
S. El messoussi
Department of Biology, Semlalia, BP 2319 Marrakech, Morocco
H. Hafid
Department of Biology, Semlalia, BP 2319 Marrakech, Morocco
A. Lahrouni
Department of Physics, Semlalia, BP 2319 Marrakech, Morocco
M. Afif
Department of Mathematics, Faculty of Sciences,
Semlalia, BP 2319 Marrakech, Morocco
Journal of Agronomy
Year: 2007 | Volume: 6 | Issue: 2 | Page No.: 374-377







ABSTRACT
Effects of the temperature on immature stage developmental times of the medfly Ceratitis capitata (Diptera; Tephritidae) were used to study the dynamics of population with FRUFLY model. To test the model in the field, flight activity was studied using para-pheromone traps. The FRUFLY model simulations agree with experimental data results of the insect collected by para-pheromone traps and predicts the appearance of the various generations of adults with time FRUFLY simulation modelling is an important tool for identifying insect pest population's size and can help to determine the urgency of action and evaluating options for management.
PDF Abstract XML References
How to cite this article
S. El messoussi, H. Hafid, A. Lahrouni and M. Afif, 2007. Simulation of Temperature Effect on the Population Dynamic of the Mediterranean Fruit Fly Ceratitis capitata (Diptera; Tephritidae). Journal of Agronomy, 6: 374-377.
DOI: 10.3923/ja.2007.374.377
URL: https://scialert.net/abstract/?doi=ja.2007.374.377
DOI: 10.3923/ja.2007.374.377
URL: https://scialert.net/abstract/?doi=ja.2007.374.377
INTRODUCTION
It has long been recognized that temperature is the most important factor determining development rates of the immature stages and the adult maturation rates of the majority of insects. As a result, numerous attempts have been made to determine the temperature-development rate relationships of the tephritid fruit flies because of their importance in predicting phonological events in the field for ecological and pest management purposes, the optimization of mass rearing procedures under constant conditions and the construction of computer simulation models of population processes. If the insect is able to develop many generations per year, the probability of building up a large population during the season and becoming a pest increases. C. capitata or medfly (Diptera; Tephritidae) is a polyphagous insect. Larvae feed on fruits.
Many references are available on the temperature-development rate relationships of the medfly pupae placed in constant conditions (Bodenheimer, 1925; Shoukry and Hafez, 1979; Tassan et al., 1983; Vargas et al., 1984; Crovetti et al., 1986). In more recent studies, Duyck and Quilici (2002) used thermal summation to model data on pupae medfly development times. The time required for 50% of individuals to achieve development was determined at the following constant temperatures: 15, 20, 25, 30 and 35°C (±1°C).
The most commonly used method of predicting the development rates of individual life stages is the temperature summation model. This approach has been made to determine a construction of computer simulation models of population processes based on the assumption that each life stage needs a quantity of heat (normally expressed as degrees-day) to complete development. The degree-day model consider that above some lower threshold for development, temperature-development rate relationships are linear (y = αt+β) and therefore, a constant number of heat units above this threshold are needed to complete development. The linear model was used to determine the threshold for development by extrapolation of the linear regression line back to the x-axis and the temperature sum or thermal constant K (the total quantity of thermal energy required to complete development). The threshold temperature and the thermal constant are then used to determine the rate of development in the field by summation of degree-days exceeding the threshold temperature (Dent, 1997). The data used to fit the model by a linear regression against temperature where the mean development rates for each temperature.
When using the thermal summation model to calculate development times in fluctuating daily temperature regimes, the number of day-degrees per day (°D) can be determined from the formula (Tmax + Tmin)/2)-Tb, as long as the minimum temperature (Tmin) is above the development threshold (Tb). The linear model was chosen because it is easy to determine, requires a minimal amount of data and is acceptable for field prediction purposes where field temperature externs are rare.
The goal of this research is to develop an environmental simulator modeling all relevant entities, medfly dynamic population and environmental factors in a new model named FRUTFLY. The desired environmental model would permit a qualitatively correct estimate of the insect population all year round. This would increase the amount of information about the medfly's host fruit infestation state, helping to decide the best time for their control.
MATERIALS AND METHODS
FRUFLY model: A dynamic population model, FRUFLY, for the complete life cycle of fruit fly pest, (Diptera: Tephritidae), is described (El messoussi et al., 2003; El messoussi et al., unpublished data). Adult population dynamics from emergence to oviposition are based on biotic and abiotic factors. These factor-dependent development and age-dependent advancement determine adult population dynamics and oviposition.
The model determines an optimal behavior of different system components during the life cycle with an adjustment by a limiting factor like temperature, humidity, parasitism and predation. The FRUFLY model consists of a set of distinct agents, each representing one phase in the insects life cycle. An insects transition from a phase to another corresponds to the creation of a new agent depicting the insect in the following phase and the departure of the agent from the completed one.
The biotic and abiotic factors are represented by functions with values ranging between 0 and 1. Thus they can act on the state variables of the fly are in a positive way (if their values are close to 1) or in a stressing way if they are close to 0. The outputs of the model can be presented in the forms of tables or curves which represent the daily evolution of the state of the various stages of the life cycle of a generation or of whole population.
Test of the model: To test the FRUFLY model in the field, the medfly activity period was studied by a Jackson trap baited with para-pheromone attractant, trimedlure (TML). The field trials were conducted in the Souss (South East of Morocco), at Ait Melloul station inside the institute of Agronomic and Veterinary Sciences of Hassan II (Agadir). This station is an area protected against public and foraging animal access; it was not treated with any pesticide and is close to a meteorological station. Traps were distributed in a randomized block design. Five blocks were established with four traps each. The field test was carried out from November 17, 1996 to January 12, 1997 and flies were removed from traps every week. During the field test, the TML was changed after each period (every 2 week). Traps were suspended 1 to 2 m above the ground in the south-east section of the tree in a clear space not touching the canopy.
| Table 1: | Duration (days) of the pupae development of medfly at four constant temperatures |
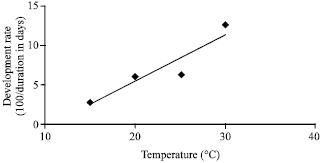 | |
| Fig. 1: | Degree-day model: Rate of development = 0.7T-7.8 (R2 = 0.99). The threshold for development Th = 11.2 is determined by extrapolating the line to the abscissa, i.e., to the temperature at which there is no development, while the thermal constant K = 143 is the reciprocal of the slope β, i.e., 1/β |
Traps were sampled twice a week and traps within each block were rotated sequentially after each sample the numbers of male medflies were recorded and the total number trapped per trap/block was summed over the 2 samples per week. Thus, one replicate consisted of the sum total catch per five blocks for each wk.
Insect development and degree-day model: The mean duration of pupal development (measured in day) at various constant temperatures with standard deviation (Duyck and Quilici, 2002) are given in Table 1. Since the number of individuals at 15°C was low, those of 35°C show no emergence. The other temperatures showed a linear regression of the development rate.
From the development rates at the four temperatures, a degree-day model was calculated (Fig. 1). A linear regression model between temperature and development rate was established over the range 15-30°C. A strong and positive linear relationship was observed between temperature and development rate (R2 = 0.99). Lower temperature threshold for pupal development was estimated as 11.2. The day degree (°D) requirement to complete the pupal stage was 143°D. of developmental rates at those fore temperatures yield a slope of 0.5 and an y-intercept of 4.8.
RESULTS
The field results: Para-pheromone (Trimedlure Plug) traps are designed to collect males medfly. Figure 2 shows the percentage mean number of captured weekly males medfly during the end of autumn and the beginning of winter.
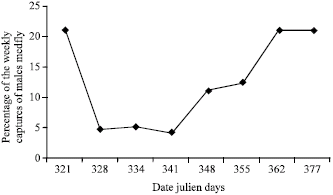 | |
| Fig. 2: | Density of the male medfly population in the field between 17 November 1996 and 12 January 1997 |
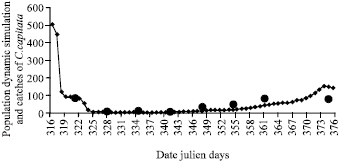 | |
| Fig. 3: | The flight activity of the Mediterranean fruit fly C. capitata (•) and the simulation results of degree-day model during 56 days from 12/11/1996 (♦) |
The caught of flies by parapheromone traps were between 17 November 1996 and 12 January 1997. the analysis of the climatic data of the station shows that the conditions most favorable to the activity of the fly are met during the first week then become unfavorable at the second and the third week and start to be restored during the two last weeks of the test.
Simulation and comparison with experimental results: The simulations carry out by FRUFLY model have reproduced the level of dynamic population behavior over the test period (Fig. 3). These results are in better agreement with experimental data results of the insect collected by para-pheromone traps.
Prediction simulation: From 12/11 to 31/3, with the area conditions, the simulation by FRUFLY, allowed to predict three successive generations which are overlapping (Fig. 4). The analysis of this figure makes it possible to determine the type of quantitative evolution of each generation. We notices for each generation evolution of a Gaussian type and consequently we can easily predict the period of the blossoming of the adults Starting from the first occurrence of the fly in the traps, the temperature sum of 240 degree-days is reached on 28/11. Around this date the second generation appeared.
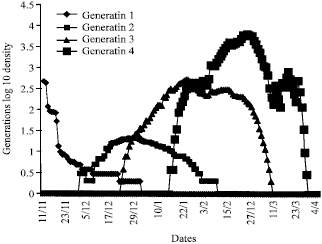 | |
| Fig. 4: | Representation of the various generations envisaged by simulation |
DISCUSSION
The degree-day model developed for the medfly seems to give a rather good estimate of the developmental time for the medfly and predictions of the occurrence of the next generations. According to the model, the second generation of medfly begins to fly in the beginning of December, which corresponds to the observed flight activity, as the peak of the catches (in the para-pheromone traps) was increased after the first week of December. But further experiments are still required to validate the model.
Problems can arise, however, in predicting development rates at times of the year when mean temperatures tend to be around the lower temperature threshold or above the optimum level for development. There are two reasons for this, first the temperature development rate relationship deviates most from linearity towards the temperatures extremes. Secondly, although at constant temperatures the range of temperatures at which development can proceed is limited by the upper and lower lethal thresholds, most immature life stages can survive and develop for short periods outside these thresholds. This problem is particularly significant when predicting ovarian maturation rates of females because adults can often survive for long periods when temperatures are above or below the maturation thresholds. In such situations, model and indeed any other model that relates maturation rates to temperature as a straightforward additive process, is inappropriate.
Since the pheromone traps used for validation only reflect male flight activity, while the model was developed from the mean developmental time of both sexes; this might be a source of error in the validation of the model. If the model had been developed using only male developmental times, the model might have predicted the peaks of flight activity occurring a little bit earlier.
Linear model have been used (Barclay, 1982; Delrio et al., 1986) relatively successfully to predict the start of adult emergence and the yearly phenology of multivoltine species and confirming their usefulness for pest management purposes.
CONCLUSION
The simulation results have provided the population dynamic that closely resemble the observation in the field. It has shown its potential for further research on insect population dynamics and its application in pest control as a support tool. The approach has the advantage of leading to precise predictions that can be quantitatively assessed. It has the disadvantage that unlike the quantitative models, certain simplifying assumptions must be invoked.
REFERENCES
- Duyck, P.F. and S. Quilici, 2002. Survival and development of different life stages of three Ceratitis sp. (Diptera: Tephritidae) reared at five constant temperatures. Bull. Entomol. Res., 92: 461-469.
Direct Link