
Khurshid Anwar
Department of Zoology, University of Azad Jammu and Kashmir,
Muzaffarabad Azad Kashmir, Pakistan
Journal of Applied Sciences
Year: 2003 | Volume: 3 | Issue: 5 | Page No.: 317-330







ABSTRACT
Present study was aimed at investigating the toxic effects of permethrin on the kidney of newly hatched chicks developed from the eggs injected with a single sublethal doses (0.05 ml) of three different concentrations of permethrin insecticide (50, 100 and 200 ppm) on day ‘0 ’of incubation. Study included the estimation of a few enzyme activities and some biochemical constituents of kidney. Among enzymes, the activities of alkaline phosphatase, acid phosphatase, glutamate oxaloacetate transaminase and glutamate pyruvate transaminase were decreased, whereas, the activity of lactate dehydrogenase was increased. However, the activity of amylase remained unchanged. From among biochemical components glucose, glycogen, total protein, total lipids, cholesterol and uric acid contents elevated significantly at different dose levels whereas DNA and RNA contents were significantly reduced. Soluble protein, free amino acids (FAA) and urea contents remained unaltered. The common histopathological changes observed in the kidney sections from 100 and 200 ppm of permethrin-treated animals were tubular necrosis, reduction in the size of glomeruli as well as complete distortion of glomeruli in some sections accompanied by blood cell infiltration. Endothelial layer of Bowmann`s capsule and epithelial layer of glomeruli also showed severe damage. Histopathological changes were not observed in the kidney sections from the animals treated with 50 ppm of permethrin.
PDF Abstract XML References Citation
How to cite this article
Khurshid Anwar, 2003. Effect of Permethrin Treatment on the Kidney of Newly Hatched Chick (Gallus
domesticus). Journal of Applied Sciences, 3: 317-330.
DOI: 10.3923/jas.2003.317.330
URL: https://scialert.net/abstract/?doi=jas.2003.317.330
DOI: 10.3923/jas.2003.317.330
URL: https://scialert.net/abstract/?doi=jas.2003.317.330
INTRODUCTION
Development and growth of domestic chicks, which are the rich source of animal protein in the form of eggs and meat can be affected by the insecticides. Insecticides from hens, fed on contaminated feed, can be transported to young embryos through eggs and thus can cause severe teratological abnormalities, biochemical changes, organ dysfunction and mortality in the young embryos.
Pyrethroid insecticides, including permethrin are used against pests to increase the production of food grains and other agricultural-products like cotton, vegetable oils, vegetables and fruit etc. They also produce deleterious effects on the non-target organisms including both invertebrates and vertebrates living in the exposed area (Sibley and Kaushik, 1991 and Majmunder et al., 1994). A lot of information is available on the toxicity of insecticides, other than pyrethroids in chick embryo (Mufti and Nasim, 1987), whereas, a little information is available about the toxicity of pyrethroids including permethrin on the development of chicks (Kapoor et al., 1988).
Permethrin (3-(2,2-dichloro-ethenyl)-2,2-dimethylcyclopropanecarboxylic acid-(3-phenoxyphenyl) methylester), being a most promising pyrethroid is photostable and possess high insecticidal activity. Permethrin is widely used in Pakistan and there is increased risk of food being contaminated with the insecticides. In addition, permethrin is also used as prophylactic agent against scabies in humans (Chouela et al., 2002).
Many workers have undertaken the toxicological studies of permethrin on chicks (Ferguson and Audesirk, 1990). Qadri et al. (1987) observed the acute toxicity of permethrin to hemoglobin RBC count and chloride level in chick blood. Permethrin is also known to damage liver in newly hatched chicks (Anwar, 2003) In mammals including humans it also causes deleterious effects. In mice permethrin has been found to depress several immune responses in isolated splenic leukocytes including splenic T-cell proliferative response to mitogen, splenic macrophage hydrogen peroxide production and splenic B lymphocyte-specific antibody production (Prater et al., 2003). Permethrin is known to cause the release of inflammatory cytokine, Interleukin-8 (IL-8) from human epidermal kinetocytes (Monteiro- Riviere et al., 2003). Kostka et al. (1997) observed the induction of CYP 2B and slight increase in CYP 1A in rats treated with 620 mg kg-1 of permethrin. Permethrin is known to cause neurotoxicity in mammals (Abdel-Rehman et al., 2001 and Karen et al., 2001). Sheets (2000) observed that young rats are more sensitive than old rats at lethal dose to pyrethroids and this greater susceptibility of the neonates to pyrethroids appears to be due to the limited metabolic capacity. Imamura et al. (2002) found that permethrin represses the induction of activity-dependent c- fos and brain-derived neurotrophic factor (BDNF) gene expression, accompanying a decrease in Ca (2+) influx into neurons in neonate mice. Spencer and Berhane (1982) observed the fetal changes in rats caused by permethrin.
Toxins are metabolised via mixed function oxidase system. Kapoor et al. (1988) studied the effect of permethrin on the mixed function oxidase system and found that permethrin induces microsomal protein, cytochrome P-450 and NADPH cytochrome C reductase in chicks in dose dependent manner and also observed permethrin as a weak inducer of hepatic microsomal mixed function oxidases in chicks fed Vitamin A deficient diet. Heder et al. (2001) found that permethrin induces CYP2B1 in primary rat hepatocyte cultures. The most common metabolites of permethrin found in rat plasma and urine are m-phenoxybenzoic acid and m-phenoxybenzoyl alcohol (Abu-Qare and Abou-Donia, 2001). Akhtar and Mahadevan (1992) have already shown that 3-phenoxy benzoic acid, the major metabolite of several pyrethroid insecticides is rapidly metabolized by chicken kidney microsomal preparations. Since kidney is also involved in drug metabolism, so it can be vulnerable to toxic insult caused by the insecticides. In addition, kidney plays a pivotal role in the excretion of nitrogenous wastes produced in the body as well as maintains body electrolytes balance and is also involved in the synthesis of various hormones. Damage to kidney can result in the accumulation of toxic compounds in the body i.e urea and uric acid, causes disturbance in water-elctrolyte balance, which in turn can lead to the death of whole organism. A very little information is available on the toxicity of permethrin on the development of kidney in chicks. So the present study was designed to evaluate the toxic effects of permethrin on the development of kidney in newly hatched chicks at both biochemical and histological levels.
MATERIALS AND METHODS
Chicks were developed from fertilized eggs (thirty) obtained from Government Poultry Farm, at Muzaffarabad Azad Kashmir, Pakistan, injected with different doses of permethrin insecticide. Dilutions of permethrin were made in acetone. LD50 was obtained using probit analysis. A single sublethal dose (0.05 ml) of the insecticide of each concentration (50, 100 and 200 ppm) was administered through injection to 4 groups (atleast 4 eggs in each group) at day '0' of incubation. Insecticide was administered into the yolk of each egg at vegetal pole by disposable tuberculin syringes. Equal volume of acetone was injected into the controls. The eggs were incubated at 38 ± 0.5°C in incubators with a relative humidity of 70% with proper ventilation. The eggs were rotated every two hours to avoid the sticking of the embryo to the shell membranes.
On the day of hatching the kidneys from each chick was taken out, weighed and divided into three parts. One part was used for making saline homogenate, while the other part was used for the extraction of lipid, cholesterol and nucleic acids. The third part was fixed in Bouin’s fluid for light microscopical studies. Saline homogenate was made in ice-cold 0.89% saline using motor driven Teflon glass homogeniser and used for the estimation of various enzyme activities and some other biochemical components.
The activities of aspartate aminotransferase (ASAT; L, aspartate: 2 oxoglutarate aminotransferase, EC 2:6:1:1) and alanine amino transferase (ALAT; L, alanine: 2 oxoglutarate aminotransferase (EC 2:6:1:2) by the method of Reitman and Frankel (1957). The activities of alkaline phosphatase (AkP, orthophosphoric monoester phosphohydrolase, alkaline optimum, EC: 3:1:3:1) and acid phosphatase (AcP, orthophosphoric monoester phosphohydrolase, acid optimum, EC: 3:1:3:2) were estimated according to the method of Kind and King (1954). Lactate dehydrogenase (LDH, L, lactate: NAD oxidoreductase (EC 1:1:1:27) activity was estimated by a method based on Cabaud and Wroblewski (1958). The amylase (1, 4 a-D glucanhydrolase, EC 3:2:1:1) activity was estimated according to the procedure described by Wootton (1964).
Soluble proteins were determined from saline tissue extract, while same saline extract was digested in 0.5N NaOH for 24 h and used for the estimation of total proteins. Both total and soluble proteins were estimated according to Lowry et al. (1951). Glucose content was estimated by O-toluidine method of Hartel et al. (1969). Glycogen content in the supernatant left after centrifugation of saline homogenate was precipitated with ethanol and then dissolved in distilled water and estimated by the Anthrone method of Consolazio and Lacono (1963). Amino acid contents were estimated according to the Ninhydrin method of Moore and Stein (1957). Estimation of urea was performed according to the DAM method as described by Natelson et al. (1951). Uric acid content was determined according to the method described by Carraway (1963).
For the extraction of total lipid and cholesterol, the tissue was ground in hot ethanol (60°C) and kept for extraction overnight. After centrifugation at 5,000 rpm for 10 minutes, the supernatant was obtained and used for the estimation of total lipid by Vanillin reagent (Zollner and Kirsch, 1962) and cholesterol content according to Liebermann and Burchardt Reaction (Henry and Henry, 1974). Nucleic acids were extracted according to the method described by Shakoori and Ahmed (1973). The pellet left during lipid extraction was used for preparation of DNA and RNA extracts. DNA was extracted in hot PCA and estimated according to diphenylamine method, while RNA extract was prepared in cold PCA and estimated according to the orcinol method. Both these estimations follow the procedure as described in Schneider (1957).
Instruments
Teflon Glass homogeniser (TRI-R STIR-R, Model S63C USA), UV Spectrophotometer (Model M 302, Camspec, England), Spectrophotometer (Sequola-Turner, Model 340, USA), Refrigerated Centrifuge (Sigma, Germany), Centrifuge (PHG Hermle Z 230, West Germany), Water Bath (LCB 800 NEDTEX Co Taiwan), Incubator (Memmert, West Germany) and Analytical Balance (Sartorius, West Germany).
Place of work
All the work was done in Biochemistry and toxicology Laboratory, Zoology Department, Azad Jammu and Kashmir University Muzaffarabad, Azad Kashmir.
RESULTS
Enzyme activities
Among enzymes, the activities of AkP, AcP, AST and ALT were decreased while the activity of LDH was increased. However, the activity of amylase remained unchanged. The activities of both the phosphatases AkP and AcP decreased at 200 ppm; the activity of AkP decreased by 50% and the activity of AcP by 27%. The activities of both the transaminases, ALT and AST were decreased at 100 and 200 ppm. The activity of AST decreased by 21% at 100 ppm and by 19% at 200 ppm. Similarly, the activity of ALT was decreased by 25% at 100 ppm and by 20% at 200 ppm. Activity of LDH increased at all dose levels: 81% at 50 ppm, 61% at 100 ppm and 58% at 200 ppm (Table 1 and 2).
Biochemical components
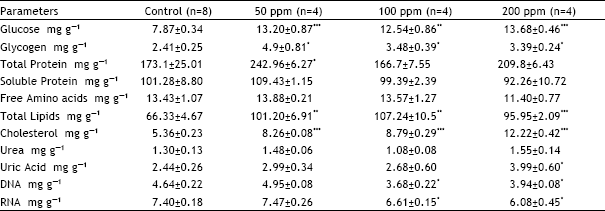
Table 3 and 4 show the effect of permethrin treatment on biochemical components of kidney. From among biochemical components glucose, glycogen, total protein, total lipids, cholesterol and uric acid contents elevated significantly at different dose levels, whereas, DNA and RNA contents were significantly reduced. Soluble protein, free amino acids (FAA) and urea contents remained unaltered. Both the glucose and glycogen contents increased at all the doses.
| Table 1: | Toxicological effects of a single treatment of Permethrin of various concentrations (50, 100 and 200 ppm) administered in to the eggs at ‘0’ day of incubation on some Enzymes Activities of Kidney of newly hatched chick |
 | |
| *= significantly different from controls at P < 0.05, using student ’t’’test | |
| **= significantly different from controls at P < 0.01, using student ’t’’test | |
| ***= significantly different from controls at P < 0.001, using student ’t’’test | |
| IU: International unit, the amount of enzyme which under defined assay conditions will catalyse the conversion of one micro mole of substrate per minute | |
| SoU: Somogyi Unit: The amount of enzyme that catalyses digestion of 5 mg of starch under the experimental condition | |
| KAU; King Armstrong Unit: The amount of enzyme that transforms 1 mg of phenol in 15 min. | |
| Table 2: | Percent change in the Enzymes Activities of Kidney of newly hatched chicks developed from eggs injected with a single dose of permethrin of various concentrations (50, 100 and 200 ppm) at ‘0’ day of incubation |
 | |
| Table 3: | Toxicological effects of a single treatment of permethrin administered into the eggs at ‘0’ day of incubation on some Biochemical Components of Kidney of newly hatched chicks |
 | |
| * significantly different from controls at P < 0.05, using student ’t’’test | |
| **, significantly different from controls at P < 0.01, using student ’t’’test | |
| ***, significantly different from controls at P < 0.001, using student ’t’’test | |
| Table 4: | Percent change in the Biochemical Components of Kidney of newly hatched chicks developed from eggs injected with a single dose of permethrin of various concentrations (50, 100 and 200 ppm) at ‘0’ day of incubation |
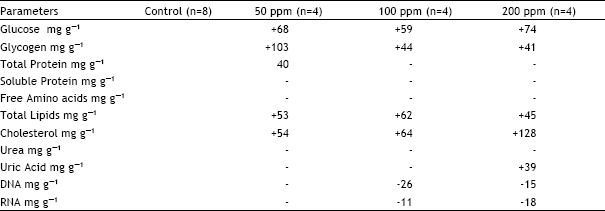 | |
Glucose content increased by 68, 59 and 74% at 50, 100 and 200 ppm, respectively. Glycogen content increased by 103% at 50 ppm, 44 at 100% and by 41% at 200 ppm. Increase in total protein content was observed only at 50 ppm and this increase was 40%. Total lipids and cholesterol contents increased at all the dose levels. Total lipids increased by 53, 62 and 45% at 50, 100 and 200 ppm, respectively. Cholesterol content showed increase of 54% at 50 ppm, 64% at 100 ppm and 128% at 200 ppm. Uric acid content showed increase only at 200 ppm and that increase was 39%. Both the DNA and RNA contents showed decrease at 100 and 200 ppm while at 50 ppm they remained unaffected by permethrin treatment. DNA content decreased by 26 and 15% and RNA by 11 and 18%, at 100 and 200 ppm, respectively.
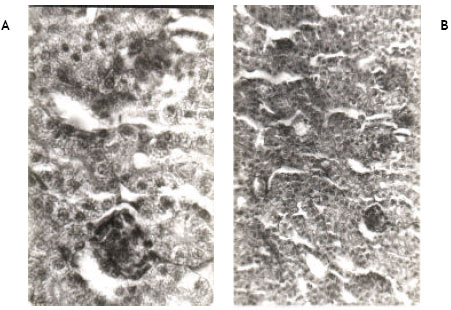 | |
| Fig. 1A, B: | Show the histology of the kidney of newly hatched chicks (Control) developed from the eggs injected with (0.2 ml) of vehicle acetone at ‘0’ day of incubation. Histological sections are normal |
 | |
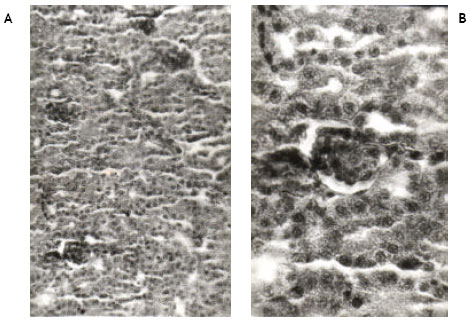
| Fig. 2A, B: | Show the histological changes in the kidney of newly hatched chicks developed from the eggs injected with 50 ppm (0.2 ml) of permethrin dissolved in acetone at ‘0’ day of incubation. No any histopathological change is observed |
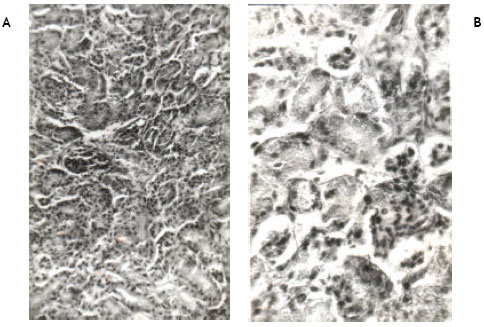 | |
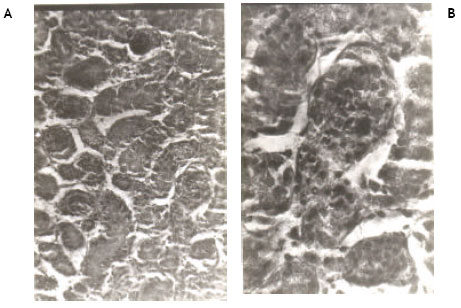
| Fig. 3A, B: | Show the histological changes in the kidney of newly hatched chicks developed from the eggs injected with 100 ppm (0.2 ml) of permethrin dissolved in acetone at ‘0’ day of incubation. Note the reduction in the size of glomeruli and tubular necrosis accompanied by blood cell infiltration. Some glomeruli are completely damaged. Damage to endothelial layer of Bowman’s capsule and epithelial layer of glomeruli is also seen |
 | |
| Fig. 4A, B: | Show the histological changes in the kidney of newly hatched chicks developed from the eggs injected with 200 ppm (0.2 ml) of permethrin dissolved in acetone at ‘0’ day of incubation. Note the reduction in the size of glomeruli and a few necrotic cells in renal tubules |
Histopathological changes
Light microscopic examinations of hematoxylin and eosin stained kidney sections from the control chicks revealed normal appearance (Fig. 1A, B). Permethrin at 50 ppm did not cause any pathological change in the kidney (Fig. 2A, B). However, permethrin treatment at higher doses of 100 and 200 ppm induced some histopathological changes including tubular necrosis and glomerulitis. At 100 ppm, there was reduction in the size of glomeruli and tubular necrosis accompanied by blood cell infiltration. Both proximal and distal tubules were damaged. Kidney sections at higher magnification reveal complete distortion of some of the glomeruli. Endothelial layer of Bowmann’s capsule and epithelial layer of glomeruli were severely damaged. Necrosis of epithelial cells of the tubules was prominent as evidenced by disappearance of nuclei in these cells (Fig. 3A, B). Kidney sections from the animals treated with 200 ppm of permethrin revealed the reduction in the size of renal corpuscles as well as the presence of a few necrotic cells in the tubules (Fig. 4A, B).
DISCUSSION
In addition to liver, kidney also plays an important role in drug metabolism (Akhtar and Mahadevan, 1992). These authors showed that 3-phenoxy benzoic acid, the major metabolite of several pyrethroid insecticides is rapidly metabolized by chicken kidney microsomal preparations. Since kidney is also involved in drug metabolism, so it can be vulnerable to toxic insult caused by the insecticides. In addition, kidney plays a pivotal role in the excretion of nitrogenous wastes produced in the body as well as maintains body electrolytes balance and is also involved in the synthesis of various hormones. Damage to kidney can result, in the accumulation of toxic compounds in the body, disturbance in water-elctrolyte balance which in turn can lead to the death of whole organism. In kidney damage, the urea, which is not excreted, retains in the blood and causes hemolysis of red blood cells. Present study was undertaken to see whether permethrin exerts its toxic effects on the development of kidney and damages it. Toxicity of permethrin was evaluated in terms of biochemical (estimation of a few enzyme activities and some biochemical components) and histological changes.
Enzyme activities
Of the enzyme activities, the activities of AkP, AcP, AST and ALT were decreased while the activity of LDH was increased. However, the activity of amylase remained unchanged. The activities of both the phosphatases AkP and AcP decreased at 200 ppm, AkP is membrane bound enzyme, it is found on all cell membranes where active transport occurs and is hydrolase and transphosphorylase in function. This decrease in AkP activity may be taken as an index of parenchymal damage (Onikienko, 1963). Role of Alkaline phosphatase activity had also been implicated in the development of the brush border of renal tubules (Washstein, 1955). Since AkP is also known to be involved in the active transport across the cell membrane (Kaplan, 1972) and transfer of metabolites across cell surface (Zamboni and Merchant, 1973), its decrease in the renal tissue may reflect the decreased function of the kidney. Therefore, decrease in the AkP activity can be attributed to the renal damage in the developing chicks as a result of permethrin toxicity. AcP is a lysosomal enzyme (de Duve and Wattiaux, 1966) and is used as an indicator of cell death, its decrease in the renal tissue may be due to its release from the tissue in to the blood stream as a result of tissue damage. The activities of both the transaminases, ALT and AST were decreased at 100 and 200 ppm. Worowski and Farbiszewski, (1970) have associated increases in serum transaminase activities with nephrotoxicity. Increase in serum transaminase activity occurs by the leakage of the enzymes from the damaged tissue. Decrease in the activities of these enzymes also reflect the damage to the renal tissue by permethrin. Activity of LDH increased only at 100 and 200 ppm. Recent findings that apoptosis, a programmed cell death, in addition to necrotic cell death, plays an important role in cell/tissue damage have made revolutionary changes in understanding the mechanism of tissue damage, which occur both by endo and exotoxins. El-Hassan et al. (2001) observed involvement of mitochondria in the paracetamol-induced hepatocytic apoptosis. Though this study was performed on liver but it still indicates the involvement of mitochondria in the cells undergoing apoptosis. Damage to mitochondria may result in the accumulation of lactic acid, which in turn can induce the increased synthesis of the LDH enzyme to convert lactic acid back to pyruvic acid, an initial substrate of the Kreb’s cycle. So the increase in LDH activity can be attributed to the hypoxic damage to the cells of renal tubules as well as renal corpuscle.
Biochemical changes
Table 3 and 4 show the effect of permethrin treatment on biochemical components of kidney. From among biochemical components glucose, glycogen, total protein, total lipids, cholesterol and uric acid contents elevated significantly at different dose levels whereas DNA and RNA contents were significantly reduced. Soluble protein, free amino acids (FAA) and urea contents remained unaltered. Both the glucose and glycogen contents increased at all the doses. Increase in glucose and glycogen content indicates less demand for the energy by the kidney as a result of tubular damage. Since energy is utilized for the active transport of ions across the cell membrane, the non-utilization of energy that have resulted in glucose and glycogen content can be attributed to the membrane damage as evidenced by the decrease in AkP activity, a membrane bound enzyme. Increase in total protein content might have occurred to repair the injured tissue. The evidence for the permethrin-induced damage to kidney is further supported by the reduced renal DNA content as observed in the present study. Abu-Qare and Abou-Donia, (2000) observed oxidative DNA damage in rats as a result of permethrin toxicity. Increase in total lipid and cholesterol contents may be due to fatty change in the kidney. Increase in the renal uric acid content indicates also indicates the renal damage. Decrease in both the DNA and RNA contents also indicates tissue damage.
Histopathological changes
In the present study, tubular necrosis, reduction in the size of glomeruli, degeneration of glomeruli as well as blood cell infiltration have been observed in the kidney sections from the animals treated with permethrin (100 and 200 ppm). Such changes in chick kidney have also been observed by Bhattacharya. et al. (1993) as a result of endosulfan treatment Fragmentation of glomerular capillaries and tubular nephrosis has been observed in chick kidney with Ochratoxin treatment (Subramanian et al., 1986).
Hydropic degeneration and necrosis occurs as a result of disturbance in Ca2+ homeostasis. Mitochondria have a key role in the regulation of intracellular calcium (Ca2+) (Carafoli, 1987) and the disturbance of mitochondrial function is linked with increased Ca2+ and hence multiple downstream effects in necrosis (Trump and Berezesky, 1992). So in the present study tubular necrosis might have resulted from disturbance in Ca2+ homeostasis. In addition, both direct and indirect evidences support another kind of cell death, the programmed cell death or apoptosis occurring as a result of permethrin toxicity. Prater et al. (2002) observed permethrin-induced apoptosis in CD4(-)8(-) and CD4(-)8(+) thymocytes in mice. Abu Qare and Abou-Donia (2003) reported the release of cytochrome c from mitochondria with permethrin. The role of cytochrome c in the induction of apoptosis has been recently investigated (Bossy-Wetzel and Green, 1999). Nuclear condensation and DNA fragmentation is the hallmarks of apoptosis (Ray et al., 1996). Nuclear condensation has been observed in the present study with permethrin toxicity. In light of these evidences it can be inferred that damage to renal tissue with permethrin treatment might have occurred through the induction of apoptosis.
Abu-Qare and Abou-Donia. (2001) observed the two metabolites of permethrin M-phenoxybenzoic acid and m-phenoxybenzoyl alcohol in rat plasma and urine. Kapoor et al., (1988) observed that permethrin induces the microsomal proteins cytochrome P-450 and NADPH reductase responsible for the bioactivation of permethrin. Akhtar and Mahadevan (1992) showed that 3-phenoxy benzoic acid, the major metabolite of several pyrethroid insecticides is rapidly metabolized by chicken kidney microsomal preparations. The metabolites of permethrin may be detoxified through conjugating with glutathione. glutathione-S-transferases (GSTs), a family of enzymes found at high levels in the liver that play an important role in the detoxification of electrophilic alkylating agents through conjugation (Mannervik and Danielson, 1988). Yin et al. (2001) observed the significant increase in Glutathione-s-transferase activity in Gammarus after 48h exposure of permethrin and that increase might have resulted to conjugate the reactive metabolites of permethrin with glutathione. Similar observations were made by Vontas et al (2001) who found Glutathione-s-transferases as an antioxidant defence against pyrethroid-induced lipid peroxidation, protein oxidation and depleted reduced glutathione in insects. Oxidative damage occurring through the release of cytochrome c from mitochondria has also been observed in rates following exposure to permethrin (Abu Qare and Abou-Donia, 2003). These data suggest that permethrin causes damage to the various organs through the generation of reactive oxygen species. In the present study the damage to renal tissue might have resulted from the oxidative damage of tissue macromolecules by reactive oxygen species generated during the metabolism of permethrin. Results of present experiment indicates that permethrin damages the kidney in developing chicks.
REFERENCES
- Abdel-Rehman, A., A.K. Shetty and M.B. Abou-Donia, 2001. Subchronic dermal application of N,N-diethyl-m-toluamide (DEET) and permethrin to adult rats, alone or in combination, causes diffuse neuronal cell death and cytoskeletal in the cerebral cortex and hippocampus and purkinje neuron loss in the cerebellum. Exp. Neurol., 172: 153-171.
PubMedDirect Link - Abu-Qare, A.W. and M.B. Abou-Donia, 2000. Increased 8-hydroxy-2'-deoxyguanosine, a biomarker of oxidative DNA damage in rat urine following a single dermal dose of DEET (N,N-diethyl-m-toluamide) and permethrin, alone and in combination. Toxicol. Lett., 117: 151-160.
PubMedDirect Link - Abu-Qare, A.W. and M.B. Abou-Donia, 2001. Simultaneous determination of chlorpyrifos, permethrin and their metabolites in rat plasma and urine by high-performance liquid chromatography. Anal. Toxicol., 25: 275-279.
Direct Link - Abu-Qare, A.W. and M.B. Abou-Donia, 2003. Combined exposure to DEET (N,N-diethyl-m-toluamide) and permethrin: Pharmacokinetics and toxicological effects. J. Toxicol. Environ. Health B. Crit. Rev., 6: 41-53.
Direct Link - Bossy-Witzel, E. and D.R. Green, 1999. Caspases induce cytochrome c release from mitochondria by activating cytosolic factors. J. Biol. Chem., 274: 17484-17490.
Direct Link - Cabaud, P.G. and F. Wroblewski, 1958. Colorimetric measurement of lactic dehydrogenase activity of body fluids. Am. J. Clin. Pathol., 30: 234-236.
PubMedDirect Link - Carafoli, E., 1987. Intracellular calcium homeostasis. Ann. Rev. Biochem., 56: 395-433.
PubMedDirect Link - Chouela, E., A. Abeldano, G. Pellerano and M.I. Hernandez, 2002. Diagnoses and treatment of scabies: A practical guide. Am. J. Clin. Dermatol., 3: 9-18.
PubMedDirect Link - De Duve, C. and R. Wattiaux, 1966. Functions of lysosomes. Ann. Rev. Physiol., 28: 435-492.
CrossRefDirect Link - Heder, A.F., K.I. Hirsch-Ernst, D. Bauer, G.F. Kahland and H. Desel, 2001. Induction of cytochrome P450 2B1 by pyrethroids in primary rat hepatocyte cultures. Biochem. Pharmacol., 62: 71-79.
CrossRefPubMedDirect Link - Imamura, L., H. Hasegawa, K. Kurashina, T. Matsunoand and M. Tsuda, 2002. Neonatal exposure of newborn mice to pyrethroid (permethrin) represses activity-dependent c-fos mRNA expression in cerebellum. Arch. Toxicol., 76: 392-397.
Direct Link - Kapoor, R.K., S.S. Chauhan, N. Sing and U.K. Misra, 1988. Induction of hepatic mixed-function oxidases by permethrin and cypermethrin in chicks fed vitamin A-deficient diet. Pestic. Biochem. Physiol., 32: 205-211.
CrossRefDirect Link - Kind, P.R.N. and E.J. King, 1954. Estimation of plasma phosphatase by determination of hydrolysed phenol with amino-antipyrine. J. Clin. Pathol., 7: 322-326.
CrossRefPubMedDirect Link - Kostka, G., D. Palut and B. Wiadrowska, 1997. The effect of permethrin and DDT on the activity of cytochrome P-450 1A and 2B molecular forms in rat liver. Rocz Panstw Zakl Hig., 48: 229-237.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Majmunder, S., A.K. Chakraborty, T.K. Mandal, A. Bhattacharya and K. Basak, 1994. Subacute toxicity of fenvalerate in broiler chicks: Concentration, cytotoxicity and biochemical profiles. Indian J. Exp. Biol., 32: 752-756.
PubMedDirect Link - Mannervik, B. and U.H. Danielson, 1988. Glutathione transferases-structure and catalytic activity. Crit. Rev. Biochem., 23: 283-337.
PubMedDirect Link - Monteiro-Riviere, N.A., R.E. Bavnes and J.E. Riviere, 2003. Pyridostigmine bromide modulates topical irritant-induced cytokine release from human epidermal keratinocytes and isolated perfused porcine skin. Toxicology, 183: 15-28.
PubMedDirect Link - Mbouangouere, R.N., P. Tane, D. Ngamga, P. Djemgou, M.I. Choudhary and B.T. Ngadjui, 2007. Piptaderol from Piptadenia africana. Afr. J. Traditional Complimentary Alternative Med., 4: 294-298.
Direct Link - Natelson, S., M.L. Scott and C. Beffa, 1951. A rapid method for the estimation of urea in biologic fluids. Am. J. Chem. Pathol., 21: 275-281.
PubMedDirect Link - Prater, M.R., Jr., R.M. Gogal, B.L. Blaylock and S.D. Holladay, 2003. Cis-urocanic acid increases immunotoxicity and lethality of dermally administered permethrin in C57BL/6N mice. Int. J. Toxicol., 22: 35-42.
CrossRefDirect Link - Prater, M.R., Jr., R.M. Gogal, B.L. Blaylock, J. longstreth and S.D. Holladay, 2002. Single-dose topical exposure to the pyrethroid insecticide, permethrin in C57BL/6N mice: Effects on thymus and spleen. Food Chem. Toxicol., 40: 1863-1873.
PubMedDirect Link - Qadri, S.S.H., K. Jabeen, M. Mahboob and M. Mustafa, 1987. Haemotoxicity to chicken (Gallus gallus-domesticus) by technical and formulation grades of some phosphoric and synthetic pyrethroid esters. J. Applied Toxicol., 7: 367-371.
PubMedDirect Link - Ray, S.D., V.R. Mumaw, R.R. Raje and M.W. Fariss, 1996. Protection of acetaminophen-induced hepatocellular apoptosis and necrosis by cholesteryl hemisuccinate pretreatment. J. Pharmacol. Exp. Ther., 279: 1470-1483.
PubMed - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Schneider, W.C., 1957. Determination of Nucleic Acids in Tissues by Pentose Analysis. In: Methods in Enzymology, Colowick, S.P. and N.O. Kaplan (Eds.). Vol. 3, Academic Press, New York, USA., pp: 680-684.
Direct Link - Sheets, L.P., 2000. A consideration of age-dependent differences in susceptibility to organophosphorus and pyrethroid insecticides. Neurotoxicology, 21: 57-63.
PubMedDirect Link - Sibley, P.K. and N.K. Kaushik, 1991. Toxicity of microencapsulated permethrin to selected nontarget aquatic invertebrates. Arch. Environ. Contam. Toxicol., 20: 168-176.
CrossRefDirect Link - Spencer, F. and Z. Berhane, 1982. Uterine and fetal characteristics in rats following a post-implantational exposure to permethrin. Bull. Environ. Contam. Toxicol., 29: 84-88.
CrossRefDirect Link - Vontas, J.G., G.J. Small and J. Hemingway, 2001. Glutathione-s-transferases as antioxidant defence agents confer pyrethroid resistance in Nilaparvata lugens. Biochem. J., 357: 65-72.
PubMedDirect Link - Washstein, M., 1955. Histochemical staining reactions of the normal functioning and abnormal kidney. J. Histochem., 3: 246-270.
Direct Link - Worowski, K. and R. Farbiszewski, 1970. Activity of aminotransferases and phosphatases in experimental poisoning of rabbits with mercuric chloride. Acta Physiol. Pol., 21: 385-392.
PubMedDirect Link - Yin, D., Jin, H.H. Yu and L. Chen, 2001. A comparative study on the sensitivity and specificity of cholinesterase and glutathione s-transferase in Gammarus pulex L. Ying Yong Sheng Tai Xue Bao., 12: 615-618.
PubMedDirect Link - Zamboni, I. and H. Merchant, 1973. The fine morphology of mouse primordial germ cells in extragonadal locations. Am. J. Anat., 137: 299-335.
PubMedDirect Link - Karen, D.J., W. Li, P.R. Harp, J.S. Gillette and J.R. Bloomquist, 2001. Striatal dopaminergic pathways as a target for the insecticide permethrin and chloropyrifos. Neurotoxicology, 22: 811-817.
CrossRef