
Murat Ali Turan
Department of Soil Science, Faculty of Agriculture, Uludag University, 16059 Bursa, Turkey
Nilufer Turkmen
Department of Soil Science, Faculty of Agriculture, Ankara University, (361 1 C) Ankara, Turkey
Nilgun Taban
Department of Soil Science, Faculty of Agriculture, Ankara University, (361 1 C) Ankara, Turkey
Journal of Agronomy
Year: 2007 | Volume: 6 | Issue: 2 | Page No.: 378-381







ABSTRACT
The effects of NaCl salinity on the growth, stomatal resistance, proline, total chlorophyll and Na, Cl and K concentrations of lentil (Lens calinaris L. cv. Sultan) were investigated. A greenhouse experiment was conducted where NaCl was applied 25, 50 and 100 mM to Aridisol soil. The addition of NaCl resulted in a decrease of the dry weight of plants. Stomatal resistance and concentrations of proline, Na and Cl were increased by treatment with NaCl, while K and total chlorophyll concentrations were decreased.
PDF Abstract XML References
How to cite this article
Murat Ali Turan, Nilufer Turkmen and Nilgun Taban, 2007. Effect of NaCl on Stomatal Resistance and Proline, Chlorophyll, Na, Cl and K Concentrations of Lentil Plants. Journal of Agronomy, 6: 378-381.
DOI: 10.3923/ja.2007.378.381
URL: https://scialert.net/abstract/?doi=ja.2007.378.381
DOI: 10.3923/ja.2007.378.381
URL: https://scialert.net/abstract/?doi=ja.2007.378.381
INTRODUCTION
Salinity is the main environmental factor accountable for decreasing crop productivity in many areas of the world especially in arid and semi-arid regions (De Pascale and Barbieri, 1997). On the other hand, not only uncontrolled irrigation but also poor-quality water may cause salinity (Tanji, 1990; Maas and Grattan, 1999). It was estimated that about one-third of irrigated land has been affected by salinity (Shannon, 1984).
Salinity reduces growth and yield of the non-halophytes plants by decreasing the availability of water to the roots due to the osmotic effect of external salt and by toxic effects of excessive salt accumulation within the plant (Mass and Hoffman, 1977; Turan et al., 2007a).
NaCl toxicity, the predominant form of salt in most saline soils, enhances the Na and Cl contents and consequently affects the absorption of other mineral elements (Greenway and Munns, 1980). It is stated that high levels of Na inhibits Ca and K absorption, which results in a Na/K antagonism (Al-Karaki, 1997) and net photosynthesis is affected strongly by NaCl-saline conditions, which is related directly to the closure of stoma as well as to low intercellular CO2 levels.
Under salt stress, plants have evolved complex mechanisms allowing for adaptation to osmotic and ionic stress caused by high salinity. These mechanisms include osmotic adjustment by accumulation of compatible solutes such as proline and lowering the toxic concentration of ions in the cytoplasm by restriction of Na influx or its sequestration into the vacuole and/or its extrusion. However, the significance of proline accumulation in osmotic adjustment is still debated and varies according to the species (Lutts et al., 1996; Rodriguez et al., 1997; Turan et al., 2007a and b).
The lentil is an annual winter/summer grain legume, needing low to medium rainfall. Lentils (Lens culinaris) have been used as a staple food since the beginning of agriculture, in south west Asia. They are an important source of protein for people in the Mediterranean, Africa, the Middle East, Southern Asia and South America.
Currently the lentil ranks about fifth in the world production of pulses. Food and Agriculture Organisation statistics indicate that the lentil is grown on about 3.4 million hectares. Total world production was 2.7 million tonnes in 1993. India produces nearly one-third of the world’s annual production, largely for domestic consumption. Other major producers are Turkey, Canada, Bangladesh, Syria, Nepal, United States of America, Iran and New Zealand. Turkey is the largest exporter of lentils in the world and produces 0.3-0.5 million tonnes of split lentils per year (Anonymous, 2005).
Lentil grown in Turkey might be exposed to salt stress because of dry-growing and low rainfall areas. Therefore, objective of this research is to examine the influence of soil salinity on the growth and stomatal resistance, proline, total chlorophyll and some mineral element concentrations of lentil plants.
MATERIALS AND METHODS
An experiment was conducted under greenhouse conditions (humidity 65-75%, air temperature 25-30°C and neutral light intensity 340-450 μmol m-2 sec-1) in Ankara-Turkey, between April 1st to May 30th, 2006, with lentil (Lens calinaris L. cv: Sultan) plants. Lentil genotype was very sensitive to the salt stress. The experimental soil taken from Aridisol great soil group was calcareous (212 g kg-1), clay loam in texture (clay 30.49%, silt 41.88% and sand 27.63%), slightly alkaline (pH 8.04, EC 0.108 dS cm-1; both in 1:2.5 water extract). The soil sample has 42.9 mg kg-1 exchangeable Na and 6.37 mg kg-1 water extractable Cl.
The experimental soil (2000 g) was placed into the pots and soil was salinised at the levels of 25, 50 and 100 mM NaCl, before seeds were sown into the pots. Plants grown without NaCl addition were taken as control. For basal fertilizers, 100 mg N kg-1 as ammonium nitrate, 60 mg P kg-1 as triple super phosphate and 75 mg K kg-1 as potassium sulfate were applied to the pots.
Eight seeds of lentil were planted in each pot, thinned to six per pot after germination. The experimental plants were irrigated with de-ionized water, when moisture level decreased below the 60% of field water capacity. Eight weeks after germination, the vegetative growth (shoots plus leaves) was harvested. After weighting the fresh material, plants were washed and 2.0 g of samples were taken for total chlorophyll and proline determinations. The remaining plant samples were dried at 65°C and digested with HNO3:HClO4 acid mixture (4:1) in order to determine dry matter and the concentrations of Na, Cl and K.
Sodium and potassium concentrations were determined by flame photometer (Eppendorf Elex 6361 model) described by Miller (1998). Chloride was analyzed by precipitation as AgCl and titration according to Johnson and Ulrich (1959).
For determination of proline concentration the samples were extracted and quantified in Shimadzu UV-1201 model spectrophotometer at 520 nm, as described by Bates et al. (1973).
Total chlorophyll of plants was extracted in 80% (v/v) aqueous acetone and absorption was measured in a Shimadzu UV-1201 model spectrophotometer at 645 and 663 nm (Arnon, 1949).
Stomatal resistance was determined during the span of time (14 and 15 h) before harvest by use of a steady-state porometer (EA 540-026 AP4 model) attached to the abaxial side of leaves. The readings were taken on six fully-expanded leaves per treatment, situated at the different position of the canopy. Likewise, temperature (27°C), relative humidity (65%) and light intensity (340-400 μmol m-2 sec-1) were also recorded.
The experiment was arranged in a completely randomized design with three salt treatments and three replicates per treatment. The data were analyzed statistically by using Minitab package program (Minitab Release 10.51) and treatment means were compared using the MSTAT package program (Version 3.00) for Duncan’s Multiple Range Test.
RESULTS
Increasing NaCl concentration gradually decreased the growth of lentil plant, expressed as dry weight (Table 1). A slight reduction of dry weight was observed at the lowest level of NaCl as 12.1% in comparison to the control. In parallel to the increase NaCl concentration the effect of salinity was significant for lentil, i.e., growth reduction of plant at 100 mM NaCl treatment was 45.2% compare with the control.
NaCl salinity exhibited controversial effects on ion concentrations of lentil plants (Table 2). Salt treatments increased significantly (p<0.001) Na and Cl concentrations of plants, whereas potassium concentration and K/Na ratio of plants were decreased.
Total chlorophyll and proline concentrations and stomatal resistance of lentil plants were also affected significantly (p<0.001) by salinity (Table 3). The addition of NaCl in the soil decreased total chlorophyll concentration in plants. On the contrary increased proline concentration and stomatal resistance were determined.
| Table 1: | Effects of NaCl on dry weight of lentil plants |
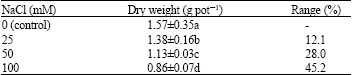 | |
| Means followed by the different letter(s) are significantly different (Duncan’s Multiple Range Test, p<0.01) | |
| Table 2: | Effects of NaCl on Na, Cl and K concentrations and K/Na ratio of lentil plants |
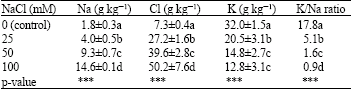 | |
| ***Significant and p<0.001, Means followed by the same letter(s) are not significantly different (Duncan’s Multiple Range Test, p<0.01) | |
| Table 3: | Effects of NaCl on total chlorophyll and proline concentrations and stomatal resistance of lentil plants |
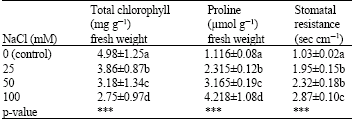 | |
| ***Significant and p<0.001, Means followed by the same letter(s) are not significantly different (Duncan’s Multiple Range Test, p<0.01) | |
DISCUSSION
Saline soils and saline irrigations constitute a serious production problem for vegetable crops as these are known to suppress plant growth (Lauchli, 1984; Cusido et al., 1987; Turan et al., 2007a and b). The present study demonstrates that salinity, particularly when applied at high levels (50 and 100 mM NaCl), adversely affect the growth of lentil plants.
In the experiment, salt stress, induced by increasing NaCl in the soil solution, resulted in an increase in Na and Cl concentrations. There is an overwhelming amount of evidence to indicate that NaCl-induced-salinity increases Na and Cl uptake by plants (Chavan and Karadge, 1986; Alpaslan et al., 1999; Taban et al., 1999). The data have shown that K uptake and transport to the aerial part of the lentil plant leaves were significantly reduced with increased salinity. This implies a competition between Na and K absorption in lentil plants, resulting in a Na/K antagonism (Siegel et al., 1980). The reduction in K uptake caused by Na is likely to be the result of the competitive intracellular influx of both ions (Cerda et al., 1995). It is well established that many K transport systems have significant affinity for Na (Schachtman and Liu, 1999).
Measurement of stomatal resistance provides a sensitive tool for determining the degree of stress in plants. In the present study, the measured stomatal resistance at the end of the experimental period showed that under salinity lentil plants close their stomata which lead to enhanced stomatal resistance. Similar results were reported by Turan et al. (2007b). Stomata closure is known to be an effective mechanism for economical water utilisation under salt stress and limitation of the harmful salt ions uptake (Hasegawa et al., 2000).
One of the most important mechanisms exerted by higher plants under salt-stress conditions is the accumulation of compatible solutes such as proline. The salt treatments induced an increase in proline concentration in lentil plants. Proline accumulation under salinity may contribute to osmotic adjustment, protecting cell structure and function and/or may serve as metabolic or energetic reserve in plants (Lewitt, 1980; Harivandi et al., 1982; Chowdhury et al., 1993; Madan et al., 1994).
The total chlorophyll concentration of lentil leaves was reduced by increasing the level of NaCl applied. Similar results were reported for total leaf chlorophyll concentration of Phaseolus vulgaris (Petolino and Leone, 1980), rice (Yeo and Flowers, 1983) and wheat and bean (Turan et al., 2007a and b). This effect of NaCl was attributed to a salt-induced weakening of protein-pigment-lipid complex (Strogonov et al., 1970) and/or increasing chlorophyllase (EC: 3.1.1.14) activity (Stivsev et al., 1973).
Consequently, soil salinity significantly inhibited the growth and resulted in a decrease of dry matter of lentil. However, NaCl caused significant variations in stomatal resistance, proline and total chlorophyll concentrations and mineral compositions by uptaking excess mineral elements.
ACKNOWLEDGMENTS
The authors thank Dr. Arzu Akpınar Bayızıt and Mrs. Cigdem Vardar for technical assistance at this research.
REFERENCES
- Al-Karaki, G.N., 1997. Barley response to salt stress at varied levels of phosphorous. J. Plant Nutr., 20: 1635-1643.
CrossRefDirect Link - Alpaslan, M., A. Gunes and S. Taban, 1999. Salinity resistance of certain rice (Oryza sativa L.) varieties. Turk. J. Biol., 23: 499-506.
Direct Link - Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Cerda, A., J. Pardines, M.A. Botella and V. Martinez, 1995. Effect of potassium on growth, water relations and the organic solute contents for two maize grown under saline conditions. J. Plant Nutr., 18: 839-851.
Direct Link - Chavan, D. and B.A. Karadge, 1986. Growth, mineral nutrition, organic constituents and rate of photosynthesis in Sesbania grandiflora L. grown under saline conditions. Plant Soil, 93: 395-404.
CrossRefDirect Link - Chowdhury, J.B., S. Jain and R.K. Jain, 1993. Biotechnological approaches for developing salt-tolerant crops. J. Plant Biochem. Biotechnol., 2: 1-7.
Direct Link - Cusido, R.M., J. Palazon, T. Altobella and C. Morales, 1987. Effect of salinity on soluble protein, free amino acids and nicotine contents in Nicotiona rustica L. Plant Soil, 102: 55-60.
CrossRefDirect Link - Greenway, H. and R. Munns, 1980. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol., 31: 149-190.
CrossRefDirect Link - Hasegawa, P.M., R.A. Bressan, J.K. Zhu and H.J. Bohnert, 2000. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol., 51: 463-499.
CrossRefDirect Link - Lutts, S., J.M. Kinet and J. Bouharmont, 1996. Effects of salt stress on growth, mineral nutrition and proline accumulation in relation to osmotic adjustment in rice (Oryza sativa L.) cultivars differing in salinity tolerance. J. Plant. Grow. Regul., 19: 207-218.
CrossRef - Maas, E.V. and G.J. Hoffman, 1977. Crop salt tolerance-current assessment. J. Irrig. Drainage Div., 103: 115-134.
Direct Link - Madan, S., H.S. Nainawatee, S. Jain, R.K. Jain, M.S. Malik and J.B. Chowdhury, 1994. Leaf position-dependent changes in proline, proline-5-carboksilate reductase activity and water relations under salt stress in genetically stable salt-tolerant samaclones of Brassica juncea L. Plant Soil., 163: 151-156.
- Rodriguez, H.G., J.K.M. Roberts, W.R. Jordan and M.C. Drew, 1997. Growth, water relations and accumulation of organic and inorganic solutes in roots of maize seedlings during salt stress. Plant Physiol., 113: 881-893.
Direct Link - Schachtman, D. and W. Liu, 1999. Molecular pieces to the puzzle of the interaction between potassium and sodium uptake in plants. Trends. Plant Sci., 4: 281-287.
CrossRef - Siegel, S.M., B.Z. Siegel, J. Massey, P. Lahne and J. Chen, 1980. Growth of corn in saline waters. Physiologia Plantarum, 50: 71-73.
CrossRefDirect Link - Turan, M.A., V. Katkat and S. Taban, 2007. Variations in proline, chlorophyll and mineral elements contents of wheat plants grown under salinity stress. J. Agron., 6: 137-141.
CrossRefDirect Link - Turan, M.A., K. Vahap and T. Suleyman, 2007. Salinity-induced stomatal resistance, proline, chlorophyll and ion concentrations of bean. Int. J. Agric. Res., 2: 483-488.
CrossRefDirect Link - Yeo, A.R. and T.J. Flowers, 1983. Varietal differences in the toxicity of sodium ions in rice leaves. Physiologia Plantarum, 59: 189-195.
CrossRefDirect Link