early Aptian age and can be extended to Early Albian. The palynological results could reveal also that the North Western Desert (NWD) of Egypt belongs to the North Gondwana phytogeoprovince in the Early Cretaceous time. Results recovered here show that the age may not be Aptian. This is different from that given by composite well logs. Therefore, we argue that the correct age is Late Barremian-Aptian-Early Albian.
Finally, the author suggests that more studies should be carried out in the future for these stratigraphic sediments from different views of biostratigraphy to precisely determine the age of the Cretaceous time in the Western Desert of Egypt.
LIST OF MIOSPORES
All specimens are illustrated in the Fig. (3-23) at the magnification x 800 unless otherwise stated.
Ptreidophyte spores:
| • | Triplanosporites sp. (Fig. 3) |
| • | Todisporites minor Couper, 1953 (Fig. 4) |
| • | Trilete spore (Fig. 5) |
| • | Cyathidites australis Couper 1953 (Fig. 12) |
| • | Cyathidites minor Couper 1953 (Fig. 6) |
| • | Elaterosporites klaszii (Jardine and Magloire) Jardine, 1967 (Fig. 7) |
| • | Gleicheniidites cf. senonicus Ross, 1949 (Fig. 9) |
| • | Cicatricosisporites cf. venustus Deak, 1963 (Fig. 10) |
Gymnosperm pollen:
| • | Spheripollenites sp. (Fig. 8) |
| • | Classopollis sp./cf. C. torosus (Reissinger) Couper, 1958 (Fig. 11) |
| • | Classopollis sp. (Fig. 15) |
| • | Ephedripites sp., 32 μm (Fig. 21) |
| • | Eucommiidites sp. (Fig. 18) |
| • | Cycadopites sp. (Fig. 22) |
Angiosperms like-pollen:
| • | Stellatopollis sp. (Fig. 12) |
| • | Brenneripollis sp. (Fig. 13) |
| • | Afropollis operculatus Doyle et al., 1982 (Fig. 14) |
| • | Tricolporopollenites sp. x2000 (Fig. 16) |
| • | Cretacaeiporites polygonalis (Jardine and Magloire) Herngreen, 1973 (Fig. 19) |
| • | C. scabratus Herngreen, 1973 (Fig. 23) |
Phytoplanktons:
| • | Dinoflagellates (Fig. 17) |
| • | Chorate dinoflagellates (Fig. 19) |
REFERENCES
- Ibraheim, M.I.A., 1996. Aptian-Turonian palynology of the Ghazalat-1 Well (GTX-1) Qattara Depression, Egypt. Rev. Palaeobot. Palynol., 94: 137-168.
Direct Link - Ismail, A.A. and S.L. Soliman, 1997. Cenomanian-Santonian foraminifera and ostracodes from Horus Well-1, North Western Desert, Egypt. Micropleontology, 43: 165-183.
Direct Link - Mahmoud, M.S. and A.R. Moawad, 2000. Jurassic-Cretaceous (Bathonian to Cenomanian) palynology and stratigraphy of the West Tiba-1 borehole, northern Western Desert, Egypt. J. Afr. Earth Sci., 30: 401-416.
Direct Link - Schrank, E. and M.S. Mahmoud, 2002. Barremian angiosperm pollen and association palynomorphs from the Dakhla Oasis area, Egypt. Paleontology, 45: 33-56.
Direct Link