
Zelalem Getnet
Department of Biology, College of Natural and Computational Sciences, University of Gondar, P.O. Box 196, Gondar, Ethiopia
Azamal Husen
Department of Biology, College of Natural and Computational Sciences, University of Gondar, P.O. Box 196, Gondar, Ethiopia
Masresha Fetene
Department of Plant Biology and Biodiversity Management, College of Natural Sciences, Addis Ababa University, P.O. Box 3434, Addis Ababa, Ethiopia
Gietahun Yemata
Department of Plant Biology and Biodiversity Management, College of Natural Sciences, Addis Ababa University, P.O. Box 3434, Addis Ababa, Ethiopia
Journal of Agronomy
Year: 2015 | Volume: 14 | Issue: 4 | Page No.: 188-202







ABSTRACT
Drought is a major limiting factor for crop expansion. Currently efforts are focused on improving crop genotypes for drought-prone areas. Considering this, a field experiment was conducted in the Shewa robit (a drought-prone area), Amhara Regional State, Ethiopia to evaluate the various growth attributes, physiological, biochemical and agronomic performances of different Stay-Green (SG) Sorghum accessions. Three Sorghum collections (Sorcoll) namely, Sorcoll 141/07, Sorcoll 146/07, Sorcoll 163/07 and a check (Afeso) genotypes were used. The experiment was arranged in a randomized complete block design. Data on the height, girth diameter, number of tillers, number of green leaf, leaf senescence, leaf area, chlorophyll content index, flowering, physiological maturity, Relative Water Content (RWC), net photosynthetic rate (Pn), transpiration rate (E), Water Use Efficiency (WUE), maximum quantum yield of PSII efficiency (Fv/Fm), leaf pigments, proline, nitrogen, protein, soluble sugar contents, Shoot Dry Weight (SDW), Root Dry Weight (RDW), Root to Shoot Ratio (RSR) and yield revealed significant difference among the accessions at p<0.05 level. Afeso and Sorcoll 163/07 showed better stress tolerance and the SG property. These accessions were also given maximum grain yield per hectare, while Sorcoll 146/07 has shown early maturity. Further, the correlation analysis revealed that girth diameter, number of tillers, chlorophyll content, leaf area, RWC, Pn, E, WUE, Fv/Fm, rate of leaf senescence, SDW, RDW and RSR had significant and positive relation with grain yield per hectare; and have been found to be effective screening tools for stress tolerance and SG property. Thus, they can be used in Sorghum breeding program.
PDF Abstract XML References Citation
Received: May 14, 2015;
Accepted: August 26, 2015;
Published: September 30, 2015
How to cite this article
Zelalem Getnet, Azamal Husen, Masresha Fetene and Gietahun Yemata, 2015. Growth, Water Status, Physiological, Biochemical and Yield Response of Stay Green Sorghum (Sorghum bicolor (L.) Moench) Varieties-A Field Trial Under Drought-Prone Area in Amhara Regional State, Ethiopia. Journal of Agronomy, 14: 188-202.
DOI: 10.3923/ja.2015.188.202
URL: https://scialert.net/abstract/?doi=ja.2015.188.202
DOI: 10.3923/ja.2015.188.202
URL: https://scialert.net/abstract/?doi=ja.2015.188.202
INTRODUCTION
Drought is one of the major constraints limiting crop production worldwide and feeding more people with less water is a major challenge at our planet. Several crop growth models predict that this issue will be more severe in the future. Drought impairs normal growth, disturbs water status and affects the yield; on the other hand a variety of morpho-physiological and biochemical alteration at cellular and whole organism level, making it a more complex phenomenon (Bota et al., 2004; Anjum et al., 2008; Husen, 2010; Aref et al., 2013; Husen et al., 2014). Effects of drought depend on the intensity and duration of stress, developmental stages and genotypic variation of the plants (Prasad et al., 2008; Anjum et al., 2008; Husen, 2010). Among the physiological characteristics, photosynthesis is severally affected by drought (Du et al., 2010; Husen et al., 2014), this is due to stomatal closure, which restricts CO2 diffusion to chloroplasts (Muller and Whitsitt, 1996). Non stomatal mechanism, in prolonged or severe drought, belongs to change in chlorophyll synthesis, functional and structural changes in chloroplast and also disturbance in accumulation and distribution of assimilation product (Medrano et al., 2002). Photosynthesis disturbance at molecular level are connected with the low electron transport through PSII and or/with structural injuries of PSII and light harvesting complexes (Van Rensburg and Kruger, 1993; Dubey, 1997). Measurement of maximum quantum yield of PSII efficiency (Fv/Fm) is a very useful method for the detection of changes in photosynthetic apparatus under stress condition (Ashraf et al., 2004; Husen, 2010; Kalaji et al., 2011; Husen et al., 2014) and also used to determine the seedling-stock quality (Husen, 2009; 2013). Reports are also available on drought-induced accumulation of osmolyte compounds such as inorganic ions, sugars and amino acids within the cell is often associated with a possible mechanism to tolerate the harmful effect of water stress (Elsheery and Cao, 2008; Xiao et al., 2008; Husen, 2010). Osmolytes accumulations in plant cells possibly contribute, via lowering the cell osmotic potential, to maintain several physiological processes under drought stress (Bajji et al., 2001) or vital in adaptive processes, i.e., osmotic adjustments (Yin et al., 2005; Xiao et al., 2008; Husen, 2010).
Sorghum is a C4 plant, evolved in Africa after splitting with rice 50-70 million years ago (Wolfe et al., 1989) and is a valuable global crop grown for food, feed, fibre and fuel (Paterson et al., 2009a, b). Sorghum is considered as a crop for future due to the changing global climatic trends and increase in use of marginal lands for agriculture (Paterson et al., 2008; CSA., 2011). It has been predicted that the global population will increase from about 7-9 billion by 2050 and most of the increase will occur in Sub-Saharan Africa, where population growth is the highest, in comparison to other part of the world (Haub, 2013) and gives the risk of food insecurity in this region (UNDP., 2012). Sorghum is widely grown in the high lands, low lands and semi-arid regions of Ethiopia, especially in moisture stressed parts where other crops least survive (Tesso et al., 2008). Sorghum production in Ethiopia has been declined due to population growth, land degradation, use of traditional farm implements and global climate change (Adugna, 2007). Drought is the major cause for underproduction of the crop (Kebede, 1991; Adugna, 2007). Drought response in Sorghum has been classified into two distinct stages, i.e., pre-flowering and post-flowering (Tuinstra et al., 1996). Stay-Green (SG) is integrated drought adaptation trait in Sorghum (Borrell et al., 2014). Severe drought condition during post-flowering stages led to loss of chlorophyll and increased senescence of leaves (Bray, 1993). Post-flowering drought resistance referred to as the SG trait has been reported in Sorghum (Xu et al., 2000; Borrell et al., 2000a) and in maize (Monneveux et al., 2006). Genotypes that have SG trait are reported to have reduced lodging and resistance to stalk rots, high level of stem carbohydrates, improved grain filling and grain yield under stress condition (Borrell et al., 2000a). The SG quantitative trail loci reduces canopy size at flowering stage by modifying tillering, leaf number and leaf size and smaller canopy size at flowering stage reduces pre-anthesis water uses, which under post flowering water stress increases water availability during grain filling and thus grain yield has been recorded (Borrell et al., 2014). Drought tolerant genotype proteome analysis indicated that the combined activities of several protein groups may enable the plants to tolerate drought stress and efficiently recover after removing the stress conditions (Jedmowski et al., 2014). Sorghum varieties have exhibited drought-tolerance in the field, did not necessarily exhibited photosynthetic adaptation to the various components of drought stress under controlled laboratory condition at a young developmental stage (Jagtap et al., 1998). Further, Sorghum under water deficit stress in sand culture also reduced dry weight, photosynthetic rate, transpiration rate, stomatal conductance and leaf water status (Liu et al., 2014).
In Ethiopia, many efforts have been made to address the drought problem in Sorghum production. Breeding programme has released a number of varieties from lowland areas, which gives a reasonable response and yield in drought-prone areas. Currently, Sorghum breeding in Ethiopia is fully engaged in different research activities in Sorghum drought tolerance. So far, two sources of SG, B-35 and E-36-1 were identified from the Ethiopian gene pools by the International Crops Research Institute for the Semi-Arid Tropics and now in use in different part of the world to generate drought resistance Sorghum varieties (Kebede, 1991). Sorghum germplasm originated in Ethiopia is also noted worldwide as a source for useful genes such as cold tolerance, good grain quality and disease/or insect resistance (Kebede, 1991). Sorghum accessions genetic diversity and primary screening of post-flowering drought tolerance has been investigated using few morphological and agronomic criteria (Mengiste, 2008; Yalew, 2010). The presence of variation among seven SG Sorghum varieties, under semi controlled conditions in glasshouse using morphological and physiological criteria has been examined and three better SG accessions were screened (Yalew, 2010). However, a detailed investigation related with morphological, physiological, biochemical and yield parameters under field condition on Sorghum accessions are not available. Therefore, the current experiment was designed with three Sorghum collections (Scroll) namely, Sorcoll-141/07, Sorcoll-146/07 and Sorcoll-163/07 in addition, a local accession (Afeso) was also used as a check.
MATERIALS AND METHODS
Study area and planting material: The experiment was conducted during 2013 cropping season at Shewa robit, North Shewa Zone of the Amhara Regional State, Ethiopia. The farm is situated at 10°00' N latitude and 39°53' E, longitude with an altitude of 1280 mean above sea level. During experimentation at the study area meteorological data was recorded and the rainfall received during cropping period ranged from 150.50 mm (July) to 0.00 mm (January), while the maximum and minimum temperature were 34.20°C (May) and 9.62°C (December), respectively. Shewa robit was chosen because this area is one of the most drought-prone areas and also represents the major Sorghum growing areas in the region. Three Stay-Green (SG) Sorghum accessions were selected. These accessions were Sorcoll-141/07, Sorcoll-146/07 and Sorcoll-163/07. A local accession (Afeso) was used as a check, since, it is a known source of Sorghum genotype trait and being used by local farmers.
Soil and fertilizer: Crops previously grown on the experimental area were maize (Zea may), Sorghum (Sorghum bicolor), Timbaho (Tobacco), teff (Eragrostis tef) and Masho (Vigna radiata). Some physical and chemical properties of the soil were analysed by collecting composite samples from each of the three plots at three depths i.e., 0-15, 15-30 and 30-45 cm. The soil pH was determined according to Juo (1978) and its electric conductivity was measured by using calibrated conductivity meter while, the texture class of the soil was determined by soil textural triangle, following the method of Juo (1978). The pH and electric conductivity of the composite soil sample was 7.5 and 979 μS cm–1, respectively, while the texture class of the mixed soil was silt clay loam. Urea [CO (NH2)2] and diammonium phosphate (DAP) were added to supplement the nutrient requirement of the plant.
Experimental design, plot management and sowing: The field experiment was laid out in Randomized Complete Block Design (RCBD) with three replications (three farmlands). Each farmlands covered an area of 484 m2 (22×22 m) and each was divided in to four equal parts to sow four different Sorghum varieties. There were 2 m gap in between the accessions. Inter-row distance was 0.75 m, which helps to create conducive environment for the proper management of the plots, while intra-row distance was 0.20 m maintained to avoid competition. Further, all land was prepared according to the local practice. It was ploughed three times using oxen before planting and the last ploughing was used for sowing. Sowing was done on July 8, 2013 and recommended rate of fertilizer was applied on the same day with the rate of 100/50 kg ha–1 urea and DAP. Tinning was done 15 days after sowing. Weeding and all other recommended crop protection activities were done manually, similar to the farmers practice recommended for Sorghum cultivation.
Plant growth measurement: Plant height, girth diameter, number of tillers, number of green leaves, number of senesced leaves and leaf area were determined on five randomly selected plants at 14, 30 and 42 DAF. Number of green leaves (More than 50% green portion) was counted. Number of senesced leaves was also counted when more than 50% of its area had senesced at the same date as number of green leaves was counted. Leaf area was measured by adopting Stickler’s linear measurement method (Stickler and Pauli, 1961).
Chlorophyll content index: Leaf chlorophyll content index was measured using Chlorophyll Content Meter (Opti-Sciences, Inc. USA) at 30 DAF. The readings were taken between 10:00 and 11:30 AM. A mean of 30 readings per plot was taken from five tagged plants. The readings were taken at the base of the leaf lamina, middle and towards the 1/3 of the tip of both the 2nd and flag leaves.
Flowering and physiological maturity: Anthesis (flowering) was noted when 50% population of S. bicolour accessions in each plot were flowered. Similarly, physiological maturity was determined by the appearance of black spot on the hilum of seed.
Relative Water Content (RWC): For RWC, the second or third fully expanded leaf from the top was brought from the field in polyethylene bags and stored in an ice box. Immediately, twenty leaf discs were weighted on an electronic balance (Citizen Scale, CY510, Poland) and Fresh Weight (FW) was determined. The weighted leaf discs were floated over night in a petridish containing distilled water and subsequently blotted gently and weighted again for Turgid Weight (TW). After taking turgid weight, the leaves were oven dried at 80°C for 48 h and Dry Weight (DW) was recorded separately. The RWC was calculated using the following formula (Weatherley, 1950):
Foliar gas exchange and water use efficiency: Net photosynthesis (A) and transpiration rate (E) were measured using Lc pro+photometer (ADC Bio Scientific Limited, UK). The readings were recorded forenoon between 10:00 and 11:30 am. A mean of 30 readings per plot were taken from five tagged plants at the base of the leaf lamina, middle and towards the 1/3 of the tip of both the second and flag leaves. Gas exchange measurements were performed on fully expanded and sun-exposed leaves throughout the natural photoperiod. Water Use Efficiency (WUE) was also determined by taking the ratio of assimilation rate to water lost due to transpiration.
Chlorophyll fluorescence (Fv/Fm): The Fv/Fm of leaves was recorded in the forenoon (10:00 and 11:30 an) by using Efficiency Analyzer (Hansatech Instruments Ltd., England) from the youngest and fully expanded leaves (five randomly selected plants) at 30 DAF from all accessions. Leaves were covered with clips and kept in dark for 30 min before measurements. The transients were induced by red light of 3000 μmol m‾2 sec–1 provided by an array of six light emitting diodes, which focused on the sample surface to give homogenous illumination over exposed area of sample surface and maximal quantum yield. After 30 min the maximum quantum yield of PSII efficiency (Fv/Fm) was measured (Genty et al., 1989).
Photosynthetic pigments: The leaf sample was taken from third/or fully expanded leaf from the top of five tagged plants and transported to the laboratory and preserved in polyethylene bag and kept in icebox. Total chlorophyll, chlorophyll ‘a’ and chlorophyll ‘b’ content of Sorghum leaf were estimated according to Witham et al. (1971) at 30 DAF.
Proline and total soluble sugar: Leaf proline content was measured according to the method of Bates et al. (1973); while, total soluble sugars was estimated by phenol sulphuric acid method (DuBois et al., 1956) at 30 DAF.
 | |
| Fig. 1(a-d): | Panicles of (a) Afeso, (b) Sorcoll 141, (c) Sorcoll 146 and (d) Sorcoll 163 captured on the study area (Shewa robit, Amhara Regional State, Ethiopia) at harvesting time |
Nitrogen and protein: Leaf nitrogen content was estimated by modified micro Kjeldhal method (Jackson, 1967) at 30 DAF with three steps (digestion, distillation and titration).
Biomass: Each accession was harvested and separated into shoot and root parts at 153 days. These parts were dried at 80°C in an oven until constant dry weight was obtained. Consequently, the dry weights were determined using a balance (OHAUS 1650-W0). Root to shoot ratio was determined from dry shoot and root biomass.
Grain weight and grain number per panicle: Dried panicles with mature seeds were taken from five previously tagged plants. Subsequently the panicles were allowed to air dry at room temperature (25oC) to remove excess moisture. Once the panicles were dried the seeds were threshed mechanically (using hand). Grain weight per plant was determined weighing all the seeds per panicle using balance (Citizen Scale, CY510, Poland). To determine the grain number per panicle, 1000 grains were counted from each of previously tagged five plants. The weight of 1000 grains was recorded separately in grams and the grain number per panicle was calculated from the grain weight per panicle and the corresponding 1000 grain weight as follow:
Grain yield (kg ha–1): All the panicles (Fig. 1) of each accession in every plot were collected, sun dried, threshed, cleaned and weight of the grains was recorded and expressed as kilogram per plot and converted to kilogram per hectare.
Data presentation and statistical analysis: All the collected data were subjected to analysis of variance using SPSS Software (Version 20, SPSS Inc., Chicago USA) to see variations between Sorghum accessions. The accession means were separated using the Least Significant Differences (LSD) test at 5% level of significance. The correlations between the studied traits were analysed using Pearson’s test.
RESULTS
Plant growth: Data showing the effect of field trial under drought-prone area on the studied plant growth is presented in Fig. 2 and Table 1. The check accession namely, Afeso, followed by Sorcoll 141/07 exhibited significantly maximum plant height, while minimum was recorded for Sorcoll 146/07 at 14, 30 and 42 Days After Flowering (DAF) (Fig. 2a). The average data of three DAF showed maximum plant height with Afeso followed by Sorcoll 141/07, Sorcoll 146/07 and Sorcoll 163/07 (Table 1). Girth diameter increment was more with Sorcoll 163/07, however, it showed insignificant variation with Afeso and the lowest was recorded with Sorcoll 146/07 at 14, 30 and 42 DAF (Fig. 2b). The average data of three DAF exhibited maximum girth diameter with Sorcoll 163/07 but again it showed insignificant variation with Afeso and Sorcoll 141/07 and minimum was found with Sorcoll 146/07 (Table 1). Insignificant variation was found for number of tillers (data not shown) through passage of time. However, the average data of three DAF showed significantly more tillers formation with Sorcoll 163/07 followed by Afeso, Sorcoll 146/07 and Sorcoll 141/07 (Table 1). Further, number of green leaf formation and leaf area exhibited a reduction trend in the studied Sorghum accession at 14, 30 and 42 DAF (Fig. 2c and e). Sorcoll 141/07 and Sorcoll 146/07 showed more reduction, while Sorcoll 163/07 and Afeso were able to maintain almost same amount of leaf throughout the measurement interval. The average data of three DAF showed the lowest value in Sorcoll 146/07 while the highest was found with Afeso (Table 1). Number of leaf senescence was increased as the time passed from 14, 30 and 42 DAF, its increasing rate was less in Afeso in comparison to other Sorghum accessions (Fig. 2d). The average data of three DAF also exhibited less number of senescence with Afeso and more with Sorcoll 146/07 (Table 1).
| Table 1: | Morphological and agronomic parameters of Sorghum bicolour accessions |
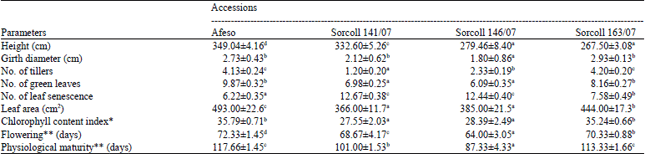 | |
| *as measured on 30 days after flowering using chlorophyll content meter; **Were noted when 50% of Sorghum bicolour accessions population in each plot flowered and the appearance of black spot on the hilum of seed was taken as the indication of physiological maturity, Data represent the Mean±SE of the three replicates, means followed by the same letter in a row are not significantly different at p<0.05 level | |
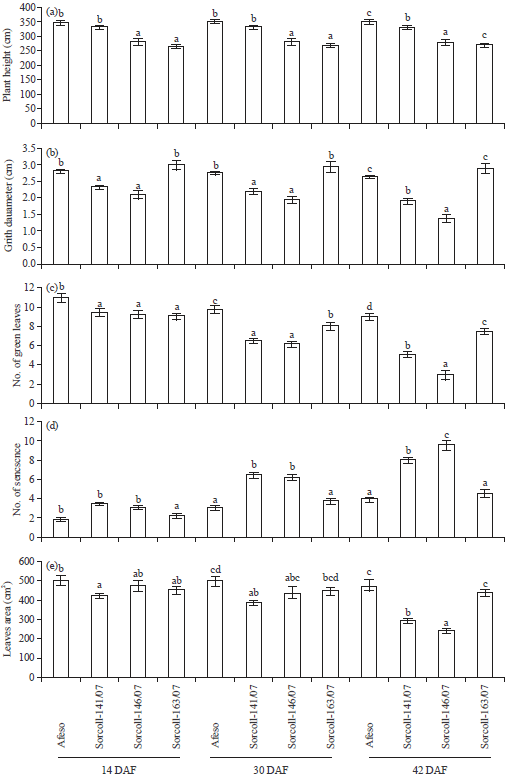 | |
| Fig. 2(a-e): | Changes in (a) Plant height, (b) Girth diameter, (c) No. of green leaves, (d) No. of leaf senescence and (e) Leaf area of Sorghum bicolour accessions at 14, 30 and 42 days after flowering. Bars represent Mean±SE of the three replicates and designated with the same letter are not significantly different between accessions at p<0.05 level |
Leaf chlorophyll content index was measured only once at 30 DAF, Sorcoll 163/07 exhibited the highest value and which was found statistically same as Afeso, while Sorcoll 146/07 showed the lowest value and insignificantly differs with Sorcoll 141/07 (Table 1). Further, the days for 50% flowering and physiological maturity of the studied Sorghum accessions showed that the Afeso was the best followed by Sorcoll 163/07, Sorcoll 141/07 and Sorcoll 146/07 (Table 1).
Physiological traits: Relative Water Content (RWC), net photosynthetic rate (Pn), transpiration rate (E), Water Use Efficiency (WUE) and maximum quantum yield of PSII efficiency (Fv/Fm) was measured at 30 Days After Flowering (DAF) and presented in Fig. 3. The RWC was found more in Afeso which exhibit insignificant variation with Sorcoll 163/07, followed by Sorcoll 141/07 and Sorcoll 146/07 (Fig. 3a). The highest Pn and WUE were recorded in Sorcoll 163/07 accession (Fig. 3b and d). Statistically, Pn response was similar for Sorcoll 163/07 and Afeso. At 42 DAF, value for E was more for Afeso and less for Sorcoll 163/07 (Fig. 3c). The Fv/Fm value was found higher among the Afeso followed by Sorcoll 163/07, Sorcoll 141/07 and Sorcoll 146/07 accessions. However, the higher value for Fv/Fm responded statistically same for Afeso and Sorcoll 163/07. Likewise, Sorcoll 141/07 and Sorcoll 146/07 accessions showed insignificant variation (Fig. 3e).
Biochemical traits: Photosynthetic pigments, proline, nitrogen, protein and soluble sugar contents in leaves were estimated at 30 Days After Flowering (DAF) and given in Table 2. Chlorophyll a, b and total chlorophyll content found more in Sorcoll 163/07 and Afeso and both were shown insignificant variation, while less value was observed in Sorcoll 146/07 accession (Table 2). Leaf proline was observed maximum in Sorcoll 163/07 followed by Afeso, Sorcoll 141/07 and Sorcoll 146/07 accessions (Table 2). Total soluble sugar was highest in Afeso but it shows insignificant variation with Sorcoll 163/07, while Sorcoll 141/07 exhibited the lowest value (Table 2). Leaf nitrogen and protein contents was found more in Sorcoll 163/07 followed by Afeso, Sorcoll 141/07 and Sorcoll 146/07 accessions (Table 2).
Biomass: Shoot dry weight at the harvesting time (153 days) was recorded more in Afeso followed by Sorcoll 141/07, Sorcoll 163/07 and Sorcoll 146/07 accession (Fig. 4a). Root dry weight was also found more with Afeso followed by Sorcoll 163/07, Sorcoll 141/07 and Sorcoll 146/07 accessions (Fig. 4b).
| Table 2: | Biochemical activities of Sorghum bicolour accessions as measured on 30 days of flowering |
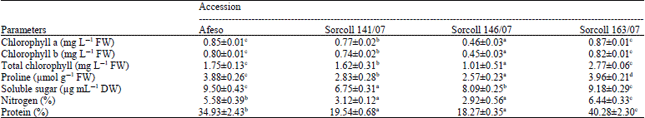 | |
| Data represent the Mean±SE of the three replicates. Means followed by the same letter in a row are not significantly different at p<0.05 level | |
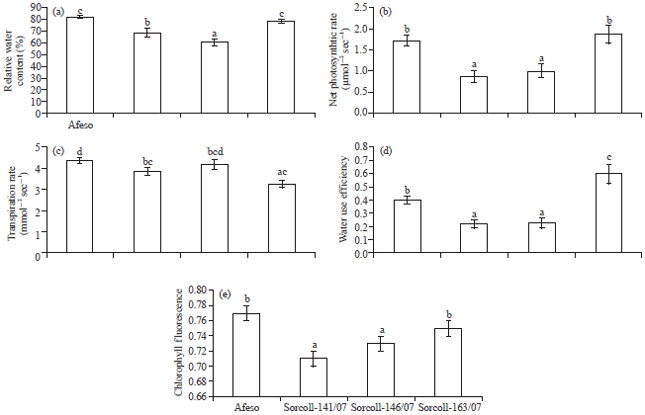 | |
| Fig. 3(a-e): | Changes in (a) Relative water content, (b) Net photosynthetic rate, (c) Transpiration rate, (d) Water use efficiency and (e) Chlorophyll fluorescence (Fv/Fm) of Sorghum bicolour accessions at 42 days after flowering. Bars represent Mean±SE of the three replicates and designated with the same letter are not significantly different between accessions at p<0.05 level |
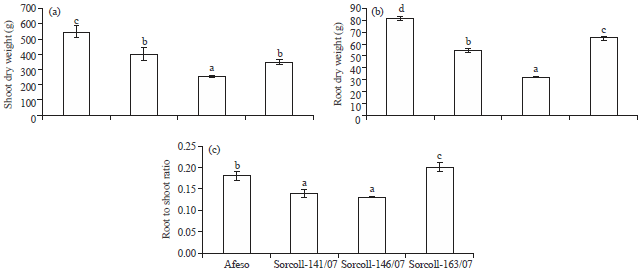 | |
| Fig. 4(a-c): | Changes in biomass (a) Shoot dry weight, (b) Root dry weight and (c) Root to shoot ratio at the harvesting time (153 days). Bars represent Mean±SE of the three replicates and designated with the same letter are not significantly different between accessions at p<0.05 level |
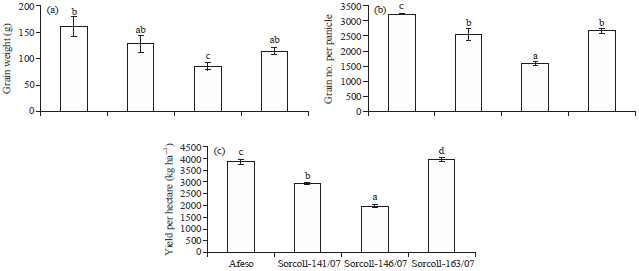 | |
| Fig. 5(a-c): | Changes in yield response (a) Grain weight, (b) Grain number per panicle and (c) Yield per hectare. Bars represent Mean±SE of the three replicates and designated with the same letter are not significantly different between accessions at p<0.05 level |
However, root to shoot ratio was observed more in Sorcoll 163/07 followed by Afeso, Sorcoll 141/07 and Sorcoll 146/07 accessions (Fig. 4c).
Yield: A comparative observation for grain weight, grain number per panicle and yield per hectare is presented in Fig. 5. Grain weight and grain number per panicle was found maximum by Afeso and minimum by Sorcoll 146/07 (Fig. 5a and b). However, yield per hectare was recorded maximum by Sorcoll 163/07 which showed very less variation with Afeso and minimum was observed by Sorcoll 146/07 accession (Fig. 5c).
Correlation studies: Correlation was performed for various parameters and presented in Table 3. Plant height was positively correlated with number of green leaves, chlorophyll content, shoot dry weight, root dry weight and root to shoot ratio under this field trial. Leaf area showed a positive relation with girth diameter, Fv/Fm, root dry weight, root to shoot ratio and with all studied biochemical traits such as nitrogen, protein, proline and soluble sugar contents. Chlorophyll content index had also significant positive correlation with girth diameter, leaf area, A, WUE, RWC, protein, nitrogen, shoot dry weight, root to shoot ratio and yield per hectare. However, yield per hectare had a negative correlation with number of leaf senescence. Further, 50% flowering had significant positive relation with physiological maturity, RWC and yield per hectare.
| Table 3: | Pearson’s correlation study among the different morphological, physiological and biochemical traits of Sorghum bicolour accessions |
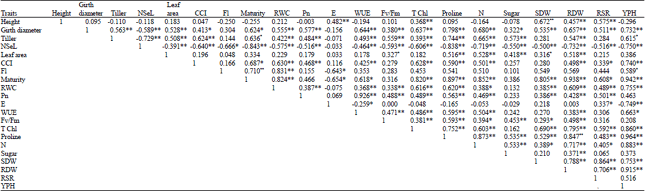 | |
| NSeL: Number of leaf senescence, CCI: Chlorophyll content index, Fl: Flowering, RWC: Relative water content, Pn: Net photosynthetic rate, E: Transpiration rate, WUE: Water use efficiency, Fv/Fm: Chlorophyll fluorescence, T Chl: Total chlorophyll, N: Leaf nitrogen, SDW: Shoot dry weight, RDW: Root dry weight, RSR: Root to shoot ratio and YPH: Yield per hectare, ** and *: Significant at p = 0.01 and p = 0.05 levels (N = 12-60) | |
Physiological maturity also had a significant positive correlation with girth diameter, chlorophyll content index, RWC, WUE, chlorophyll, protein, proline and nitrogen contents, shoot dry weight, root dry weight and root to shoot ratio. However, both 50% flowering and maturity had a negative correlation with plant height, number of leaf senescence and E. Further, A had a significant positive correlation with many traits like girth diameter, number of green leaves, RWC, WUE, Fv/Fm, chlorophyll content and all other estimated biochemical traits, while A was negatively related with number of leaf senescence. Chlorophyll contents had positive correlation with chlorophyll content index, WUE, RWC, leaf nitrogen and proline contents. Proline content had significant positive correlation with leaf area, Fv/Fm, chlorophyll content and grain yield. Correlation study also indicated a positive association of total sugars with total chlorophyll content of the tested accessions. Further, positive correlation of leaf nitrogen with leaf area, RWC, root to shoot ratio and grain yield was observed. Grain yield was also had a positive correlation with girth diameter, number of green leaves, leaf area, chlorophyll content index, chlorophyll content, proline accumulation, shoot dry weight, root dry weight and root to shoot ratio while, grain yield per panicle had negative correlation with number of leaf senescence.
DISCUSSION
Growth of crop plants and yield are under increasing pressure by global anthropogenic changes, including rising population, diversion of cereals to biofuels, increased protein demands and extreme unfavourable climatic condition. The extreme unfavourable climatic condition will also increase the intensity and frequency such as higher temperature, more intense precipitation, increase risk and duration of drought. In our study, Sorghum varieties exhibited different tolerance response under field condition. It was found that plant height, number of green leaves, leaf area, chlorophyll content index, flowering and physiological maturity were more with the local accession (Afeso) in comparison to the tested accessions namely, Sorcoll-141/07, Sorcoll-146/07 and Sorcoll-163/07. In addition, plant biomass was also found higher in Afeso. However, root-shoot and yield per hectare was found more with Sorcoll-163/07. Afeso is a local accession and growing in drought-prone area, as a consequence, perhaps have developed various mechanisms to enhance their tolerance, including well-developed root system and certain leaf morpho-physiological characteristic features (Larcher, 1995; Kozlowski and Pallardy, 2002; Husen, 2010; Husen et al., 2014). Tillering is an important agronomical trait and it has a major impact on leaf area development of S. bicolor in current study and others (Lafarge et al., 2002). Similarly, Lafarge et al. (2002) report the number of fertile tillers per plant can vary from zero to around four depending on growing conditions and variety. The present study show significant difference in the number of tillers among accessions and it can be suggested that the higher potential of Sorcoll 163/07 and the check (Afeso) for tiller formation perhaps due to the genetic background of the accessions (Dolciotti et al., 1998; Yalew, 2010). Afeso and Sorcoll 163/07 showed lower rate of leaf senescence. Sorcoll 163/07 showed Stay Green (SG) phenotype, characterized by an extended green leaf period. This can arise from delays in the beginning of senescence and/or in senescence progress, as previously reported in rice varieties (Rampino et al., 2006). Afeso and Sorcoll 163/07 not only had higher chlorophyll content index but also sustained green leaves longer throughout post-flowering condition than the remaining accessions. Sorghum plants with the SG trait resist premature senescence when subjected to stress factors during grain-filling stage (Borrell et al., 2000a). In contrast, Sorcoll 146/07 is considered as senescent accessions than the remaining three accessions because this accession revealed the highest rate of senescence. The observed remarkable variation might be due to genetic variation for leaf senescence and differences in series of biochemical and physiological changes occurring during senescence (Lim et al., 2007). This suggests that the leaf senescence could provide complementary information about stress tolerance and for screening genotypes with SG for breeding purpose. Sorcoll 163/07 had taken more number of days to 50% flowering and maturity. The number of days taken for grain filling was also higher. Sorghum accessions which have delayed maturity and higher number of days to grain filling under drought stress are better SG genotypes has been also reported (Rosenow et al., 1983; Craufurd and Peacock, 1993; Prasad et al., 2008). Accordingly in this regard, Sorcoll 163/07 and the Afeso were found better SG accessions. However, the early matured accession Sorcoll 146/07 had significantly lower number of days to 50% flowering, maturity and the number of days for grain filling under drought-prone area. Thus, such traits can be considered as ways of drought escape mechanism on several crop plants (Turner and Kramer, 1980). Another remarkable aspect is that the behaviour of different drought-tolerant Sorghum accessions was not uniform in terms of pigment concentration, proline, soluble sugar, nitrogen and protein content. Soluble sugar was found more in Afeso while remaining all studied biochemical activities were found more in the Sorcoll-163/07. This could well be a case of hormesis, under low level of stress (Aref et al., 2015). The physiological, growth and developmental mechanisms which allow a plant species to tolerate prolonged periods of drought can involve numerous attributes. One means of increasing drought tolerant is by the accumulation of osmotically active solutes, so that turgor and turgor-dependent processes may be maintained during episode of dry-down. The accumulation of protective solutes, such as sugars (Elsheery and Cao, 2008) and proline (Husen, 2010) is a unique response to environmental stress, specifically to drought. Correlation between sugar accumulation and osmotic stress tolerance has been widely reported, including transgenic experiments (Chaves et al., 2003; Bartels and Sunkar, 2005). Sugars have different functions in plants from energy storage to signaling and plants utilize several sugar-based strategies to adapt to environmental stresses (Anderson and Kohorn, 2001; Chaves et al., 2003). Proline plays a prominent role as an osmoticum and, because of its zwitterionic in plant cells appears to be related to mechanism associated with tolerance to desiccation (Rontein et al., 2002); also as a sink of energy or reducing power (Verbruggen et al., 1996), as a scavenger of ROS and their derivatives (Hong et al., 2000; Bashir et al., 2007) and as a source for carbon and nitrogen (Peng et al., 1996) and thereby helping the plants to tolerate stress effects (Manivannan et al., 2007). All these may explain the increase in sugar and proline content with the Sorcoll-163/07 under drought stress in this study. Further, decrease in chlorophyll content with the Sorcoll-146/07 and Sorcoll-141/07, perhaps due to reduction in the lamellar content of the light harvesting Chl a/b protein that accounts for the elevated Chl a/b ratio (2.8:4.5) and/or due to drought stress-induced decrease in the availability of mineral nutrients, and/or to the damages to chloroplasts by ROS and or either slow synthesis or fast breakdown of chlorophyll pigments (Khanna-Chopra et al., 1980; Ashraf, 2003).
Relative Water Content (RWC) has been used for evaluation of SG expression to measure post-flowering drought tolerance (Rosenow and Clark, 1981). In this study, Afeso maintained the highest RWC than other accessions; perhaps the better SG accessions keep the stalk transpiration system functioning under severe drought conditions (Xu et al., 2008). Sorcoll 163/07 revealed a better SG property owing to the possession of higher RWC, next to Afeso, during post-flowering terminal drought (Xu et al., 2008). This indicates that RWC is a potential tool for screening genotypes under post flowering drought stress. Drought stress-induced decrease in RWC has been also reported in many previous studies (Elsheery and Cao, 2008; Xiao et al., 2008; Husen et al., 2014). In the present study, net photosynthetic rate (Pn), transpiration rate (E), Water Use Efficiency (WUE) and maximum quantum yield of PSII efficiency (Fv/Fm) significantly varied among the tested Sorghum cultivars under drought-prone area. Sorcoll 163/07 was found photosynthetically more active, followed by the local Afeso, while Sorcoll 141/07 had the lowest value. Higher Pn under limited water supply conditions is one of the factors for realizing higher grain yield as found in the present investigation because, it is expected to provide the raw material and the energy required for growth and development. This phenomenon also reveals that these cultivars employ the physiologically drought-avoidance strategy. Better yield bearing accession Sorcoll 163/07 had the low transpiration rate than all other accessions. Stomatal closure is a probable factor responsible for the reduction in transpiration under drought condition because, stomatal closure is a well-known and most efficient way of reducing transpirational water loss (Cornic, 2000). Moreover, the Afeso had higher E. It has been reported that crops with SG characteristics have enhanced transpiration efficiency, which enables the plant to set a higher yield potential during anthesis that ultimately leads to higher grain yield (Borrell et al., 2001). Sorcoll 163/07 and the local accession Afeso had higher WUE. These two accessions showed comparatively higher WUE, suggesting improved drought tolerance achieved by optimizing CO2 uptake per unit water lost through transpiration. Possibly, this is done through maintaining better RWC and opening of stomata partially, this feature allowing a relatively higher rate of assimilation and transpiration (Cornic, 2000). Since, stomatal regulation controls the exchange of water and carbon between the leaf and the atmosphere it in turn would affect WUE (Condon et al., 2002). Furthermore, the lower WUE of Sorcoll 141/07 and Sorcoll 146/07 were perhaps due to extensive stress-induced leaf senescence as found in this investigation. Therefore, WUE indicates the tissue water relation of Sorghum accessions and suggests difference in adaptation strategies among the accessions to stress. The Fv/Fm is a valuable tool for quantifying the effect of abiotic stress on photosynthesis (Tezara et al., 2005; Husen, 2010; Husen et al., 2014), particularly for estimating the degree of photo-inhibition (Tezara et al., 2005). Sorcoll 163/07 and Afeso maintained higher Fv/Fm ratio in comparison to other accessions. The ability of these cultivars in maintaining high Fv/Fm ration under terminal drought condition might indicate high efficiency of radiation use for photochemistry and carbon assimilation (Massacci and Jones, 1990; Husen, 2010).
Yield is a complex polygenic trait, which is highly influenced by the environment and season. In the present study, the accessions varied significantly on yield per panicle. Higher grain yield per panicle in SG types have been directly attributed to maintaining photosynthetic capability during the grain filling period (Borrell et al., 2000b). Sorcoll 163/07 and Afeso, which has resulted in high WUE could have favoured high assimilation and finally high grain yield under the studied drought-prone area. Similarly, the minimum grain yield per panicle of Sorcoll 146/07 might be attributed to the lowest assimilation rate (Sinclair and Jamieson, 2008). Similar results were obtained by Borrell et al. (2000b) and Yalew (2010). Moreover, the reduction of grain yield per panicle might be as a result of fewer seeds per unit area and smaller size of the panicle as it has been reported in soybean (Praba et al., 2009). Perhaps, drought induced leaf senescence might affect CO2 uptake and that in turn could reduce grain yield. Similarly, studies on Sorghum by Yalew (2010) and on maize varieties by Monneveux et al. (2006) have indicated that drought greatly reduces grain yield, which was dependent on the level of senescence due to the stress during grain filling. Thus, the result helps to infer the existence of genotypic difference in grain yield per panicle. Since, the variation in grain yield per panicle has found to be a function of terminal drought (Khanna-Chopra et al., 1994).
Correlation analysis revealed that plant height had a positive correlation with most of the parameters while it showed negative relationship with grain yield. Similar observation was reported by Hiramath and Parvatikar (1985) and Pawar (2007), while Bakheit (1990) and Blum et al. (1989) found a positive correlation between plant height, dry matter production and grain yield. This may be because of increased translocation of stored photosynthates from the stem reserves when the current photosynthesis ceases due to environmental stress factors, particularly during grain filling and grain development period (Erick and Musick, 1979). Leaf area was found to have significant correlation with few morpho-physiological and biochemical traits and these results are consistent with previous reports (Rosenow et al., 1983; Borrell et al., 2000a; Pawar, 2007). Chlorophyll content index had a positive relation with most of the parameters; except while grain yield have no correlation with number of leaf senescence. These results are in conformity with those obtained by Kadam et al. (2002) in Sorghum and Sairam (1994) in wheat. RWC and girth diameter, leaf area, 50% flowering, physiological maturity, WUE, grain yield, chlorophyll, proline, nitrogen and protein contents had shown positive correlation while it was insignificant for E and number of leaf senescence. Similar result has been reported by many workers (McBee and Miller, 1982; Salunke et al., 2003; Prasad et al., 2008). Net photosynthetic rate had positive correlation with the studied parameters except number of leaf senescence; leaf pigments were also positively correlated with chlorophyll content index, WUE, RWC, leaf nitrogen and proline contents. Proline content was correlated with leaf area, Fv/Fm, chlorophyll content and grain yield. Variations in photosynthesis are associated with leaf protein content, nitrogen and chlorophyll content per unit area (Swain, 1986). It also indicates that the accession with a higher proline accumulation under a water limited environment had a better SG trait by preventing the water in the plant from loss. Proline accumulation increased in response to water stress is a common phenomenon (Husen, 2010). In addition, Pawar (2007) also reported correlation of proline accumulation with grain yield in water limited environment. Total sugars with total chlorophyll content could be another reason for higher yield in these genotypes. Premachandra et al. (1995) also reported that total soluble sugars are the major solutes contributing to osmotic potential in Sorghum. Leaf nitrogen was also found for higher yield response. It also had a positive correlation with chlorophyll contents. This was expected because nitrogen is one of the components of chlorophyll molecule. The higher nitrogen content in the leaf naturally increases chlorophyll content, which in turn increases the photosynthetic rate (Borrell and Hammer, 2000).
CONCLUSION
In conclusion, the results of the present study showed that the studied Sorghum accessions demonstrated variation in response to the measured parameters under post-flowering in the drought-prone area. Thus, the variations might elucidate the existence of genotypic difference among the accessions. Sorcoll 163/07 and Afeso appeared to be more resistant to severe post-flowering drought stress than Sorcoll 141/07 and Sorcoll 146/07; this can be ascribed to maintaining the higher green leaf area, RWC, chlorophyll content index, assimilation rate, Fv/Fm, WUE, delayed leaf senescence, root dry weight and root to shoot ratio. They also had more accumulation of biochemical metabolites like (leaf pigments, soluble sugar, free proline, leaf nitrogen and protein contents) and higher grain yield. On the basis of the various morphological, physiological and biochemical traits measured and correlation studies, Sorcoll 163/07 and Afeso was found to show better SG property and higher grain yield under post flowering drought stress, next to Afeso, while Sorcoll 146/07 was an early matured genotype. These two SG accessions on the genetic aspect for breeding purpose can be explored further. Moreover, the better performances of the local accession, namely Afeso have indicated that still additional better SG accessions are available in different part of Ethiopia including the study site.
ACKNOWLEDGMENTS
The research was supported by University of Gondar, Gondar, Ethiopia under Teaching and Learning Program (UoG/Budget code: 6417). Authors are also thankful to Addis Ababa University and the Bio-innovate Project for partially providing materials and running costs during this investigation. Thanks to the Ethiopian Meteorological Agency for providing climatic data. Zelalem Getnet is thankful to Mr. Awol Assefa for technical assistance, Addis Ababa University, Addis Ababa, Ethiopia.
REFERENCES
- Aref, I.M., P.R. Khan, A.A. Al Sahli, A. Husen, M.K.A. Ansari and M. Iqbal, 2015. Response of Datura innoxia Linn. to gamma rays and its impact on plant growth and productivity. Proc. Natl. Acad. Sci. India Sect. B: Biol. Sci., (In Press).
CrossRefDirect Link - Genty, B., J.M. Briantais and N.R. Baker, 1989. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimica et Biophysica Acta (BBA)-General Subjects, 990: 87-92.
CrossRefDirect Link - Kalaji, H.M., Govindjee, K. Bosa, J. Koscielniak and K. Zuk-Golaszewska, 2011. Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environ. Exp. Bot., 73: 64-72.
CrossRefDirect Link - Weatherley, P.E., 1950. Studies in the water relations of the cotton plant. I. The field measurement of water deficits in leaves. New Phytol., 49: 81-97.
CrossRefDirect Link - Anderson, C.M. and B.D. Kohorn, 2001. Inactivation of Arabidopsis SIP1 leads to reduced levels of sugars and drought tolerance. J. Plant Physiol., 158: 1215-1219.
CrossRefDirect Link - Anjum, N.A., S. Umar, M. Iqbal and N.A. Khan, 2008. Growth characteristics and antioxidant metabolism of moongbean genotypes differing in photosynthetic capacity subjected to water deficit stress. J. Plant Interactions, 3: 127-136.
CrossRefDirect Link - Aref, I.M., A.I. Ahmed, P.R. Khan, H.A. El-Atta and M. Iqbal, 2013. Drought-induced adaptive changes in the seedling anatomy of Acacia ehrenbergiana and Acacia tortilis subsp. raddiana. Trees, 27: 959-971.
CrossRefDirect Link - Adugna, A., 2007. The role of introduced sorghum and millets in Ethiopian agriculture. SAT e-J., 3: 1-9.
Direct Link - Ashraf, M., M.Y. Ashraf, A. Khaliq and E.S. Rha, 2004. Growth and leaf gas exchange characteristics in Dalbergia sissoo Roxb. and D. latifolia Roxb. under water deficit. Photosynthetica, 42: 157-160.
CrossRefDirect Link - Ashraf, M., 2003. Relationships between leaf gas exchange characteristics and growth of differently adapted populations of Blue panicgrass (Panicum antidotale Retz.) under salinity or waterlogging. Plants Sci., 165: 69-75.
CrossRefDirect Link - Bajji, M., S. Lutts and J.M. Kient, 2001. Water deficit effects on solute contribution to osmotic adjustment as a function of leaf ageing in three durum wheat (Triticum durum Desf.) cultivars performing differently in arid conditions. Plant Sci., 160: 669-681.
Direct Link - Bakheit, B.R., 1990. Variability and correlations in grain sorghum genotypes (Sorghum bicolor [L.] Moench) under drought conditions at different stages of growth. J. Agron. Crop Sci., 165: 355-360.
CrossRefDirect Link - Bartels, D. and R. Sunkar, 2005. Drought and salt tolerance in plants. Crit. Rev. Plant Sci., 24: 23-58.
CrossRefDirect Link - Bashir, F., Mahmooduzzafar, T.O. Siddiqi and M. Iqbal, 2007. The antioxidative response system in Glycine max (L.) Merr. exposed to Deltamethrin, a synthetic pyrethroid insecticide. Environ. Pollut., 147: 94-100.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Blum, A., J. Mayer and G. Golan, 1989. Agronomic and physiological assessments of genotypic variation for drought resistance in sorghum. Crop Pasture Sci., 40: 49-61.
CrossRefDirect Link - Borrell, A.K., G.L. Hammer and E.V. Oosterom, 2001. Stay-green: A consequence of the balance between supply and demand for nitrogen during grain filling? Ann. Applied Biol., 138: 91-95.
CrossRefDirect Link - Borrell, A.K., G.L. Hammer and A.C.L. Douglas, 2000. Does maintaining green leaf area in sorghum improve yield under drought? I. Leaf growth and senescence. Crop Sci., 40: 1026-1037.
CrossRefDirect Link - Borrell, A.K., G.L. Hammer and R.G. Henzell, 2000. Does maintaining green leaf area in sorghum improve yield under drought? II. Dry matter production and yield. Crop Sci., 40: 1037-1048.
CrossRefDirect Link - Borrell, A.K., J.F. Mullet, B. George-Jaeggli, E.J. van Oosterom, G.L. Hammer, P.E. Klein and D.R. Jordan, 2014. Drought adaptation of stay-green sorghum is associated with canopy development, leaf anatomy, root growth and water uptake. J. Exp. Bot., 65: 6251-6263.
CrossRefDirect Link - Borrell, A.K. and G.L. Hammer, 2000. Nitrogen dynamics and the physiological basis of stay-green in sorghum. Crop Sci., 40: 1295-1307.
CrossRefDirect Link - Bota, J., H. Medrano and J. Flexas, 2004. Is photosynthesis limited by decreased Rubisco activity and RuBP content under progressive water stress? New Phytol., 162: 671-681.
CrossRefDirect Link - Bray, E.A., 1993. Molecular responses to water deficit. Plant Physiol., 103: 1035-1040.
PubMedDirect Link - Chaves, M.M., J.P. Maroco and J.S. Pereira, 2003. Understanding plant responses to drought-from genes to the whole plant. Funct. Plant Biol., 30: 239-264.
CrossRefDirect Link - Condon, A.G., R.A. Richards, G.J. Rebetzke and G.D. Farquhar, 2002. Improving intrinsic water-use efficiency and crop yield. Crop Sci., 42: 122-131.
PubMedDirect Link - Cornic, G., 2000. Drought stress inhibits photosynthesis by decreasing stomatal aperture-not by affecting ATP synthesis. Trends. Plant Sci., 5: 187-188.
CrossRefDirect Link - Craufurd, P.Q. and J.M. Peacock, 1993. Effect of heat and drought stress on sorghum (Sorghum bicolor). II. Grain yield. Exp. Agric., 29: 77-86.
CrossRefDirect Link - Dolciotti, I., S. Mambelli, S. Grandi and G. Venturi, 1998. Comparison of two sorghum genotypes for sugar and fiber production. Ind. Crops Prod., 7: 265-272.
CrossRefDirect Link - Du, N., W. Guo, X. Zhang and R. Wang, 2010. Morphological and physiological responses of Vitex negundo L. var. heterophylla (Franch.) Rehd. to drought stress. Acta Physiol. Plant., 32: 839-848.
CrossRefDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Elsheery, N.I. and K.F. Cao, 2008. Gas exchange, chlorophyll fluorescence and osmotic adjustment in two mango cultivars under drought stress. Acta Physiol. Plant., 30: 769-777.
CrossRefDirect Link - Erick, H.V. and J.T. Musick, 1979. Plant water stress effects on irrigated grain sorghum. I. Effects on yield. Crop, 19: 589-592.
CrossRefDirect Link - Hiramath, S.M. and S.R. Parvatikar, 1985. Growth and yield analysis in sorghum-identification of genotypes with low leaf area and high dry matter production. Sorghum Newslett., 28: 108-112.
Direct Link - Hong, Z., K. Lakkineni, Z. Zhang and D.P.S. Verma, 2000. Removal of feedback inhibition of Δ1-pyrroline-5-carboxylate synthetase results in increased proline accumulation and protection of plants from osmotic stress. Plant Physiol., 122: 1129-1136.
Direct Link - Husen, A., 2009. Growth, chlorophyll fluorescence and biochemical markers in clonal ramets of shisham (Dalbergia sissoo Roxb.) at nursery stage. New For., 38: 117-129.
CrossRefDirect Link - Husen, A., 2010. Growth characteristics, physiological and metabolic responses of teak (Tectona grandis Linn. f.) clones differing in rejuvenation capacity subjected to drought stress. Silvae Genet., 59: 124-136.
Direct Link - Husen, A., 2013. Growth characteristics, biomass and chlorophyll fluorescence variation of Garhwal Himalaya's fodder and fuel wood tree species at the nursery stage. Open J. Forestry, 3: 12-16.
CrossRefDirect Link - Husen, A., M. Iqbal and I.M. Aref, 2014. Growth, water status and leaf characteristics of Brassica carinata under drought and rehydration conditions. Braz. J. Bot., 37: 217-227.
CrossRefDirect Link - Jagtap, V., S. Bhargava, P. Sterb and J. Feierabend, 1998. Comparative effect of water, heat and light stresses on photosynthetic reactions in Sorghum bicolor (L.) moench. J. Exp. Bot., 49: 1715-1721.
CrossRefDirect Link - Jedmowski, C., A. Ashoub, T. Beckhaus, T. Berberich, M. Karas and W. Bruggemann, 2014. Comparative analysis of Sorghum bicolor proteome in response to drought stress and following recovery. Int. J. Proteomics, Vol. 2014.
CrossRefDirect Link - Kadam, G.N., S.R. Gadakh and V.R. Awari, 2002. Physiological analysis of rabi sorghum genotypes for shallow soil. J. Maharashtra Agric. Univ., 27: 274-276.
Direct Link - Khanna‐Chopra, R., G.S. Chaturverdi, P.K. Aggarwal and S.K. Sinha, 1980. Effect of potassium on growth and nitrate reductase during water stress and recovery in maize. Physiol. Plantarum, 49: 495-500.
CrossRefDirect Link - Khanna-Chopra, R., P.S.S. Rao, M. Maheswari, L. Xiaobing and K.S. Shivshankar, 1994. Effect of water deficit on accumulation of dry matter, carbon and nitrogen in the kernel of wheat genotypes differing in yield stability. Ann. Bot., 74: 503-511.
Direct Link - Kozlowski, T.T. and S.G. Pallardy, 2002. Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev., 68: 270-334.
CrossRefDirect Link - Lafarge, T.A., I.J. Broad and G.L. Hammer, 2002. Tillering in grain sorghum over a wide range of population densities: Identification of a common hierarchy for tiller emergence, leaf area development and fertility. Ann. Bot., 90: 87-98.
CrossRefDirect Link - Lim, P.O., H.J. Kim and H.G. Nam, 2007. Leaf senescence. Ann. Rev. Plant Biol., 58: 115-136.
CrossRefDirect Link - Liu, P., L. Yin, X. Deng, S. Wang, K. Tanaka and S. Zhang, 2014. Aquaporin-mediated increase in root hydraulic conductance is involved in silicon-induced improved root water uptake under osmotic stress in Sorghum bicolor L. J. Exp. Bot., 65: 4747-4756.
CrossRefDirect Link - Manivannan, P., C.A. Jaleel, B. Sankar, A. Kishorekumar, R. Somasundaram, G.M.A. Lakshmanan and R. Panneerselvam, 2007. Growth, biochemical modifications and proline metabolism in Helianthus annuus L. as induced by drought stress. Colloids Surf. B: Biointerfaces, 59: 141-149.
CrossRefDirect Link - Massacci, A. and H.G. Jones, 1990. Use of simultaneous analysis of gas-exchange and chlorophyll fluorescence quenching for analysing the effects of water stress on photosynthesis in apple leaves. Trees, 4: 1-8.
CrossRefDirect Link - McBee, G.G. and F.R. Miller, 1982. Carbohydrates in sorghum culms as influenced by cultivars, spacing and maturity over a diurnal period. Crop Sci., 22: 381-385.
CrossRefDirect Link - Medrano, H., J.M. Escalona, J. Bota, J. Gulias and J. Flexas, 2002. Regulation of photosynthesis of C3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot., 89: 895-905.
Direct Link - Monneveux, P., C. Sanchez, D. Beck and G.O. Edmeades, 2006. Drought tolerance improvement in tropical maize source populations. Crop Sci., 46: 180-191.
CrossRefDirect Link - Muller, J.E. and M.S. Whitsitt, 1996. Plant cellular responses to water deficit. Plant Growth Regul., 20: 41-46.
CrossRefDirect Link - Swain, P., 1986. Net assimilation rate and its related plant characters of high yielding rice varieties. Indian J. Plant Physiol., 29: 53-60.
Direct Link - Paterson, A.H., J.E. Bowers, R. Bruggmann, I. Dubchak and J. Grimwood et al., 2009. The sorghum bicolor genome and the diversification of grasses. Nature, 457: 551-556.
CrossRefDirect Link - Paterson, A.H., J.E. Bowers, F.A. Feltus, H. Tang, L. Lin and X. Wang, 2009. Comparative genomics of grasses promises a bountiful harvest. Plant Physiol., 149: 125-131.
CrossRefPubMedDirect Link - Peng, Z., Q. Lu and D.P.S. Verma, 1996. Reciprocal regulation of delta 1-pyrroline-5-carboxylate synthetase and proline dehydrogenase genes controls proline levels during and after osmotic stress in plants. Mol. Gen. Genet., 253: 334-341.
PubMedDirect Link - Praba, M.L., J.E. Cairns, R.C. Babu and H.R. Lafitte, 2009. Identification of physiological traits underlying cultivar differences in drought tolerance in rice and wheat. J. Agron. Crop Sci., 195: 30-46.
CrossRefDirect Link - Prasad, P.V., S.R. Pisipati, R.N. Mutava and M.R. Tuinstra, 2008. Sensitivity of grain sorghum to high temperature stress during reproductive development. Crop Sci., 48: 1911-1917.
CrossRefDirect Link - Premachandra, G.S., D.T. Hahn, D. Rhodes and R.J. Joly, 1995. Leaf water relations and solute accumulation in two grain sorghum lines exhibiting contrasting drought tolerance. J. Exp. Bot., 46: 1833-1841.
CrossRefDirect Link - Rampino, P., G. Spano, S. Pataleo, G. Mita and J.A. Napier et al., 2006. Molecular analysis of a durum wheat stay green mutant: Expression pattern of photosynthesis-related genes. J. Cereal Sci., 43: 160-168.
Direct Link - Rontein, D., G. Basset and A.D. Hanson, 2002. Metabolic engineering of osmoprotectant accumulation in plants. Metab. Eng., 4: 49-56.
CrossRefDirect Link - Rosenow, D.T., J.E. Quisenberry, C.W. Wendt and L.E. Clark, 1983. Drought tolerant sorghum and cotton germplasm. Agric. Water Manage., 7: 207-222.
CrossRefDirect Link - Sinclair, T.R. and P.D. Jamiesson, 2008. Yield and grain number of wheat: A correlation or causal relationship?: Author's response to "The importance of grain or kernel number in wheat: A reply to Sinclair and Jamieson" by R.A. Fischer. Field Crop Res., 105: 22-26.
CrossRefDirect Link - Stickler, F.C. and A.W. Pauli, 1961. Leaf removal in grain sorghum. I. Effects of certain defoliation treatments on yield and components of yield. Agron. J., 53: 99-102.
CrossRefDirect Link - Tesso, T., I. Kapran, C. Grenier, A. Snow and P. Sweeney et al., 2008. The potential for crop-to-wild gene flow in sorghum in Ethiopia and Niger: A geographic survey. Crop Sci., 48: 1425-1431.
Direct Link - Tezara, W., O. Marin, E. Rengifo, D. Martinez and A. Herrera, 2005. Photosynthesis and photoinhibition in two xerophytic shrubs during drought. Photosynthetica, 43: 37-45.
Direct Link - Tuinstra, M.R., E.M. Grote, P.B. Goldsbrough and G. Ejeta, 1996. Identification of quantitative trait loci associated with pre-flowering drought tolerance in sorghum. Crop Sci., 36: 1337-1344.
CrossRefDirect Link - Van Rensburg, L. and G.H.J. Kruger, 1993. Differential inhibition of photosynthesis (in vivo and in vitro) and changes in chlorophyll a fluorescence induction kinetics of four tobacco cultivars under drought stress. J. Plant Physiol., 141: 357-365.
CrossRefDirect Link - Verbruggen, N., X.J. Hua, M. May and M. van Montagu, 1996. Environmental and developmental signals modulate proline homeostasis: Evidence for a negative transcriptional regulator. Proc. Nat. Acad. Sci., 93: 8787-8791.
PubMedDirect Link - Wolfe, K.H., M. Gouy, Y.W. Yang, P.M. Sharp and W.H. Li, 1989. Date of the monocot-dicot divergence estimated from chloroplast DNA sequence data. Proc. Nat. Acad. Sci., 86: 6201-6205.
PubMedDirect Link - Xiao, X., X. Xu and F. Yang, 2008. Adaptive responses to progressive drought stress in two Populus cathayana populations. Silva Fennica, 42: 705-719.
Direct Link - Xu, W., P.K. Subudhi, O.R. Crasta, D.T. Rosenow, J.E. Mullet and H.T. Nguyen, 2000. Molecular mapping of QTLs conferring stay-green in grain sorghum (Sorghum bicolor L. Moench). Genome, 43: 461-469.
PubMedDirect Link - Xu, Z.Z., G.S. Zhou, Y.L. Wang, G.X. Han and Y.J. Li, 2008. Changes in chlorophyll fluorescence in maize plants with imposed rapid dehydration at different leaf ages. J. Plant Growth Regulation, 27: 83-92.
CrossRefDirect Link - Yin, C., Y. Peng, R. Zang, Y. Zhu and C. Li, 2005. Adaptive responses of Populus kangdingensis to drought stress. Physiol. Plantarum, 123: 445-451.
CrossRefDirect Link - Juo, A.S.R., 1978. Selected Methods for Soil and Plant Analysis. 2nd Edn., International Institute of Tropical Agriculture, Ibadan, Nigeria, Pages: 52.
Direct Link