
Yesim Buyukates
Department of Fisheries, SUF, Canakkale OnsekizMart University, Canakkale, 1 71 DC), Turkey
Daniel L. Roelke
Texas A&M University, Wildlife and Fisheries Sciences, 2258 TAMUS, College Station, TX 77843-2258, USA
Journal of Applied Sciences
Year: 2005 | Volume: 5 | Issue: 5 | Page No.: 820-828







ABSTRACT
Aquatic systems are dynamic in nature and can be affected diversely by various fluctuations. Additionally, survival and development of organisms living in require that the system in question be of sufficient quality to sustain the different components. Therefore, a thorough knowledge of the system and the processes affecting the system are required. Field samplings were performed to gather information on the plankton community and nutrient dynamics of a tidal creek of lower Nueces Delta, southeast Texas, USA on 15 March, 7 June and 8 September 2001. On site measurements and microscopic analyses showed that environmental parameters and plankton community composition changed considerably among sampling stations and sampling dates. The system was found to be nitrogen limited confirming previous studies. Naturally, silica concentrations were high in the system, however flooding events caused an additional increase in September. The natural plankton community structure was diverse in the system. Both zooplankton and phytoplankton community composition changed due to environmental conditions, especially rain events and decrease in salinity.
PDF Abstract XML References Citation
How to cite this article
Yesim Buyukates and Daniel L. Roelke, 2005. Investigating System Characteristics of a Southeast Texas Wetland: Nutrient and Plankton Dynamics of a Tidal Creek in Lower Nueces Delta. Journal of Applied Sciences, 5: 820-828.
DOI: 10.3923/jas.2005.820.828
URL: https://scialert.net/abstract/?doi=jas.2005.820.828
DOI: 10.3923/jas.2005.820.828
URL: https://scialert.net/abstract/?doi=jas.2005.820.828
INTRODUCTION
A thorough knowledge of an aquatic system is paramount to understand the individual and/or mutual effects of physical and biological environments on the plankton community composition.
Nueces Estuary system includes the upper Nueces Delta salt marsh and lower Nueces Delta (southeast Texas, USA). Lower Nueces Delta consists of brackish and saltwater marshes, mudflats and open water and covers approximately 75 km2 (Fig. 1)[1]. Other dominant features of the study site include the lower Nueces River, which encompasses a discharge site for the Corpus Christi Allison Sewage Treatment Plant (STP) and an area of heavy industry dominated by oil refineries[2]. The system is shallow with a maximum depth of 1.2 m and it is completely mixed through the water column. Salinity varies with river inflow from Calallen Gauge, weak tidal exchange, precipitation and strong wind-driven water exchange with the bay. Regarding productivity, the system supports many estuarine organisms that are commercially important. These include the brown shrimp Penaeus aztecus, many species of fin fish, crabs and oyster reefs[1,3]. Additionally, benthic diatoms, cyanobacteria, Spartina spartinea provide the main food sources for these organisms[1,3]. Furthermore, phytoplankton primary production is controlled by nitrogen (N) availability throughout the year[1,4].
Fundamental understanding of the system depends on the freshwater requirements of the region, which consists of many elements: the environmental health of the wetland, the region’s industry, damming and channelization within the Nueces watershed[1,2]. Freshwater input to the wetland is important to supply the nutrients that are necessary for the growth and development of phytoplankton that is the main food source of zooplankton, mollusks and fishes[1,2]. In recent years, it is shown that extended drought conditions, altered hydrology of the delta due to dam construction and channelization and increased water demand by the petroleum industry has caused lack of freshwater input and hypersalinity in the system[1,5]. Hypersaline conditions are known to alter the osmotic concentration of seawater, relative solutes within the estuarine water, concentration of dissolved gasses and density and viscosity of water[6].
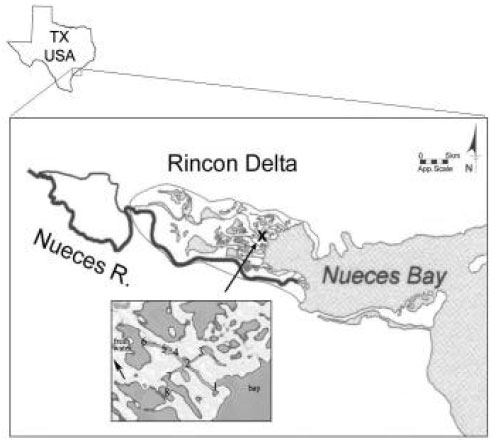 | |
| Fig. 1: | The Nueces River and Bay and the Rincon Delta. The delta is covered with vegetated marshes, mudflats and open water. The system is shallow with a maximum depth of 1.2 m (from Bureau of Reclamation 2000) |
Consequently, the growth and survival of phytoplankton and the associated higher trophic levels are affected. As demand for freshwater from other elements increases, the amount of water available to meet the needs of the marsh ecosystem decreases. Therefore, it becomes problematic to maintain the environmental health of the marsh ecosystem.
We conducted this study to show that the natural plankton community structure in a tidal creek of the Nueces Delta is diverse and the community structure will change due to physical processes especially rain events and/or extended drought conditions. Additionally, the overarching objective of this study was to understand the relations between phytoplankton growth and their physical, chemical and biological environments in the system.
MATERIALS AND METHODS
Field samplings were performed to gather information on the plankton community of a tidal creek of Nueces Delta. Two end members (freshwater and saltwater) and 8 stations are assigned for sampling through one of the creeks of Nueces Delta (Fig. 1). Water samples were collected from each station for nutrient, chlorophyll a (chl a) analyses, as well as phytoplankton and zooplankton enumeration on 15 March, 7 June and 8 September 2001. Environmental parameters such as pH, temperature, dissolved oxygen, redox potential and salinity were measured on site at the time of sampling from surface water using a Hydrolab H20 Water Quality Multiprobe.
Water samples (100 mL) for nutrient analysis were collected from the surface and filtered through 47 mm Whatman GF/F glass fiber filters at the site. Samples were kept frozen until analyses. Analyses of inorganic nutrients such as nitrate (NO3), ammonium (NH4) and nitrite (NO2) which form the dissolved inorganic nitrogen (DIN) pools and orthophosphate (PO4) and silicate (SiO3) were conducted using an automated continuous flow analyzer[7].
Chl a was analyzed with a model 10-AU Turner Designs fluorometer after extraction by 90% acetone[8]. Filters that were used for filtration of surface water for the nutrient analysis were wrapped in aluminum foil and kept frozen until analysis.
Whole water phytoplankton samples were collected to use in the enumeration of phytoplankton groups in the system. This was done to the genus level. Zooplankton samples were collected using a Schindler trap having a 63 μm mesh size cod end and enumerated to major taxonomic groups, i.e., adult copepods, nauplii, rotifers and protozoan. Phytoplankton samples were preserved with buffered gluteraldehyde (5% v/v) and zooplankton samples were preserved with buffered formaldehyde (5% v/v). Plankton identification and enumeration were conducted using settling chambers and inverted microscopy[9]. Sample volumes of 3 mL for phytoplankton and 5 mL for zooplankton were used. At least 20 random fields of view were counted at 1000X, 400X and 200X magnifications for different cell-size classes of phytoplankton. This resulted in at least 400 individuals counted of each dominant phytoplankton species and a ±10% counting precision within 95% confidence limit [10]. The entire settled area was counted for zooplankton. Phytoplankton cell volumes were estimated by measuring cell dimensions and using common geometric shapes[11] and zooplankton sizes were estimated by measuring the length of each individual. Phytoplankton species diversity was determined using the Shannon-Weaver index[12]:
| (1) |
where, pi was the proportion of biomass for species i relative to the total biomass at a specific time.
Finally, the relation among phytoplankton and zooplankton groups as well as N:P (dissolved inorganic nitrogen to phosphate ratio) and salinity were analyzed with simple regression analysis using Statview 5[13]. Although some of the parameters were used in the analyses, all data have been included in the study.
RESULTS
Analyses showed that Si concentrations were always high in the system, NO3-nitrogen was the main source of the DIN and N was the limiting nutrient for the primary production (Fig. 2). Depending on the weather conditions, tidal action and river flow, salinity and water temperature changed considerably among sampling dates (Table 1). Especially during September sampling, the salinitydecreased up to 5 psu. due to recent rain and flooding and the temperature was 27°C (Table 1).
| Table 1: | Environmental parameters from each station on different dates |
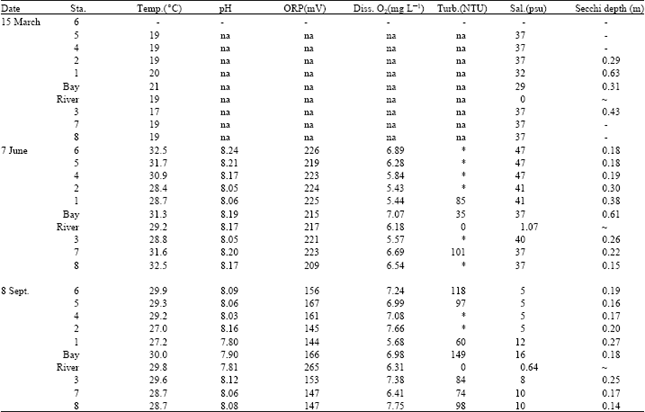 | |
na : not able to sample due to equipment problems; * : too turbid to sample; - : too shallow to sample; ~ : not measured in that particular station | |
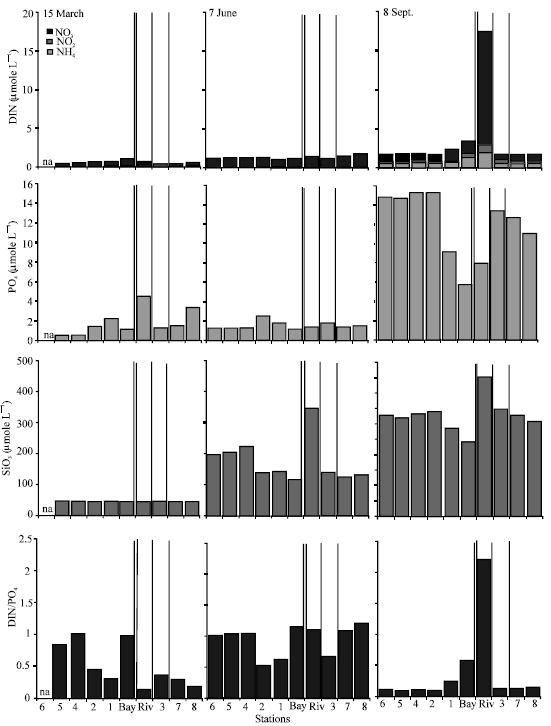 | |
| Fig. 2: | Concentration of dissolved inorganic nitrogen, phosphate, silicate and N:P distribution for March, June and September sampling. The thin, thick and double vertical lines separate the stations that were in the pool areas and river (Fig. 1. St. 3, 7, 8) from the other areas |
In all samplings chl a maximum coincided with the phytoplankton concentration maximum (Fig. 3). Analyses showed moderately strong correlation and statisticallysignificant relationship at 95% confidence level between total phytoplankton biovolume and chl a in all samplings(Table 2). In March and September, diatoms and green algae dominated the phytoplankton while in June diatoms and dinoflagellates were the dominant groups (Fig. 3). Diatoms were mostly dominated by pennates, specifically Nitzschia and Navicula species at all times (Fig. 4). Additionally, small centric diatoms were abundant in September.
Composition of green algae changed considerably depending on the season (Fig. 4).
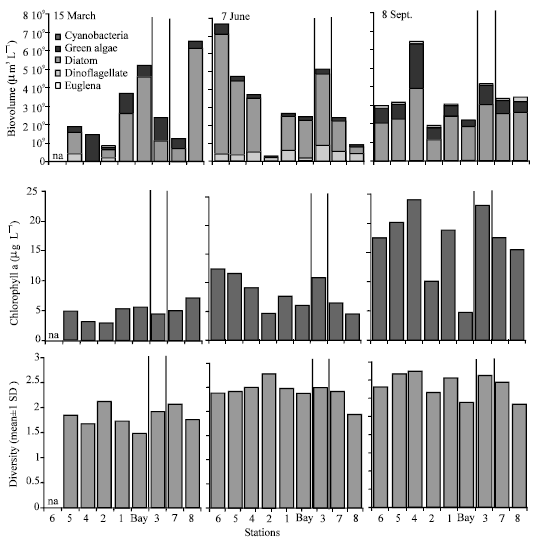 | |
| Fig. 3: | Phytoplankton biovolume, chl a concentration and phytoplankton species diversity for March, June and September sampling. Data were not available for station 6 in March. The thin and thick vertical lines separate the stations that were in the pool areas (Fig. 1. St. 3, 7, 8) from the other areas |
| Table 2: | Dependence of % phytoplankton groups on dissolved inorganic nitrogen to phosphate ratio and salinity and total phytoplankton biovolume to chla |
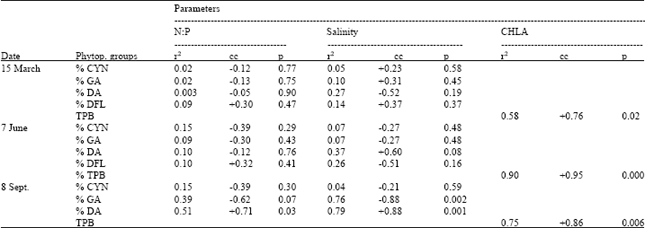 | |
CYN = Cyanobacteria, GA = Green algae, DA = Diatoms, DFL = dinoflagellates, TPB = Total Phytoplankton Biovolume, N:P = dissolved inorganic nitrogen to phosphate ratio, CHLA = chl a, cc = Correlation coefficient | |
| Table 3: | Dependence of large zooplankton (adult copepods, nauplii and rotifers) and micro-zooplankton (protozoa) group abundance on cyanobacteria, green algae, diatoms, dinoflagellates as well as large zooplankton on micro-zooplankto |
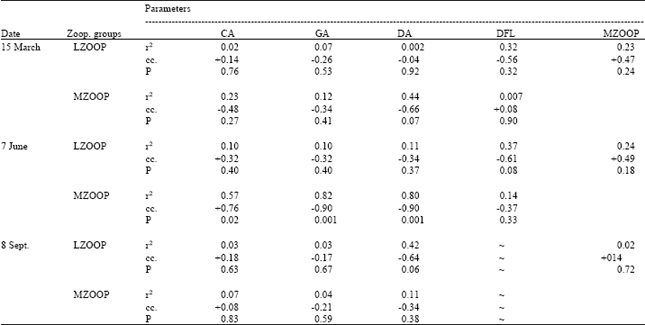 | |
LZOOP = large zooplankton, MZOOP = micro-zooplankton, CYN = cyanobacteria, GA = green algae, DA = diatoms, DFL = dinoflagellates, cc = correlation coefficient, ~ = no dinoflagellates were counted in that particular date | |
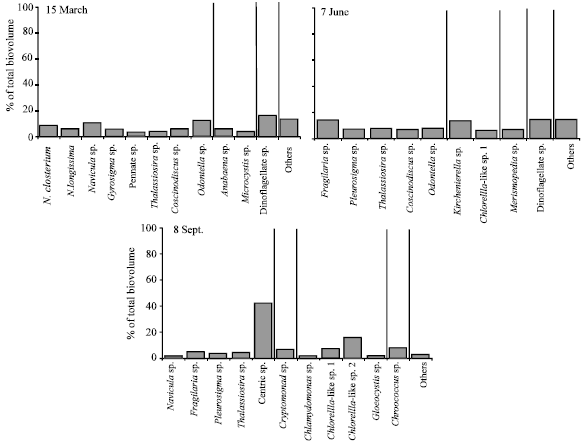 | |
| Fig. 4: | Phytoplankton community composition placed into dominant genera in March, June and September. Figure shows only the abundant groups in each month. Vertical lines separate the generic taxonomic groups as diatoms, green algae, cyanobacteria, dinoflagellates and others (a combination including less abundant genera) |
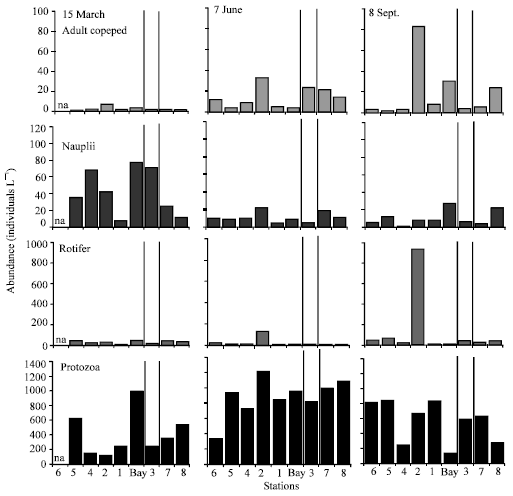 | |
| Fig. 5: | Adult copepod, nauplii, rotifer and protozoa abundance for March, June and September sampling. Data were not available for station 6 in March. The thin and thick vertical lines separate the stations that were in the pool areas (Fig. 1. St. 3, 7, 8) from the other areas |
Coccoid and oblong forms were dominant in March and June while Crucigenia, Scenedesmus, Tetraedron and Tetrastrum species dominated the green algae in September (in the group "Others" in Fig. 4). Cryptomonad sp. was also present in September. Peridinum and some other forms dominated the dinoflagellates while coccoid forms dominated the cyanobacteria. Both the abundance and composition of phytoplankton changed considerably between the stations (Fig. 3). Finally, phytoplankton species diversity was higher in September when compared to March and June (Fig. 3).
In March, poor correlation between N:P and cyanobacteria, green algae, diatoms and dinoflagellates were observed system wide (Table 2). Except diatoms, cyanobacteria, green algae and dinoflagellates showed relatively weak relations with salinity (Table 2). In June, relatively weak correlation between N:P and cyanobacteria, green algae, diatoms and dinoflagellates were prevalent. Moderately strong correlation was observed between salinity, diatoms and dinoflagellates at this sampling (Table 2). In September, except cyanobacteria all other variables showed strong relations with N:P and salinity (Table 2)
Similar to phytoplankton composition, zooplankton composition and abundance changed considerably between stations (Fig. 5). Adult copepods and rotifers were more abundant in September when compared to March and June. There were moderately strong relations between micro-zooplankton and diatoms as well as large zooplankton and dinoflagellates in March (Table 3). Strong correlation between large zooplankton and diatoms, micro-zooplankton and cyanobacteria, green algae and diatoms were observed in June (Table 3). In September, except the moderately strong relation between large zooplankton and diatoms, large and micro-zooplankton showed relatively weak correlation with phytoplankton groups (Table 3).
DISCUSSION
Field data showed spatial variability among sampling dates. Although Si concentration was always high in the system, import of new Si due to heavy rain events caused a dramatic increase in Si concentration in September. This is because unlike other nutrients the main source of Si to an aquatic system is via weathering of silicate bearing rocks through runoff[6]. N concentration was low and DIN:P was much less than Redfield stoichiometry throughout the stations in all sampling times. Similar to Si, heavy rainfall and flooding caused an increase in N and P concentrations in September.
The study system was nitrogen-limited and phosphorus sufficient at the time of sampling in March and June. This favored some k-selected species. These included Anabaena spp., Peridinum spp. and Euglena sp. Representatives from these genera are typically slower growing and less edible[14-17]. As a result of increased freshwater inflows following heavy rains at the time of sampling in September, r-selected species of green algae and diatoms dominated the system. Typically, r-selected species are characteristic of higher maximum growth rates relative to k-selected species[18-20].
Numerically, adult copepods and nauplii dominated the macro-zooplankton in March and June. Rotifers were more abundant in September. Copepod adults and nauplii can graze not only on the phytoplankton that is susceptible to rotifer grazing but also on larger phytoplankton species[20,21]. Additionally, previous studies have shown that copepod adults can graze on rotifers and also protozoa[21,22]. Therefore, grazing by adult copepods may likely contribute to the lower abundance of rotifers and protozoa in March and lower abundance of rotifers in June. As mentioned previously, field sampling was conducted shortly after heavy rain events in September. Low salinity and high nutrient concentration lead to an increase of rapidly-growing and edible phytoplankton forms, which often have smaller cell size and follow favorable nutrient perturbations[4,20,23]. This likely favored rotifers, which reproduce more rapidly than copepods and prefer prey of smaller cell size[6,20].
The natural plankton community structure in a tidal creek of the Nueces Delta was diverse and community structure changed due to physical processes, i.e., rain events. Nutrient loading due to additional freshwater input to the system caused an increase in more edible forms of phytoplankton, which in turn affected the zooplankton structure. This study, one more time, confirmed that freshwater input to a wetland is important to maintain the environmental health of a marsh ecosystem.
ACKNOWLEDGMENTS
This study was funded, in part, by the Texas A&M Sea Grant College Program (DOC-NOAA), award number NA86RG0058, Texas Water Resources Institute (TWRI), award number USGS-502229-5232 and USDA/NRI, award number 00-35101-9275.