
N. Nakhro
Microbial Ecology Laboratory, Department of Botany,North-Eastern Hill University, Shillong, Meghalaya-793022, India
M. S. Dkhar
Microbial Ecology Laboratory, Department of Botany,North-Eastern Hill University, Shillong, Meghalaya-793022, India
Journal of Agronomy
Year: 2010 | Volume: 9 | Issue: 3 | Page No.: 102-110







ABSTRACT
A field experiment was conducted to examine the microbial populations and microbial biomass carbon under organic and inorganic farming practices in paddy (variety IR-64) fields. The organic fertilizer used was a mixture of farmyard manure, rock phosphate and neem cake, while on the other hand for inorganic fertilizer, a mixture of urea, single super phosphate and muriate of potash was used. Microbial population counts were analysed from samples collected from the surface (0-15 cm) and sub-surface (15-30 cm) soil depths of the treated plots by soil plate and dilution plate methods for fungi and bacteria, respectively. Results obtained showed that the organically treated plot recorded the maximum microbial population counts (fungal and bacterial) and microbial biomass carbon, followed by the inorganically treated plot and control. A significant variation in fungal population was found between control and treated plots (organic and inorganic) at the surface soil depth, whereas at the sub-surface soil depth it was between all the plots (Tukey’s test at p≤0.05). Organic plot exhibited a significant variation in bacterial population (both the soil depths) with the inorganically treated plot and control (Tukey’s test at p≤0.05). Organic carbon showed significant positive correlation with fungal and bacterial populations (p≤0.05). The application of organic fertilizers increased the organic carbon content of the soil and thereby increasing the microbial counts and microbial biomass carbon. The use of inorganic fertilizers resulted in low organic carbon content, microbial counts and microbial biomass carbon of the soil, although it increased the soil’s NPK level which could be explained by the rates of fertilizers being applied.
PDF Abstract XML References Citation
Received: March 25, 2010;
Accepted: July 22, 2010;
Published: September 28, 2010
How to cite this article
N. Nakhro and M. S. Dkhar, 2010. Impact of Organic and Inorganic Fertilizers on Microbial Populations and Biomass Carbon in Paddy Field Soil. Journal of Agronomy, 9: 102-110.
DOI: 10.3923/ja.2010.102.110
URL: https://scialert.net/abstract/?doi=ja.2010.102.110
DOI: 10.3923/ja.2010.102.110
URL: https://scialert.net/abstract/?doi=ja.2010.102.110
INTRODUCTION
Soil is the habitat of a diverse array of organisms which include both micro flora and fauna. Soil micro organisms play a very important role in soil fertility not only because of their ability to carry out biochemical transformation but also due to their importance as a source and sink of mineral nutrients (Jenkinson and Ladd, 1981). Soil microbes, the living part of soil organic matter, function as a transient nutrient sink and are responsible for releasing nutrients from organic matter for use by plants (e.g., N, P and S). An understanding of microbial processes is important for the management of farming systems, particularly those that rely on organic inputs of nutrients (Smith and Paul, 1990). The soil microbial community is involved in numerous ecosystem functions, such as nutrient cycling and organic matter decomposition, and plays a crucial role in the terrestrial carbon cycle (Schimel, 1995). The microbial communities= potential for rapid growth and turnover, means the microbial community is a more reactive component of a terrestrial ecosystem to external stress than plants and animals (Panikov, 1999). Changes in microbial communities can be use to predict the effects of ecosystem perturbations by organic and conventional management practices (Bending et al., 2000; Bruggen-Van and Semenov, 2000; Poudel et al., 2002).
Agricultural activities such as tillage, intercropping, rotations, drainage, use of pesticides and fertilizers have significant implications for the microorganisms present in the soil (Hengeveld, 1996). The soil microorganisms are sensitive to changes in the surrounding soil (Schinner and Sonnletner, 1996) and have been shown that the microbial population changes after fertilization (Hyman et al., 1990). Fertilizer can directly stimulate the growth of microbial populations as a whole by supplying nutrients and may affect the composition of individual microbial communities in the soil (Khonje et al., 1989). The application of chemical fertilizer generally improves crop production; however, concerns have been raised not only about the severe environmental problems posed by such practices but also about the long term sustainability of such systems (Mader et al., 2002). On the other hand, use of organic materials (e.g., animal manures, crop residues, green manures, etc.) as an alternative source holds promise. Organic farming has been expanding at an annual rate of ca. 20% in the last decade (Lotter, 2003) and has become a mainstream practice for some crops (Anonymous, 2004). Organic applications increased nutrient status, microbial activity and productive potential of soil while the use of only chemical fertilizers in the cropping system resulted in a poor microbial activity and productive potential of soil (Kang et al., 2005). In comparison with conventional farming, organic farming has potential benefits in promoting soil structure formation (Reganold et al., 1987; Pulleman et al., 2003), enhancing soil biodiversity (Doles et al., 2001; Mader et al., 2002; Oehl et al., 2004), alleviating environmental stresses (Horrigan et al., 2002; Macilwain, 2004), and improving food quality and safety (Giles, 2004). The use of chemical fertilizer alone was not effective in improving the nutrient status of soil (Kang et al., 2005).
Changes in soil properties due to cultivation and management and their consequences for production capacity have been a concern of research for many years. Recognition of the importance of soil microorganisms has led to increased interest in measuring the nutrients held in their biomass (Jenkinson and Powlson, 1976). Besides living plants roots and organisms, soil microbial biomass is a living portion of soil organic matter. Soil microbial biomass is considered to act both as the agent of biochemical changes in soil and as a repository of plant nutrients such as nitrogen (N) and phosphorus (P) in agricultural ecosystems (Jenkinson and Ladd, 1981). The changes in soil organic carbon contents are directly associated with changes in microbial biomass carbon and biological activity in the soil. The response to changes in inputs of organic material is much quicker in soil microbial biomass than in soil organic matter as a whole (Powlson and Jenkinson, 1981). Microbial biomass contains labile fraction of organic C and N, which are mineralized rapidly after the death of microbial cells. Soil microbes are typically C- limited (Smith and Paul, 1990); lower microbial biomass in soils from conventional agroecosystems is often caused by reduced organic carbon content in the soil (Fliebach and Mader, 2000). The quantity and quality of organic inputs are the most important factors affecting microbial biomass and community structure (Peacock et al., 2001). Continuous cultivation with frequent tillage results in a rapid loss of OM through increased microbial activity (Shepherd et al., 2001). Recently, microbial biomass and enzyme activities have been recognized as early and indicators of soil stress or productivity changes. Further, there is considerable evidence that they can be used to evaluate the influence of management and land use on soils (Caravaca et al., 2002; Saggar et al., 1999).
The present investigation was conducted with an aim to assess the impact of organic and inorganic farming practices on the physico-chemical properties of soil and the impact of these practices on the dynamics of soil microbial populations and their activities in paddy fields.
MATERIALS AND METHODS
Study site: The investigation was carried out at lowland experimental fields of Agronomy Division, Indian Council of Agricultural Research (ICAR) for North Eastern Hill (NEH) region complex at Umiam, Ri-Bhoi District, Meghalaya, India for a period of two years (2007-08) starting from pretransplanting period (June) till post harvest period (December). The geographical position of the study site is at 25038=N latitude and 91052=E longitude and is situated at an altitude of 850 m.s.l.
Experimental design: The field experiment was a randomized split-plot design, consisting of organic and inorganic fertilizer treatments. According to the type of fertilizer treatments, each of the experimental plots was designated as C, O and IN for control, organically treated and inorganically treated plots respectively. Replicates were maintained for each treatment. The experimental plots were set up at lowland field with each plot consisting of size 7H2 m2 with good drainage systems. Fertilizers were applied according to the dosage recommended by ICAR. The organically treated plot was amended with a mixture of farmyard manure (928 kg plotG1), rock phosphate (262 g plotG1) and neem cake (350 g plotG1), while the inorganically treated plot was amended with a mixture of urea (250 g plotG1), single super phosphate (525 g plotG1) and muriate of potash (100 g plotG1). Control was also set up without the addition of any fertilizers. The seedlings of high yielding variety of paddy IR-64 were transplanted after ploughing properly and addition of fertilizers in 25H25 cm row spacing.
Soil sampling: Soil samples were collected aseptically from the surface (0-15 cm) and the sub-surface (15-30 cm) soil depths in each experimental plot at monthly intervals starting from pretransplanting period (June) till post harvest period (December) for two years (2007 and 2008). From each plot, soil samples were collected randomly and mixed thoroughly to get a homogenous mixture. About 250 g of the soil samples collected were stored at 4°C and was used for microbiological analysis and the remaining were air dried and sieved for the determinations of physico-chemical properties of the soil. The data of two years (2007 and 2008) were used to assess the impact of organic and inorganic farming practices on the dynamics of soil microbial populations and their activities in paddy fields.
Physico-chemical properties: Soil temperature was noted using soil thermometer at the time of sample collection. pH of the samples was taken using an electronic digital pH meter in 1:5 soil- water suspension. The moisture content of the soil samples were determined gravimetrically by weighing, drying in hot air oven at 105°C for 24 h and then reweighing. Organic carbon (C) was determined by the method given by Anderson and Ingram (1993). Total nitrogen (N), available phosphorus (P) and exchangeable potassium (K) were determined by kjeldahl distillation (Jackson, 1973), molybdenum blue method (Allen et al., 1974) and flame photometer method (Jackson, 1973), respectively.
Enumeration of microbial populations: Isolation and estimation of microbial populations, i.e., fungi using soil plate method (Warcup, 1950) and bacteria using dilution plate method (Johnson and Curl, 1972), were carried out using rose Bengal agar media (Martin, 1950) and nutrient agar media for fungi and bacteria, respectively. Media were prepared according to the composition and sterilized in autoclave. Microorganisms were enumerated using soil plate and serial dilution methods on specified media plates and the inoculated plates were incubated at temperatures of 25 and 30°C at duration of 5-7 days and 1-2 days for fungi and bacteria, respectively. After the incubation period, the colony forming units were counted and expressed as CFU gG1 of soil on a moisture free basis.
Microbial biomass carbon. Soil microbial biomass carbon (MBC) was determined using the chloroform- fumigation- extraction method given by Anderson and Ingram (1993).
Statistical analysis. Analysis of Variance (ANOVA) for soil microbial population, microbial biomass carbon and physico chemical properties were analysed using Statistica version 6. As a post hoc analysis Tukey=s test was performed for comparison of treatment means. Simple correlation was performed to test the relationships between the variables.
RESULTS AND DISCUSSION
Physico-chemical properties of soil: The highest organic carbon content was recorded in organically treated plot and the least in control plot at both the surface and sub-surface soil depths. Organic carbon in control plot showed significant variation with organically and inorganically treated plots at the surface soil depth, whereas, at the sub-surface soil depth, significant variation was observed between all the plots according to Tukey’s test (ANOVA) at p≤0.05 (Table 1). Inorganically treated plot amended with a mixture of urea (250 g plotG1), single super phosphate (525 g plotG1) and muriate of potash (100 g plotG1) showed highest content of total nitrogen, available phosphorus and exchangeable potassium followed by the organically treated plot amended with a mixture of farmyard manure (928 kg plotG1), rock phosphate (262 g plotG1) and neem cake (350 g plotG1) and least in control at both the surface and sub-surface soil depths. Total nitrogen in control plot showed significant variation with the organically and the inorganically treated plot according to Tukey’s test (ANOVA) at p≤0.05 at both the soil depths whereas, available phosphorus and exchangeable potassium showed significant variation between all the plots viz., control, organic and inorganic according to Tukey’s test (ANOVA) at p≤0.05 at both the soil depths (Table 1).
Microbial populations: Fungal population was maximum in the organically treated plot and minimum in control plot at both surface and sub-surface soil depths. A significant variation in fungal population was found between control plot and treated plots (organic and inorganic) according to Tukey’s test (ANOVA) at p≤0.05 in the surface soil depth, whereas, in the sub-surface soil depth a significant variation was exhibited between all the plots (Fig. 1).
| Table 1: | Physico-chemical properties of paddy field soil under organic and inorganic fertilizer treatments at surface (0-15 cm) and sub-surface (15-30 cm) soil depths |
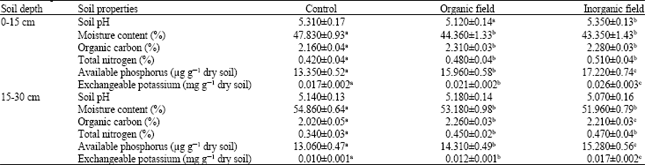 | |
| Mean±SE in columns followed by the same letter do not differ significantly according to Tukey’s test at p≤0.05 | |
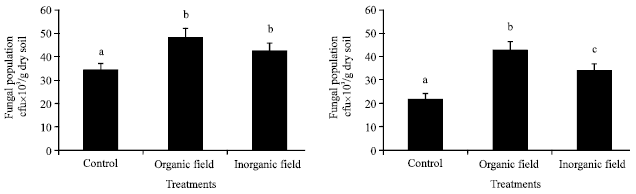 | |
| Fig. 1: | Effect of organic and inorganic fertilizer treatments on fungal population at surface (0-15 cm) and sub-surface (15-30 cm) soil depths in paddy field. Mean±SE with the same letter on top of the bars within treatments do not differ significantly according to Tukey’s test (p≤0.05) |
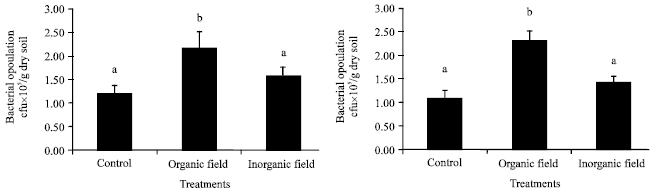 | |
| Fig. 2: | Effect of organic and inorganic fertilizer treatments on bacterial population at surface (0-15 cm) and sub-surface (15-30 cm) soil depths in paddy field. Mean±SE with the same letter on top of the bars within treatments do not differ significantly according to Tukey’s test (p≤0.05) |
The bacterial population in paddy field under different treatments also exhibited similar trend with the maximum and minimum value recorded in the organic and control plots, respectively at both the surface and sub-surface soil depths. Organic plot exhibited a significant variation in bacterial population with the inorganically treated plot and control plot according to Tukey’s test (ANOVA) at p≤0.05 at both the soil depths (Fig. 2).
Microbial biomass carbon: Microbial biomass carbon differed among soils amended with different treatments as well as at different soil depth. The highest microbial biomass carbon was observed in the organically treated plot amended with a mixture of farmyard manure (928 kg plotG1), rock phosphate (262 g plotG1) and neem cake (350 g plotG1) followed by the inorganically treated plot amended with a mixture of urea (250 g plotG1), single super phosphate (525 g plotG1) and muriate of potash (100 g plotG1) and the least in control plot. With respect to the soil depth, the increase was more in the sub-surface (15-30 cm) soil depth than in the surface (0-15 cm) soil depth. Organically treated plot exhibited a significant variation in microbial biomass carbon with control and inorganically treated plot according to Tukey’s test (ANOVA) at p≤0.05 at both the soil depths (Fig. 3).
Physico-chemical properties of soil: Agricultural use of soil affected its chemical properties. The changes in these properties were associated with the organic and inorganic fertiliser management practices at each plot. Soil from the organic plot showed an increase in organic carbon content (Table 1) compared to other plots, this might be due to the addition of organic amendments as they are the sources of nitrogen and carbon to soils. This agrees with finding from several other researchers, Kumar et al. (2000) found that the organic materials, applied alone or in combination with inorganic fertilizer gave greater residual soil fertility in terms of increase in organic carbon content from 0.36 to as high as 0.61% and the available N, P and K in the 2-years cropping cycle.
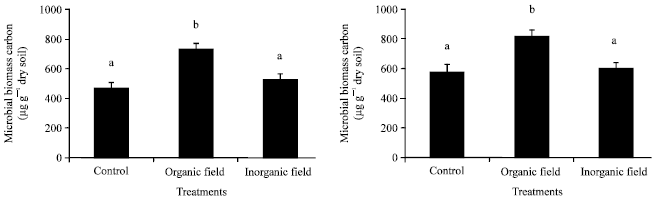 | |
| Fig. 3: | Effect of organic and inorganic fertilizer treatments on microbial biomass carbon at surface (0-15 cm) and sub-surface (15-30 cm) soil depths in paddy field. Mean"SE with the same letter on top of the bars within treatments do not differ significantly according to Tukey’s test (p≤0.05) |
The application of organic manures significantly increased the soil organic carbon content whereas, chemical fertilizers had no effect (Kang et al., 2005). The increase in soil organic carbon content can depend on both organic inputs as well as higher crop residue fall to soil.
It was found that nitrogen content was higher in the inorganic plot compared to organic and control plot. This may be due to the addition of urea (250 g plotG1) which is destined for use as nitrogen release fertiliser. Urea has the highest nitrogen content of all solid nitrogen fertilisers. Also the result is in accordance to the study done by Parham et al. (2002) where manure treated soil was compared with inorganic fertiliser treated soil in which inorganic fertiliser treated soil showed higher nitrogen content. Earlier conclusions were derived from short-term experiments (Suri and Puri, 1997) or from studies with a much higher manure application rate (Agbenin and Goladi, 1997). The lower value of total nitrogen in organic plots could be as a result of crop uptake, immobilisation by microorganisms and nitrogen loss through volatilisation (Defoer et al., 2000). Available P is higher in the inorganically treated plot compared to organic. The increase of available P in the inorganic plot could be due to the addition of single super phosphate (525 g plotG1) compared to rock phosphate (262 g plotG1) added in organic plot. Available P declined in cultivated soil where minimal P application was done. However, higher P was observed with soil that received P fertiliser. Moreover, similar conclusion was drawn by q stating that in most cultivated soils, the soil P is higher than the critical limits (5 μg gG1 in acid soils and 25 μg gG1 in calcareous soils) and the differences could be explained by the rates of fertilizer being applied at each site. Exchangeable K was also found to be higher in inorganic fertiliser treated soil. Application of muriate of potash (100 g plotG1) could have increased the K content in inorganic plot. Also K in cultivated plot decreased where there was no K application and was maintained in soil that received K fertiliser. For soils that did not receive any K fertiliser, the inherent K supply was being depleted due to continuous K outflow extracted from the soil by the crops.
Microbial populations: Result from the present experiment demonstrated that soil microbial population increased in organically amended plots compared to inorganic and control plots which may be due to the addition of organic amendments that might have large impact on the size and activity of microbial population. Bolton et al. (1985) and Ramsey et al. (1986) reported increases in microbial counts in response to fertilization. Long term stubble retention, reduced tillage systems and manuring have been shown to increase microbial biomass carbon and microbial activity in soil (Haines and Ureen, 1990). Mader et al. (2002) reported enhanced soil fertility and higher biodiversity in organic than conventional plots and concluded this may render organic systems less dependent on external inputs. In general, the bacterial population was higher than the fungal population at both the surface as well as the sub- surface soil depths. Parham et al. (2003) reported that cattle manure application promoted the growth of bacteria, but not fungi when compared with the control soil. This was partially attributed to limitations of pH values in the manure treated soils, which were around 5.6, the highest among the treatments evaluated (Parham et al., 2002). Other studies also showed that increased soil pH in the acidic range cause a shift towards dominance of the bacterial community, while fungal communities were unaffected (Frostegard et al., 1993; Pennanen, 2001). The addition of animal or green manures on organic farms provided a significantly greater input of organic carbon, which increased bacterial populations (Fraser et al., 1994).
| Table 2: | Correlation coefficient values of microbial populations (fungi and bacteria CFU) with physico-chemical properties and microbial biomass carbon of paddy field soils under organic and inorganic fertilizer treatments at the surface (0-15 cm) and sub-surface (15-30 cm) soil depths |
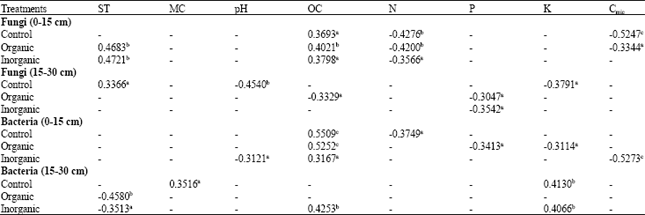 | |
| ST: Soil temperature, MC: Moisture content, OC: Organic carbon, N: Total nitrogen, P: Available phosphorus, K: Exchangeable potassium, Cmic: Microbial biomass carbon. Values marked with a, b and c are significant at p≤0.05, p≤0.01 and p≤0.001, respectively; insignificant values are marked with > | |
Moreover, other researchers have shown that incorporation of organic amendments increased soil microbial activity (Elliott and Lynch, 1994), microbial diversity (Girvan et al., 2004; Grayston et al., 2004) and densities of bacteria (Bruggen-Van and Semenov, 2000). Another possible reason is that manure promoted biological and microbial activities, which accelerated the breakdown of organic substances in the added manure. The enhanced biological activities in the manure treated soil are evidenced by relatively high Cmic content and enzyme activities. As suggested by Jenkinson and Ladd (1981), microbial biomass not only contains a labile pool of nutrients but also drives the cycling of organic matter and nutrients in soil. The fungal population showed positive correlation with organic carbon in all the treatments at the surface soil depth (Table 2) showing that the addition of fertilizers increases the organic carbon content of the soil and thereby, increases fungal population. Diversity is directly correlated with system stability (Odum and Barret, 2005) and diversity increases soil quality by affecting soil agglomeration and increasing fertility (Kirk et al., 2004). Kang et al. (2005) reported the increase in bacterial number in response to chemical fertilizers which may be attributed to a better nutrient status of the soil. There was a decrease in fungal and bacterial population with increase in soil depth which was also reported by Tangjang et al. (2009). The vertical distribution of these specific microbial groups can largely be attributed to the decline in carbon availability with soil depth.
Microbial biomass carbon: Increased microbial biomass carbon content recorded in the organically treated plot might be due to suitable conditions for microbial growth where, development had acted as a good substratum for microbial activity. Fraser et al. (1994) reported a 10-26% increase in microbial biomass under organic management. Microbial biomass carbon in the organically treated soil was significantly higher than other treatments. Leita et al. (1999) indicated that soils treated with FYM and composts showed a significant increase in total organic carbon and biomass carbon in response to the increasing amounts of organic carbon added. A positive effect of organic fertilizers on the microbial biomass nitrogen and the carbon content in the soil was also observed and reported by Cerny et al. (2008). Stimulation of microbial biomass and activities by organic carbon inputs has been well documented in various organic substrates (Goyal et al., 1999; Chowdhary et al., 2000; Garcia-Gil et al., 2000; Peacock et al., 2001; Tu et al., 2005). The present study revealed that microbial biomass carbon increased with soil depth which is also supported by other researchers. Parham et al. (2002) reported that microbial biomass carbon contents decreased with increasing soil depth in all the soils tested with an exception of higher microbial biomass carbon in the 20-30 cm manure treated soil than in the 10-20 cm soil. Sotomayor-Ramírez et al. (2009) also observed that microbial biomass carbon significantly decreased with depth in pasture and forested soils, but this was not true under agriculture soil due to the soil mixing during tillage.
ACKNOWLEDGMENTS
The authors are grateful to the Head, Department of Botany, North-Eastern Hill University, Shillong, Meghalaya for providing laboratory facilities and Division of Agronomy, ICAR, Umiam, Ri-Bhoi district, Shillong for their cooperation and support in setting up the experimental plots. The first author is grateful to Rajiv Gandhi National Fellowship for SC/ST, UGC for financial support.
REFERENCES
- Agbenin, J.O. and J.T. Goladi, 1997. Carbon, nitrogen and phosphorus dynamics under continuous cultivation as influenced by farmyard manure and inorganic fertilizers in the savanna of Northern Nigeria. Agric. Ecosyst. Environ., 63: 17-24.
CrossRef - Allen, S.E., 1974. Chemical Analysis of Ecological Materials. Blackwell Scientific Publications, London, UK., Pages: 565.
Direct Link - Bending, G.D., C. Putland and F. Rayns, 2000. Changes in microbial community metabolism and labile organic matter fractions as early indicators of the impact of management on soil biological quality. Biol. Fertil. Soils, 31: 78-84.
CrossRef - Bolton, H., L.F. Elliott, R.I. Ppendick and D.F. Bezdicek, 1985. Soil microbial biomass and selected soil enzyme activities: Effect of fertilization and cropping practices. Soil Biol. Biochem., 17: 297-302.
Direct Link - Caravaca, F., G. Masciandaro and B. Ceccanti, 2002. Land use in relation to soil chemical and biochemical properties in a semiarid Mediterranean environment. Soil Tillage Res., 68: 23-30.
CrossRef - Chowdhary, M.A.H., K. Kouno, T. Ando and T. Nagaoka, 2000. Microbial biomass, S mineralization and S uptake by African millet from soil amended with various composts. Soil Biol. Biochem., 32: 845-852.
CrossRef - Dobermann, A. and T.H. Frairhurst, 2000. Rice: Nutrient Disorders and Nutrient Management. 1st Edn., PPIC and IRRI, Philippines, Singapore, ISBN: 981-04-2742-5, Pages: 191.
Direct Link - Doles, J.L., R.J. Zimmerman and J.C. Moore, 2001. Soil microarthropod community structure and dynamics in organic and conventionally managed apple orchards in Western Colorado, USA. Applied Soil Ecol., 18: 83-96.
CrossRef - Fraser, P.M., R.J. Haynes and P.H. Williams, 1994. Effects of pasture improvement and intensive cultivation on microbial biomass, enzyme activities, and composition and size of earthworm population. Biol. Fertil. Soils, 17: 185-190.
CrossRef - Frostegard, A., E. Baath and A. Tunlid, 1993. Shifts in the structure of soil microbial communities in limed forests as revealed by phospholipids fatty acid analysis. Soil Biol. Biochem., 25: 723-730.
CrossRef - Garcia-Gil, J.C., C. Plaza, P. Soler-Rovira and A. Polo, 2000. Long-term effects of municipal solid waste compost application on soil enzyme activities and microbial biomass. Soil Biol. Biochem., 32: 1907-1913.
CrossRefDirect Link - Girvan, M.S., J. Bullimore, A.S. Ball, J.N. Pretty and A.M. Osborn, 2004. Response of active bacterial and fungal communities in soil under winter wheat to different fertilizer and pesticide regiments. Applied Environ. Microbiol., 70: 2692-2701.
CrossRef - Goyal, S., K. Chander, M.C. Mundra and K.K. Kapoor, 1999. Influence of inorganic fertilizers and organic amendments on soil organic matter and soil microbial properties under tropical conditions. Biol. Fert. Soils, 29: 196-200.
CrossRefDirect Link - Grayston, S.J., C.D. Campbell, R.D. Bardgett, J.L. Mawdsley and C.D. Clegg et al., 2004. Assessing shifts in microbial community structure across a range of grasslands of differing management intensity using CLPP, PLFA and community DNA techniques. Applied Soil Ecol., 25: 63-84.
CrossRefDirect Link - Horrigan, L., S.L. Robert and P. Walker, 2002. How sustainable agriculture can address the environmental and human health harms of industrial agriculture. Environ. Health Perspect., 110: 445-456.
CrossRefPubMedDirect Link - Jackson, M.L., 1973. Soil Chemical Analysis. 1st Edn., Prentice Hall Ltd., New Delhi, India, Pages: 498.
Direct Link - Jenkinson, D.S. and D.S. Powlson, 1976. The effect of biocidal treatments on metabolism in soil- V. A method for measuring soil biomass. Soil Biol. Biochem., 8: 209-213.
CrossRef - Kang, G.S., V. Beri, O.P. Rupela and B.S. Sidhu, 2005. A new index to assess soil quality and sustainability of wheat based cropping systems. Biol. Fertil. Soils, 41: 389-398.
CrossRefDirect Link - Khonje, D.J., E.C. Varsa and B. Klubek, 1989. The acidulation effects of nitrogenous fertilizers on selected chemical and microbiological properties of soil. Commun. Soil Sci. Plant Anal., 20: 1377-1395.
CrossRef - Kirk, J.L., L.A. Beaudette, M. Hart, P. Moutoglis, J.N. Klironomos, H. Lee and J.T. Trevors, 2004. Methods of studying soil microbial diversity. J. Microbiol. Methods, 58: 169-188.
CrossRefDirect Link - Kumar, V., B.C. Ghosh and R. Bhat, 2000. Complementary effect of crop wastes and inorganic fertilizers on yield, nutrient uptake and residual fertility in mustard (Brassica juncea)-rice (Oryza sativa) cropping sequence. Indian J. Agric. Sci., 70: 69-72.
Direct Link - Leita, L., M.D. Nobili and C. Mondini, 1999. Influence of inorganic and organic fertilization on soil microbial biomass, metabolic quotient and heavy metal bioavailability. Biol. Fertil. Soils, 28: 371-376.
CrossRefDirect Link - Martin, J.P., 1950. Use of acid, rose bengal and streptomycine in the plate method for estimating soil fungi. Soil Sci., 69: 215-232.
CrossRef - Maeder, P., A. Fliessbach, D. Dubois, L. Gunst, P. Fried and U. Niggli, 2002. Soil fertility and biodiversity in organic farming. Science, 296: 1694-1697.
CrossRefDirect Link - Oehl, F., E. Sieverding, P. Mader, D. Dubois, K. Ineichen, T. Boller and A. Wiemken, 2004. Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia, 138: 574-583.
CrossRefDirect Link - Panikov, N.S., 1999. Understanding and prediction of soil microbial community dynamics under global change. Applied Soil Ecol., 11: 161-176.
CrossRef - Parham, J.A., S.P. Deng, W.R. Raun and G.V. Johnson, 2002. Long-term cattle manure application in soil: I. Effecct on soil phosphorous levels, microbial biomass C and dehydrogenase and phosphatase activities. Biol. Fertil. Soils, 35: 328-337.
Direct Link - Parham, J.A., S.P. Deng, H.N. Da, H.Y. Sun and W.R. Raun, 2003. Long-term cattle manure application in soil. II. Effect on soil microbial populations and community structure. Biol. Fertil. Soils, 38: 209-215.
CrossRefDirect Link - Peacock, A.D., M.D. Mullen, D.B. Ringelberg, D.D. Tyler, D.B. Hedrick, P.M. Gale and D.C. White, 2001. Soil microbial community responses to dairy manure or ammonium nitrate applications. Soil Biol. Biochem., 33: 1011-1019.
CrossRefDirect Link - Pennanen, T., 2001. Microbial communities in boreal coniferous forest humus exposed to heavy metals and changes in soil pH- a summary of the use of phospholipid fatty acids biolog and 3H-thymidine incorporation methods in field studies. Geoderma, 100: 91-126.
CrossRef - Powlson, D.S. and D.S. Jenkinson, 1981. A comparision of organic matter, biomass, adenosine triphosphate and mineralisable nitrogen contents of plowed and direct-drilled soils. J. Agric. Sci., 97: 713-721.
CrossRef - Poudel, D.D., W.R. Horwarth, W.T. Lanini, S.R. Temple and A.H.C. Van-Bruggen, 2002. Compariso of soil N availability and leaching potential, crop yields and weeds in organic, low-input and conventional farming systems in northern California. Agric. Ecosyst. Environ., 90: 125-137.
CrossRef - Ramsey, A.J., R.E. Stannard and G.J. Churchman, 1986. Effect of conversion from ryegrass pasture to wheat cropping on aggregation and bacterial populations in a silt loam soil in New Zealand. Aus. J. Soil Res., 24: 253-264.
CrossRef - Reganold, J.P., L.F. Elliott and Y.L. Unger, 1987. Long-term effects of organic and conventional farming on soil erosion. Nature, 330: 370-372.
CrossRefDirect Link - Saggar, S., M.D. McIntosh, C.B. Hedley and H. Knicker, 1999. Changes in soil microbial biomass, metabolic quotient and organic matter turnover under Hieracium (H. Pilosella L.). Biol. Fert. Soil, 30: 232-238.
CrossRef - Shepherd, T.G., S. Saggar, R.H. Newman, C.W. Ross and J.L. Dando, 2001. Tillage-induced changes in soil structure and soil organic matter fractions. Aust. J. Soil Res., 39: 465-489.
CrossRef - Sotomayor- Ramirez, D., Y. Espinoza and V. Acosta-Martinez, 2009. Land use effects on microbial biomass C, β- glucosidase and β- glucosaminidase acivities, and availability, storage, and age of organic C in soil. Biol. Fertil. Soils, 45: 487-497.
CrossRef - Tu, C., J.B. Ristaino and S. Hu, 2006. Soil microbial biomass and activity in organic tomato farming systems: effects of organic inputs and surface mulching. Soil Biol. Biochem., 38: 247-255.
CrossRef - Bruggen-Van, A.H.C. and A.M. Semenov, 2000. In search of biological indicators for soil health and disease suppression. Applied Soil Ecol., 15: 13-24.
CrossRefDirect Link - Warcup, J.H., 1950. The soil-plate method for isolation of fungi from soil. Nature, 166: 117-118.
CrossRef
Sanjib ketan Sahoo Reply
My suggetion is further experiment may be done by other organic resources because due to availablity of organic manure please go for other resourse but i am well greate full to you .
Neilhousano Nakhro
Thank you so much for your valuable suggestion. i am also interested in doing further experiments with other organic resources as well.
kidanu kelbessa Reply
I am conducting my thesis on the current status and carbon sequestration potential of Integrated Nutrient Management Practice. I need the incorporation of Organic fertilizer compared to the Inorganic fertilizer effect on 1. carbon sequestration
2. particulate Organic matter
3. MBC and POCY analysis