
H. Okan Kamaci
Department of Aquaculture, Faculty of Fisheries, Ege University,
35440, Uria-lskele, In-nir, TURKEY
Kursat Firat
Department of Aquaculture, Faculty of Fisheries, Ege University,
35440, Uria-lskele, In-nir, TURKEY
Sahin Saka
Department of Aquaculture, Faculty of Fisheries, Ege University,
35440, Uria-lskele, In-nir, TURKEY
Musa Bulut
Department of Aquaculture, Faculty of Fisheries, Ege University,
35440, Uria-lskele, In-nir, TURKEY
Journal of Applied Sciences
Year: 2005 | Volume: 5 | Issue: 3 | Page No.: 546-552







ABSTRACT
The embryonic development of sharpsnout seabream (Diplodus puntazzo Cetti, 1777) eggs was investigated in detailed for culture conditions. The experiments have been carried out at 24 ± 0.2 ° C. Eggs of sharpsnout seabream were obtained by wild broodstock without hormonal treatment. The average diameters of the eggs and oil globule were determined 0.807 ± 0.008 mm and 0.245 ± 0.008 mm, respectively. Embryonic development stages were observed in every 15 min. Whenever there was an evident difference during the embryonic development, the photographs were taken before or after the due time. Early morula stage, early gastrula stage, 2 somit stage and formation of otolith of eggs were determined 03:00 h, 07:10 h, 15:15 h and 19:30 h after fertilization, respectively. At 24 ± 0.2 ° C, hatching occurred 26:10 h after fertilization.
PDF Abstract XML References Citation
How to cite this article
H. Okan Kamaci, Kursat Firat, Sahin Saka and Musa Bulut, 2005. Determination of Embryonic Development Stages of Sharpsnout Seabream (Diplodus puntazzo Cetti, 1777) Eggs in Rearing Conditions. Journal of Applied Sciences, 5: 546-552.
DOI: 10.3923/jas.2005.546.552
URL: https://scialert.net/abstract/?doi=jas.2005.546.552
DOI: 10.3923/jas.2005.546.552
URL: https://scialert.net/abstract/?doi=jas.2005.546.552
INTRODUCTION
Sharpsnout sea bream is a commercially important species which lives in rocky bottoms. This species distributed in the Mediterranean and the Black Sea, as well as in the Atlantic Ocean. Naturally spawning period (October-November) of sharpsnout seabream is earlier than Sparus aurata (November-March), Dicentrarchus labrax (February-March), Pagrus pagrus (February-April), Pagrus major and Dentex dentex (April-May), which is advantage for culture of this species. It is becoming increasingly important as a new species for aquaculture in Mediterranean and has attracted wide interest of many scientists in respect to its reproduction and physiology Micale et al.[1]; larval rearing Favaloro et al.[2]; morphological study Palma et al.[3]; nutrition and growth Katavic[4]. However, literature about its early life history is scarce or limited to embryonic and larval development under captivity Boglione et al.[5].
This study describes the embryonic development stages of sharpsnout seabream eggs in rearing conditions. The objectives were to describe the embryonic development stage, thus contributing to the identification of the planktonic stages and to the introduction of this high value species to commercial aquaculture.
MATERIALS AND METHODS
Sharpsnout seabream broodstock, 10 females (1.2±0.2 kg mean body weight) and 10 males (0.9±0.1 kg mean weight), were selected from wild breeders and stocked in 10 m3 tank with a seawater flow of 1.5 m3 per h. The fish were fed once a day to satiation at noon, with approximately equal amounts by weight of a moist pelleted breeder diet and cuttlefish (Sepia officialis) and shrimp (Palaemon elegans). No hormonal treatment was applied for breeder. After the first spawning, egg collectors of the broodstock tank were monitored and emptied every 15 min and eggs used in the study were obtained at 19:10 on September, 24, 2003. Spawning eggs were immediately collected and removed from recuparator. Following the fertilization, the viable buoyant eggs were separated from the dead sinking eggs. With the density 500 egg L-1, the eggs were placed of 10l incubators (Pyrex beakers) (mesh size, 300 μ), after statistical evaluations. The volumetric method was used to determine stocking of the eggs. Incubators placed in the tank of 1 m3 water volume. Water flow rate was adjusted in a way to change 15% of the total volume of the tank hourly. Temperature was kept at 24±0.2°C, and the salinity at a level of 37‰ during the experiment. Also, aeration rate was applied as 40 mL min-1.
Incubation took place in complete darkness and was triplicated. Diameter of egg and oil globule were measured to the nearest 0.025 mm using an ocular micrometer. To determine the common embryonic developments, 100 eggs were taken from each incubator and were investigated every fifteen minutes. Whenever there was an evident difference during the embryonic development, the photographs were taken before or after the due time. At the end of experiment all larvae were counted to determine for the survival rates. Each group were compared between them by Fischer Chi square (the significance test of the difference between two percentages in independent groups) and given within a 95% confidence interval. When the difference between the mean values was p>0.05, it was accepted as insignificant.
RESULTS
Spawning commenced on September, 3, 2003 and continued to October, 29, 2003, with a peak from September 17 to October 22, corresponding to temperatures ranging from 22 to 24.5°C. Oxygen concentration was changed approximately 6-7 mg L-1 and pH was around 7.9. Ammonia and nitrite were always <0.01 mg L-1. During the spawning season, the total number of eggs collected was estimated at 2.55 million, of which 0.48 million (18.83%) were sinking eggs and 2.07 million (81.17%) were buoyant eggs.
Sharpsnout eggs are buoyant, transparent and had a clean chorion. Egg diameters ranged from 0.801–0.816 mm with a mean of 0.807±0.008 mm. Eggs contained a single un-pigmented oil globule and were positively buoyant (Fig. 1-1). Diameters of oil globule changed about 0.245±0.008 mm to diameter. Fertilized eggs have a small perivitelline space. Most fertilized eggs floated the water surface at 37‰ salinity and temperature 24°C.
During the incubation, a two-cell stage was observed 1:10 h after the fertilization at 24±0.2°C. The two blastomeres are highly rounded and symmetrical just after cleavage. The dimensions of each blastomere were measured 206x291 μm. (Fig. 1-2). The second division occurred 1:20 h after the fertilization and then a four-cell stage was observed. The dimensions of each blastomere were measured 171x137 μm (Fig. 1-3). An 8-cell stage appeared after 1:30 h. The dimensions of blastomeres are changed between 103-132 μm (Fig. 1-4). 16-cell and 32-cell stages were observed at 2:10 and 2:30 h after fertilization, respectively and the dimensions of blastomeres are changed between 84-107 μm (Fig. 1-6). It was hard to observe the other symmetrical divisions, but they continued to divide. Early and late morula stage was observed after 3:00 h and 3:30 h after fertilization, respectively (Fig. 1-8).
Early blastula stage characterized that, nuclei from the marginal cells, migrate out of the cells and are distributed around the blastoderm and was determined 04:45 h after fertilization (Fig. 1-9). In late blastula stage, projection of the underside of the blastoderm into the yolk sphere is observed and several rows of periblast nuclei are visible around the blastoderm. Stage was observed 05:15 h after fertilization (Fig. 1-10).
06:15 h after fertilization, pre-early gastrula stage was determined and blastoderm has started flattened down (Fig. 1-11). Early gastrula stage was observed 07:15 h after fertilization. The blastoderm begins to expand over the surface of the yolk sphere and covered about 1/4 of egg. Dorsal margin of the blastoderm is thickened (Fig. 1-12). Pre-middle gastrula stage was observed 1:45 h later from early gastrula stage. Epiboly progressively advanced and the blastoderm covered about 1/3 of the yolk sphere (Fig. 2-1). The germ ring is well defined. Middle gastrula stage was determined 11:15 h after fertilization (Fig. 2-2). A streak is visible in the midline of the embryonic shield projecting into the germ ring area. The blastoderm covered about ½ of the yolk sphere. 12:30 h after fertilization, late gastrula stage was observed (Fig. 2-3). The blastoderm covered 3/4 of the yolk sphere and the embryonic shield became more clearly visible as a narrow streak.
Early naurula stage was observed 13:15 h after fertilization (Fig. 2-4). The yolk sphere was nearly covered by the thin blastoderm leaving a small area around the vegetal pole exposed. The head was recognized anteriorly in the distinct embryonic body. Thickness of head and body are same (0.85 μm) and internode was observed between head and body. A beak-like mass of cells was seen in front of the head. Kupffer's vesicles were appeared at the underside of the caudal (posterior) end of the body, which was in contact with a small blastopore. Late neurula stage was determined 14:45 h after fertilization (Fig. 2-5). The head is arrow-shaped and solid optic buds were appeared on each side of the cephalic end. The beak- like cell mass was still visible in front of the head and a small blastopore formed at the vegetal pole of the yolk sphere.
14:45 h after fertilization, a groove appeared in the dorsal area of each optic lobe and blastopore completely closed. Two somits formed indistinctly on embryo body.
 | ||||
| Fig. 1: | Embryonic development of sharpsnout seabream eggs (1-12) | |||
| 1: Activated egg stage, | 2: 2-cell stage, | 2: 2-cell stage, | 4: 8-cells stage, | |
| 5: 16 cell stage, | 6: 32 cell stage, | 7: Early morula stage, | 8: Late morula stage, | |
| 9: Early blastula stage, | 10: Late blastula stage, | 11: Pre-early gastrula stage, | 12: Early gastrula stage | |
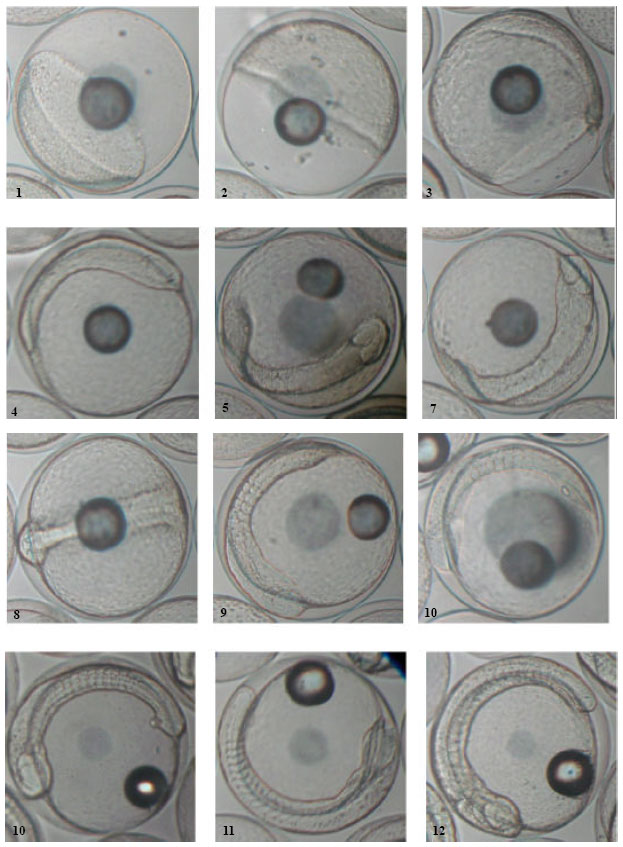 | ||||
| Fig. 2: | Embryonic developement of sharpnout seabream eggs (1-12) | |||
| 1: Pre-middle gastrula stage | 2: Middle gastrula stage | 3: Late gastrula stage | 4: Early naurula stage | |
| 5: Late naurula stage | 6: 2 somite stage | 7: 4 somite stage | 8: 6 somite stage | |
| 9: 9 somite stage | 10: 12 somite stage | 11: 16 somite stage | 12: 18-19 somite stage | |
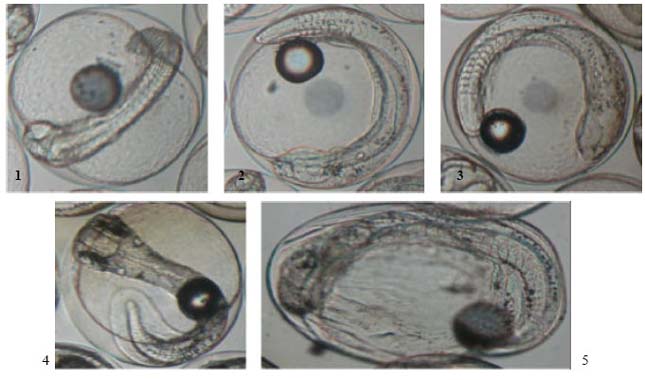 | |||
| Fig. 3: | Embryonic development of sharpsnout seabream eggs (1-5) | ||
| 1: 22 somite stage | 2: Appearance of otolith | 3: Embryo 3/4 | |
| 4: Repulsive movement | 5: Solving of chorion | ||
Thirty min after formation of two somits (Fig. 2-6), 4 somits were observed 15:45 h after fertilization (Fig. 2-7). 6 somits were determined 16:15 h after fertilization (Fig. 2-8). Optic vesicle was completely formed and departed from head region. Embryo covered ½ of yolk-sac. 9 somits and 12 somits were determined 17:00 h and 17:30 h after fertilization, respectively (Fig. 2-9, 10). The formation of tubular heart started and the projection part formed on hearth region as stripe-shape. The tubular heart (heart anlage) appears underneath the head from the posterior end of the mid-brain to the anterior end of the hind-brain.
Sixteen somits were determined 18:00 h after starting of incubation (Fig. 2-11). In this stage, disappearance of Kupffer's vesicles was observed. Also, formation of primordial fin started and was observed slightly visible. 18-19 somits were determined 18:30 h after fertilization (Fig. 2-12). Optic lens was observed on optic vesicle and also, Embryo covered 2/3 of yolk-sac. Otoliths were observed visible 15:45 h after fertilization. Development increased at heart region. 19:00 h after fertilization, 22 somits was determined (Fig. 3-1). Otoliths were observed clear and caudal fin determined independent from yolk-sac 19:30 h after fertilization (Fig. 3-2). 03:00 h later, embryo covered ¾ of yolk-sac (Fig. 3-3). Activity of embryo started.
Embryo was observed too active 25:15 h after fertilization and head region extremely developed. Embryo put pressed on chorion with sinusoidal movements. 26:10 h after fertilization, the embryos dissolved the inner layers of the chorion and tore the single outer layer by moving the body and escaped from the chorion head-first (Fig. 3-4). The large part of the body which started in the egg chorion got stable about 1-2 mm. Then, embryo slowly hatched out from the head to the oil globule by shaking movement (Fig. 3-5). After living of oil globule from chorion, embryo hatched out from the egg by shortly and/or distinctly swimming behaviour. After hatching, body of embryo was curve-shaped. Body was plane-shaped between 5-30 min after hatching out the chorion.
The mean total length of newly hatched larvae was 1.908±0.037 mm with an ellipsoid yolk-sac of 0.913±0.089 mm long and 0.519±0.056 mm wide laterally. An oil globule of 0.245±0.008 mm diameter was present at the posterior end of the yolk-sac. The yolk-sac larvae had unpigmented eyes and no fins and the mouth was not formed. Heart was visible with rhythmic heartbeat.
Hatching rates were determined between 84, 87 and 91% in incubators at 24±0.2°C. There were no significant statistically differences on embryonic development stages and hatching rates between incubators at 24±0.2°C (p>0.05).
| Table 1: | Comparison of data for egg and yolk-sac larvae of different sparid species |
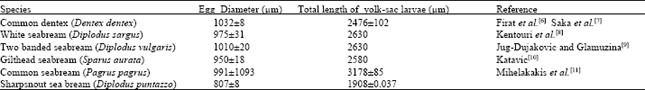 | |
DISCUSSION
The ripe sharpsnout seabream egg is a typical sparidae egg. It is difficult to identify morphological characteristics by which the eggs of this species differ from those of common dentex, Dentex dentex, white seabream, Diplodus sargus, two banded seabream, Diplodus vulgaris, gilthead seabream, Sparus aurata, or common seabream, Pagrus pagrus. Therefore it need more information for identifying and selection of egg to icthyoplaxtonic studies in survey.
Egg diameter and total length of newly hatched yolk-sac larvae of six sparidae species are compared in Table 1. The eggs of sparidae can theoretically be identified by egg diameter and oil globule. However, this is not very safe in practice as egg size can differ among females of the same species and might be thought that natural conditions (temperature, food, geographic location) and broodstock management (female age, spawning time regulation, salinity, temperature, nutrition) in captivity.
The length of newly hatched yolk-sac larvae of sharpsnout seabream was significantly less than those of the other five species. This information may assist in identifying newly hatched yolk-sac larvae; however, it would be of limited value since during the first few hours after hatching yolk-sac larvae developed very rapidly and the length changed very quickly.
In this study, oxygen, salinity, ammonia and nitrite differences were kept within the values at which embryonic developments in nature usually occurred and thus their adverse effects on the incubation of the eggs were tried to be prevented. The minimum oxygen rate in the incubators was determined as 6.4 mg L-1 and maximum 7.2 mg L-1. It has been reported that oxygen levels between 6-7 mg L-1 in the incubation of marine fish eggs have affected the embryonic development in a positive way[12,13]. In this study, the oxygen level in all the incubators was kept within specified limits.
The early development of fish is strongly affected by incubation temperature. Generally, lower temperature retards the rate of embryonic development of fish and higher temperature accelerates it. Production of viable larvae is a better indicator of temperature effect than total hatch because total hatching rate, which includes abnormal larvae, does not provide the information required for prediction of the percentage of larvae that may achieve exogenous feeding and successive normal development. The temperature levels changed for 23.9-24.1°C during incubation.
In this study, the hatching rates were determined between 82-86%. When these results were compared with hatching rate of S. aurata Katavic[10], P. major Mihelakakis and Yoshimatsu[14] and D. labrax Saka et al.[12], similar results were observed.
Culture of sharpsnout seabream was reached important point in the last decade, in Mediterranean fish culture basin. Nevertheless there are not any studies about sharpsnout seabream culture in Turkey. However, this study was to describe the detailed embryonic development of common dentex eggs and assist in the identification of planktonic stages, thus these photograph series provide certain results to hatcheries taking eggs from different farms and supply them with information on which period the eggs are and when they are expected to hatch.
REFERENCES
- Micale, V., F. Perdichizzi and G. Basciano, 1996. Aspects of the reproductive biology of the sharpsnout seabream Diplodus puntazzo (Cetti, 1777). 1. Gametogenesis and gonadal cycle in captivity during the third year of life. Aquaculture, 3: 281-291.
CrossRefDirect Link - Palma, J. and J.P. Andrade, 2002. Morphological study of Diplodus sargus, Diplodus puntazzo and Lithognathus mormyrus (Sparidae) in the Eastern Atlantic and Mediterranean Sea. Fish. Res., 57: 1-8.
CrossRefDirect Link - Boglione, C., M. Giganti, C. Selmo and S. Cataudella, 2003. Morphoecology in larval fin-fish: A new candidate species for aquaculture, Diplodus puntazzo (Sparidae). Aquac. Int., 11: 17-41.
CrossRefDirect Link - Firat, K., S. Saka and D. Coban, 2003. The effect of light intensity on early life development of the common dentex (Dentex dentex L. 1758) larvae. Aquac. Res., 34: 727-732.
Direct Link - Mihelakakis, A., T. Yoshimatsu and C. Tsolkas, 2001. Spawning in captivity and early life history of cultured red porgy, Pagrus pagrus. Aquaculture, 199: 333-352.
CrossRefDirect Link - Mihelakakis, A. and T. Yoshimatsu, 1998. Effects of salinity and temperature on incubation period, hatching rate and morphogenesis of the red sea bream. Aquac. Int., 6: 171-177.
CrossRefDirect Link