
Virgilio Gavicho Uarrota
Plant Science Center, Plant Morphogenesis and Biochemistry Laboratory, Federal University of Santa Catarina, SC, 88040-900, P.O. Box 476, Florianopolis, Brazil
Eder Carlos Schmidt
Department of Cell Biology, Embryology and Genetics, Federal University of Santa Catarina, SC, Florianopolis, 88040-900, P.O. Box 476, Florianopolis, Brazil
Zenilda Laurita Bouzon
Department of Cell Biology, Embryology and Genetics, Federal University of Santa Catarina, SC, Florianopolis, 88040-900, P.O. Box 476, Florianopolis, Brazil
Marcelo Maraschin
Plant Science Center, Plant Morphogenesis and Biochemistry Laboratory, Federal University of Santa Catarina, SC, 88040-900, P.O. Box 476, Florianopolis, Brazil







ABSTRACT
Maize (Zea mays L.) has a wide range of food and non-food applications and is predominantly composed of carbohydrates, mostly in the form of starch, with considerable amounts of protein, as well as some lipids, vitamins and minerals. Both genetic and environmental effects create significant variation in the amount and quality of each of these constituents. Despite the fact that modern agricultural systems have greatly increased the yield of maize with modern cultivars, human domination of earth’s ecosystems is markedly reducing the biodiversity. To access the potential of Brazilian germplasm, seeds of eight maize landraces differing in their phenotypes were cultivated in Anchieta (trial 1) and Ressacada Research Farm, Florianopolis (trial 2) under a low input agricultural system. Alterations in primary metabolite (structural and reserve polysaccharides) were investigated by histochemical analysis and quantified the protein content. Significant protein contents were correlated with regional climatic conditions, cultivars and management practices. Samples of maize landraces of F0 progeny reacted more intensively to Toluidine Blue, Periodic Acid-Schiff and Coomassie Brilliant Blue, suggesting the occurrence of a higher content of acidic polysaccharides, starch, cellulose and proteins in that materials. F0 progeny showed high levels of proteins (e.g., LP and RX varieties). These results, together with our previous observations, confirmed the chemical diversity of maize landraces and can be more prospected to produce protein-rich derivatives due to the concerns on plant resources, biodiversity conservation contributing to local and global food security.
PDF Abstract XML References Citation
Received: July 22, 2011;
Accepted: September 16, 2011;
Published: October 27, 2011
How to cite this article
Virgilio Gavicho Uarrota, Eder Carlos Schmidt, Zenilda Laurita Bouzon and Marcelo Maraschin, 2011. Histochemical Analysis and Protein Content of Maize Landraces (Zea mays L.). Journal of Agronomy, 10: 92-98.
DOI: 10.3923/ja.2011.92.98
URL: https://scialert.net/abstract/?doi=ja.2011.92.98
DOI: 10.3923/ja.2011.92.98
URL: https://scialert.net/abstract/?doi=ja.2011.92.98
INTRODUCTION
Maize (Zea mays L.) is a widely consumed cereal (Uarrota et al., 2011). Worldwide, maize has increased significantly its cultivated area, especially to meet the market demands for animal feed. In Brazil, the cultivation of this cereal back to periods prior to discovery of Portuguese colonization, introducing privileged position and been highlighted in the national economy in recent times. In terms of planted area and grain production, is the second most important cereal in Brazil and only in recent years has lost the first offer for soybean (Vilarinho, 2005).
Maize starches because of their desirable physical and chemical properties are known to be suitable for use as thickeners, extenders, stabilizers, gelling agents, dietary calories and texture modifiers in food formulations and other commercial exploitation (Singh and Ali, 2006). In industry, its seeds are raw material for obtaining various products such as ethyl and butyl alcohol, acetone, whiskey, sugar, dextrin, glucose, oil, beer, vinegar, flour, thick and thin, paints, varnishes and soap. Moreover, it is extensively used in animal feed in the form of grain, green fodder and balanced diet (Adebooye and Singh, 2008; Javed et al., 2006; Ayub et al., 2002). In most countries of Africa, maize has been used as complementary infant food (Oyarekua and Adeyeye, 2009) and many reports have indicated the antioxidative potential of maize leaves (Gholizadeh, 2010); seeds, silks (Kuhnen et al., 2010); stalk, cob and kernels (Agbogidi et al., 2007).
Maize is widely used in food and feed as an energy source due to its high starch content, available in an easily digestible and low cost. However, in terms of the cereal protein source is weak as well as having low average levels (about 10%), its protein is of low biological quality, especially in relation to essential amino acids (Oliveira et al., 2004; Gernah et al., 2011). Small variations in protein content have been observed in samples of maize, depending on soil fertility, climatic conditions and management systems. In addition, the chemical composition of maize grain is variable in different parts of the grain, with higher concentrations of protein in the endosperm (74%) and germ (26%). The proteins of maize can be grouped according to their solubility, as follows: albumins (water soluble), globulins (saline soluble), prolamins (soluble in strong alcohol solution) and glutelin (soluble in alkaline medium) (Kuhnen et al., 2010). The prolamin fraction (α-zein) is the highest concentration in maize, representing 50 to 60% of the total protein (Oliveira et al., 2004).
Maize landraces are potential source of new beneficial alleles that can be used to improve the performance of hybrid varieties in terms of yield, resistance to diseases and other agronomic characteristics because they are in constant co-evolution with environment (Shanbao et al., 2009). The knowledge of protein content in local varieties (maize landraces) in the agricultural sector is important, taking into account the position it occupies in the diet and livelihoods of small farmers. The aim of this study was to analyze histochemically grains of local varieties of maize in order to detect proteins, structural and reserve polysaccharides (starch grains) and quantify the protein content in two generations grown in two regions under low input agricultural systems. The knowledge of the metabolic peculiarities of maize landraces contribute to determine the potential use of raw materials from those genotypes, generating different perspective uses and possibly enhancing their economic value. In fact, the knowledge of the chemical variability is of economic interest, especially to the pharmaceutical, cosmetic and food industries and contributes to the rescue of that germplasm, with the possibility of direct application in breeding programs.
MATERIALS AND METHODS
On farm trials: Maize landrace grains were produced (2008/2009 harvest) under agro-ecological management by small farmers at the far-western region of Santa Catarina State, Southern Brazil (Anchieta county, 26° 31’ 11” S, 53° 20’ 26” W-F0 Progeny). In a second set of experiments, field trials were performed at the Experimental Field of the Plant Science Center (Federal University of Santa Catarina, Florianopolis, southern Brazil -27°35'48" S, 48°32'57" W) by cultivating the F0 maize seeds following typical agro-ecological management adopted in southern Brazil for that cereal (F1-Progeny). Eight maize varieties were studied, namely Lingua de papagaio (LP), Mato grosso-palha roxa (MG), Roxo (RX), Roxo de Emilio (RXE), Palha roxa (PR), Rajado (RJ), Rajado 8 carreiras (R8C) and MPA1. The experimental design followed a completely randomized design. For histochemical analysis were used whole-grain, while for protein content, flour samples were used.
Light microscopy (LM): Samples of maize of the two progenies were fixed in 2.5% paraformaldehyde in 0.1 M (pH 7.2) phosphate buffer overnight. Subsequently, the samples were dehydrated in increasing series of ethanol aqueous solutions (Schmidt et al., 2009). after dehydration, the samples were infiltrated with Historesin (Leica Historesin, Heidelberg, Germany). Sections 5 μm in length were stained with different histochemical techniques and investigated with an Epifluorescent (Olympus BX 41) microscope equipped with Image Q Capture Pro 5.1 Software (Qimaging Corporation, Austin, TX, USA).
Histochemical staining: LM sections were stained as follows: Periodic Acid-schiff (PAS) used to identify neutral polysaccharides (Schmidt et al., 2009), Toluidine Blue (TB-O) 0.5%, pH 3.0 (Merck Darmstadt, Germany) used for acid polysaccharides through a metachromatic reaction (Schmidt et al., 2009) and Coomassie Brilliant Blue (CBB) 0.02% in Clarke’s solution (Serva, Heidelberg, Germany) used for proteins (Schmidt et al., 2010a). Controls consisted of applying solutions to sections without the staining component (e.g., omission of periodic acid application in the PAS reaction).
Scanning Electron Microscopy (SEM): As previously described the defatting prior to preparation of flour samples was carried out by extraction under reflux (ethyl ether, 6 h) in a Soxleth apparatus. The defatted biomass was dried (100°C) and ca. Ten milligram of the flour were mounted on double adhesive carbon coated tape on an aluminum stub. The sample was coated with gold using polaron E5001 SEM coating system. The samples were examined under SEM JSM 6390 LV (JEOL Ltd., Tokyo, Japan, at 10 kV).
Protein content: The protein content was determined based on AOAC (2005) method 991.20 and the protein content was calculated using a following equation.
| (1) |
Where:
| Pr | =Crude protein |
| N | =Nitrogen content |
| 6.25 | =Conversion factor |
Statistical analysis: All the measurements were taken as triplicate. Tukey test (at 5% of probability) was applied to compare means between varieties within progeny (F0 and F1, respectively).
RESULTS
Histochemical analysis: Toluidine Blue (TB-O) staining showed a metachromatic reaction in the cell wall of cells of all varieties (F0 and F1-progenies), indicating the presence of acidic polysaccharides and the nuclei presence was observed (Fig. 1A-D). The vacuole showed metachromatic granulations and it was also observed a orthochromatic cytoplasm.
Samples stained with Periodic Acid-Schiff (PAS) exhibited a strong reaction, suggesting the presence of cellulosic compounds in the cell wall (Fig. 2A-D). This reaction also occurred in the cytoplasm with neutral polysaccharides, especially with many starch grains, the main substance reserve of source of carbohydrates attained in Z. mays L. and cellulose, an important component of the insoluble fiber fraction of the grains. When observed under SEM the starch grains showing various sizes (Fig. 3A-D). However, the reaction was more intense for MPA1 (F1 progeny) and lower intensity for RJ (F0 progeny).
Finally, the cytoplasm of cells when stained with Coomassie Brilliant Blue (CBB) was uniformly marked with the CBB, indicating a homogeneous distribution of organelles or structures rich in protein. With this reaction, the nuclei became more evident (Fig. 4A-D). In addition, cells of the F0 varieties showed more intense staining with CBB, suggesting a greater concentration of proteins in relation to F1 progeny.
Protein content: The results of quantification of grain protein confirmed the histochemical tests, revealing higher levels of these primary metabolites in samples from progeny to progeny over F0 and F1 varieties (Table 1). In the F0 generation, the LP variety was noted for its higher protein content and R8C presented the lower value. In the F1, higher concentrations of proteins were detected for RX and lower value observed for R8C.
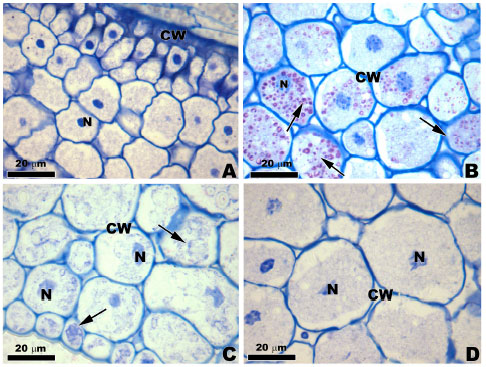 | |
| Fig. 1 (A-D): | Light microscopy of the transversal sections of maize landraces varieties stained with TB-O. (A) RX-1, (B) PR-0, (C) R8C-1 and (D) RJ-0. Note the Cell Wall (CW) shows metachromatic reaction and the visible nuclei (arrows) |
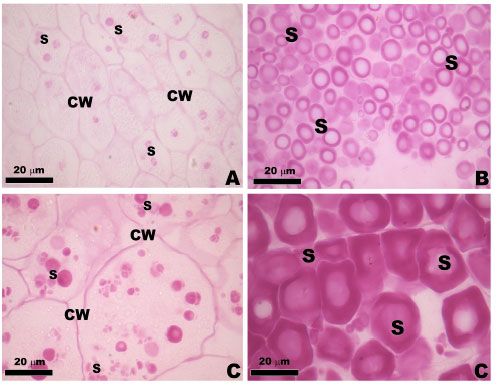 | |
| Fig. 2 (A-D): | Light microscopy of the transversal sections of maize landraces varieties stained with PAS. Observe the positive reaction in cell wall and starch grains (S). (A) RJ-0, (B) R8C-0, (C) MG-1 and (D) MPA1-1 |
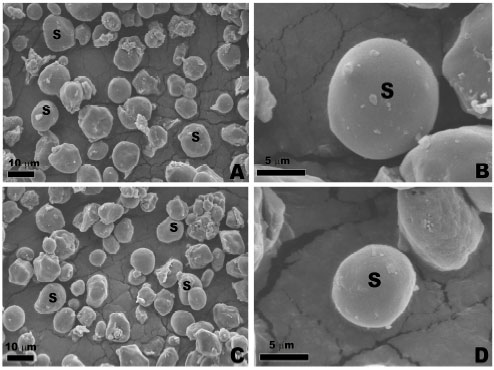 | |
| Fig. 3 (A-D): | Scanning Electron Microscopy (SEM) micrographic images of starch granules extracted from grains of maize landraces. (A) Detail of starch grain of RJ-1. (B) Magnification of previous figure showing spherical shape of the starch granule of individualized RJ-1. (C) Detail of starch grain of MG-1. (D) Magnification of previous figure showing the ellipsoid shape of the individual bead array MG-1 |
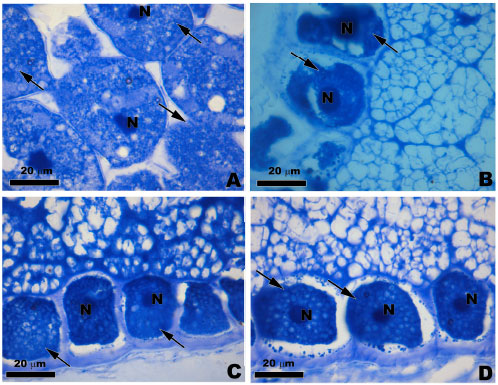 | |
| Fig. 4 (A-D): | Light microscopy of the transversal sections of maize landraces varieties stained with CBB. Showing the presence of proteins in the cytoplasm (arrows) and more intensively in the nucleus (N). (A) LP-1, (B) MPA1-1, (C) PR-0 and (D) RJ-0 |
| Table 1: | Protein content (% Nx6.25) in maize landraces of two progenies |
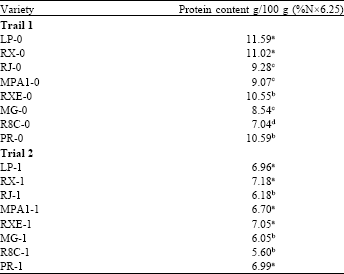 | |
| Different letters indicate significant differences at 5% probability by Tukey test between varieties in the group (F0 and F1 respectively). All data were collected in triplicate (n = 3) | |
It is noteworthy that the F0-F1 progeny was very different between samples suggesting that the soil and climatic differences, for example between the coastal regions (Florianopolis-trial 2) and a far-west of Santa Catarina (Anchieta-trial 1) are strongly related to the synthesis of these compounds of primary metabolism.
DISCUSSION
Several studies have reported protein levels of maize lower than observed in this study. Georget et al. (2008), Wu and Norton (2001) and Prasanna et al. (2001) described contents from 6.9 to 8.9 g/100 g, suggesting that some of the landraces standout as sources of this nutrient. Similar results were obtained by Oliveira et al. (2004) in maize varieties of high quality protein, so that the landraces LP, RX, RXE and PR may eventually be used, for example, in breeding programs aimed at improving the nutritional quality of grains, such as increased protein content and/or preferably for human consumption. In addition, the higher value of maize protein also means making better use for metabolic monogastric animals such as pigs, poultry, fish and horses (Oliveira et al., 2004). It is noteworthy, however, the differential effect of protein content of F0 to F1, an aspect that grains from the generations under study requires more detailed studies, especially considering the diversity of soil and climatic conditions and management of areas suitable for cultivation of that cereal in Santa Catarina state. Despite the landraces in this study have shown high levels of protein, compared to previous studies with these and other genotypes of maize landraces (Kuhnen et al., 2010), this variation may be due to soil and climatic conditions of handling and to some extent, requiring a more detailed study, to determine their protein quality and add value to this materials.
The proteins of maize contain on average about 2% lysine, half the amount recommended by the FAO (Food and Agriculture Organization) for human consumption. Under the nutritional point of view, lysine is the most limiting amino acid in the endosperm of maize, followed by tryptophan (Prasanna et al., 2001). Protein reserves can be found in the endosperm and germ; however, the germ proteins are superior in quality and quantity. Compared with other cereals, the protein quality of maize (32.1%) is similar to that of wheat (38%) and sorghum (32.5%), but very dissimilar to rice (79.3%). The proteins found in the maize landraces under study are important for the small farmers, especially in developing countries with scarce financial resources for acquisition of animal protein.
Studies reporting cellular characteristics of maize landraces are lacking in the literature, otherwise histochemical techniques has been widely applied in many algae studies (Schmidt et al., 2010b). Thus far, documenting the cellular characteristics of maize landrace ultrastructure will led to great understanding and differentiation of primary metabolites of that potential raw materials being cultivated under variety low input agricultural systems.
CONCLUSIONS
Samples of maize landraces of F0 progeny reacted more intensively to TB-O, PAS and CBB, indicating the occurrence of a higher content of acidic polysaccharides in cell wall, starch grains, the main reserve compound and large quantity of proteins. F0 progeny showed high levels of proteins (LP and RX varieties) and can be exploited to generate different protein-rich products. Both varieties of F0 and F1 showed desirable characteristics and can be used to obtain certain products in the industry (e.g., breads, puddings, desserts), which can add more value, the applicability and generate new perspectives for the use of these varieties of maize and encouragement to the small farmer who has developed these varieties for decades.
ACKNOWLEDGMENTS
The authors are grateful for financial support from the National Council for Scientific and Technological Development (CNPq) and Ministry of Science and Technology of Mozambique (MCT). We thank small farmers of Anchieta (Southern Brazil) for providing maize landraces. The authors would like to acknowledge the staff of the Central Laboratory of Electron Microscopy (LCME), Federal University of Santa Catarina, Florianopolis, Santa Catarina, Brazil, for the use of their scanning electron microscope and Fluorescence Laboratory, Biological Science Center-(CCB-BEG) UFSC, for incalculable help.
REFERENCES
- Adebooye, O.C. and V. Singh, 2008. Physico-chemical properties of the flours and starches of two cowpea varieties (Vigna unguiculata (L.) Walp). Innovative Food Sci. Emerg. Technol., 9: 92-100.
CrossRefDirect Link - Agbogidi, O.M., P.G. Eruotor and S.O. Akparobi, 2007. Effects of crude oil levels on the growth of maize (Zea mays L.). Am. J. Food Technol., 2: 529-535.
CrossRefDirect Link - Ayub, M., M.A. Nadeem, M.S. Sharar and N. Mahmood, 2002. Response of maize (Zea mays L.) fodder to different levels of nitrogen and phosphorus. Asian J. Plant Sci., 1: 352-354.
CrossRefDirect Link - Georget, D.M.R., S.A. Barker and P.S. Belton, 2008. A study on maize proteins as a potential new tablet excipient. Eur. J. Pharm. Biopharm., 69: 718-726.
PubMedDirect Link - Gernah, D.I., C.C. Ariahu and E.K. Ingbian, 2011. Effects of malting and lactic fermentation on some chemical and functional properties of maize (Zea mays). Am. J. Food Technol., 6: 404-412.
CrossRefDirect Link - Gholizadeh, A., 2010. Anti-oxidation profile in the leaves of maize inbreds: Elevation in the activity of phenylalanine ammonia lyase under drought stress. J. Plant Sci., 5: 137-145.
CrossRefDirect Link - Javed, H.I., M.A. Masood, S.R. Chughtai, H.N. Malik, M. Hussain and A. Saleem, 2006. Performance of maize genotypes on the basis of stability analysis in Pakistan. Asian J. Plant Sci., 5: 207-210.
CrossRefDirect Link - Kuhnen, S., J.B. Ogliari, P.F. Dias, M. da Silva Santos and A.G. Ferreira et al., 2010. Metabolic fingerprint of Brazilian maize landraces silks (stigmata/styles) using NMR spectroscopy and chemometric methods. J. Agric. Food Chem., 58: 2194-2200.
PubMed - Oyarekua, M.A. and E.I. Adeyeye, 2009. Comparative evaluation of the nutritional quality, functional properties and amino acid profile of co-fermented maize/cowpea and sorghum/cowpea Ogi as infant complementary food. Asian J. Clin. Nutr., 1: 31-39.
CrossRefDirect Link - Prasanna, B.M., S.K. Vasal, B. Kassahun and N.N. Singh, 2001. Quality protein maize. Curr. Sci. Assoc., 81: 1308-1319.
Direct Link - Singh, V. and S.Z. Ali, 2006. In vitro hydrolysis of starches by α-amylase in comparison to that by acid. Am. J. Food Technol., 1: 43-51.
CrossRefDirect Link - Shanbao, Q., W. Yuhua, R. Tingzhao, Y. Kecheng, G. Shibin and P. Guangtang, 2009. Effective improvement of genetic variation in maize lines derived from R08xDonor backcrosses by SSRs. Biotechnology, 8: 358-364.
CrossRefDirect Link - Schmidt, E.C., L.A. Scariot, T. Rover and Z.L. Bouzon, 2009. Changes in ultrastructure and histochemistry of two red macroalgae strains of Kappaphycus alvarezii (Rhodophyta, Gigartinales), as a consequence of ultraviolet B radiation exposure. Micron, 40: 860-869.
CrossRef - Schmidt, E.C., R. dos Santos, P.A. Horta, M. Maraschin and Z.L. Bouzon, 2010. Effects of UVB radiation on the agarophyte Gracilaria domingensis (Rhodophyta, Gracilariales): Changes in cell organization, growth and photosynthetic performance. Micron, 41: 919-930.
CrossRef - Schmidt, E.C., M. Maraschin and Z.L. Bouzon, 2010. Effects of UVB radiation on the carragenophyte Kappaphycus alvarezii (Rhodophyta, Gigartinales): Changes in ultrastructure, growth and photosynthetic pigments. Hydrobiologia, 649: 171-182.
CrossRef - Uarrota, V.G., R.B. Severino and M. Maraschin, 2011. Maize Landraces (Zea mays L.): A new prospective source for secondary metabolite production. Int. J. Agric. Res., 6: 218-226.
CrossRef - Wu, Y.V. and R.A. Norton, 2001. Enrichment of protein, starch, fat and sterol ferulates from corn fiber by fine grinding and air classification. Ind. Crops Prod., 14: 135-138.
CrossRefDirect Link