
M. Daami- Remadi
National Institute of Agronomic Research of Tunisia,
PRRDA-CE 4042 Chott-Mariem, 4042, sousse, Tunisia
H. Jabnoun- Khiareddine
High School of Horticulture and Breeding of Chott-Mariem,
4042, Sousse, Tunisia
F. Ayed
High School of Horticulture and Breeding of Chott-Mariem,
4042, Sousse, Tunisia
M. El Mahjoub
High School of Horticulture and Breeding of Chott-Mariem,
4042, Sousse, Tunisia
Journal of Agronomy
Year: 2006 | Volume: 5 | Issue: 2 | Page No.: 350-355







ABSTRACT
Mycelial growth on PDA of Fusarium graminearum, F. oxysporum f. sp. tuberosi, F. sambucinum and F. solani, causal agents of potato tuber dry rot, depends upon temperatures ranging from 5 to 40°C. An interaction between pathogens tested and temperatures used for culture incubation was observed (at p≤0.05). These Fusarium species showed in vivo both thermal pics of aggressivity: a first one at low temperatures (10-15°C) and a second at higher temperatures (30-35°C). Nevertheless, at temperatures less than 25°C, F. sambucinum and F. graminearum were the most aggressive, while F. solani was the most aggressive at temperatures superior to 30°C.
PDF Abstract XML References
How to cite this article
M. Daami- Remadi, H. Jabnoun- Khiareddine, F. Ayed and M. El Mahjoub, 2006. Effect of Temperature on Aggressivity of Tunisian Fusarium species Causing Potato (Solanum tuberosum L.) Tuber Dry Rot. Journal of Agronomy, 5: 350-355.
DOI: 10.3923/ja.2006.350.355
URL: https://scialert.net/abstract/?doi=ja.2006.350.355
DOI: 10.3923/ja.2006.350.355
URL: https://scialert.net/abstract/?doi=ja.2006.350.355
INTRODUCTION
Potato tuber dry not incidence varies upon Fusarium spp. involved in disease development, soil, environmental conditions and cultivars. F. sambucinum, F. solani var coeruleum and F. oxysporum are the most aggressive agents (Tivoli et al., 1986; Theron and Holz, 1989, 1990; Desjardins et al., 1992; Choiseul, 1996; Carnegie et al., 2001). However, these pathogens are site specific and their aggressivity varies upon experimental conditions such as temperature used for tuber incubation (Jellis and Starling, 1983; Bjor, 1987; Theron and Holz, 1987).
Fusarium has a thermal spore formation and development optima at 20°C; but this temperature does not necessarily coincide with that required for tuber infection. Fusarium thermal infection optima are specific for every country showing pathogen adaptation aptitude (Tivoli and Jouan, 1981; Tivoli et al., 1986; Seppanen, 1981).
Thermal requirements of tuber infection depend also on Fusarium species. They varied from 24 to 30°C for F. avenaceum, from 18 to 24°C for F. coeruleum and from 12 to 30°C for F. sambucinum (Seppanen, 1982).
Temperature affects also the installation phase of different Fusarium species, the number of tuber infections as well as the degree of virulence of Fusarium species and the cultivar’s resistance (Henrikson, 1977; Tivoli and Jouan, 1981; Corsini and Pavek, 1982).
In Tunisia, dry rot incidence tends to increase from year to an other due to the importance of the soil phase of the causal agents, non respect of sufficiently long rotation, application of monoculture and absence of resistant cultivars and registered fungicides (Daami-Remadi and El Mahjoub, 1996a,b; 2004, 2006; Priou and El Mahjoub 1999; Triki et al., 2001). Furthermore, dry rot causal agents are frequently present in rotten tubers as mixed infections and these phenomena are observed in refrigerated and traditional storages. Losses due to dry rot development in store are reported to exceed 50% even in refrigerated stores (Chérif et al., 2000). However, fungal isolations showed variation in Fusarium spp. frequency between both types of storages without a qualitative variation in pathogen population (Daami-Remadi, unpublished data). Thus, in order to determine the specific thermal requirements of the Tunisian Fusarium species, representing the major causal agents of tuber dry rot in local potato storage conditions, a range of temperatures is tested in vitro and in vivo on the development of F. solani, F. graminearum, F . sambucinum and F. oxysporum f. sp. tuberosi. This study, which has never been investigated in Tunisia, will also explain the presence of mixed infections observed in refrigerated and/or traditional potato storages and the predominant species of Fusarium involved.
MATERIALS AND METHODS
Pathogens: Fusarium spp. were isolated from tubers showing typical symptoms of dry rot or potato plants partially or totally wilted. It is a fungi complex composed of F. solani, F. graminearum (isolated on 2003), F. sambucinum and F. oxysporum f. sp. tuberosi (isolated on 2004). These Fusarium species are cultured at 25°C on PDA (Potato Dextrose Agar) containing streptomycin-sulphate at 300 mg L-1.
For their long term preservation, monoconidial cultures are maintained at -20°C in a solution containing 20% of glycerol.
Fusarium spp. identification is realized via several morphological (phialides, microconidies, macroconidies, chlamydospores) and cultural criteria such as colony aspect, pigmentation, front growth following keys proposed by Messiaen and Cassini (1968) and Tivoli (1988).
These agents are inoculated to healthy tubers bimonthly and re-isolated and purified after dry rot development for maintaining their virulence.
Potato cultivars: Tubers used for pathogenicity tests of Fusarium spp. belong to cv. Spunta the most cultivated in Tunisia. They are obtained on 2004 from the Technical Potato Centre of Tunisia and maintained, in the dark, at 6°C for two months and moved to room temperature several hours before inoculation.
Isolation of Fusarium spp.: Potato tuber tissues showing dry rot symptoms were superficially disinfected with an hypochlorite-sodium solution diluted at 1% during 4 min, rinsed with sterile distilled water then air dried under a laminar flow. Disinfected fragments are placed on PDA (Potato Dextrose Agar) containing streptomycin-sulphate at 300 mg L-1. Purification of isolates is realized via successive cultures from front growth of colonies, followed by a monoconidial culture. Pathogen cultures are incubated at 25°C for 7 days.
Determination of thermal optimum for Fusarium spp. in vitro growth: The technique used consists of aseptically deposing a fragment of mycelium (diameter 6 mm), removed from a seven days pathogen culture, at the centre of a Petri dish (90 mm in diameter) containing PDA. Fungal cultures were incubated, in the dark, in growth chamber under different thermal conditions: 5, 10, 15, 20, 25, 30, 35 and 40°C during 6 days.
One isolate per Fusarium specie is used. Every elementary treatment is repeated eight times. At the end of incubation period, mean colony diameter is noted.
Statistical analyses (ANOVA) are performed following a completely randomised factorial design where Fusarium isolates and temperatures are both fixed factors. Means are separated using Fisher’s protected LSD test (p≤0.05).
Effect of temperature of incubation on Fusarium spp. aggressivity on potato tubers: Tubers of cv. Spunta are individually inoculated by four Fusarium species. Inoculation technique consists of deposing an agar disc (6 mm diameter) colonized by pathogen at wounds (6 mm diameter and depth) occasioned on tubers by a corer bore. Tuber incubation is realized in growth chamber at 5, 10, 15, 20, 25, 30, 35 and 40°C for 21 days at high relative humidity. Every elementary treatment is repeated twenty times (ten tubersxtwo wounds).
After incubation period, tubers were cut longitudinally via sites of inoculation. Parameters of dry rot induced (maximal width (w) and depth (d)) are noted. The pathogen penetration within tubers, is calculated following formula of Lapwood et al. (1984) where:
Statistical analyses (ANOVA) are performed following a completely randomised factorial design where fungal isolates and temperatures are both fixed factors. Means are separated using Fisher’s protected LSD test (p≤0.05).
RESULTS
Thermal optimum for mycelial growth of Fusarium spp.: Mean diameter of colonies formed after 6 days of culture on PDA depends on Fusarium species and temperatures of incubation. A significant interaction (at p≤0.05) was observed between both fixed factors.
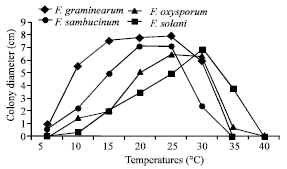 | |
| Fig. 1: | Effect of temperatures of incubation on mycelial growth of Fusarium spp. causing potato tuber dry rot (PDA, after 6 days) LSD (Fusarium spp.xTemperature) = 0.222 cm (at p≤0.05). |
At higher temperatures such as 40°C, none mycelial development was observed for all Fusarium species tested, whereas at lower temperatures such as 5°C, there is no mycelial growth for F. solani and F. oxysporum f. sp. tuberosi but a minimum growth is noted for F. sambucinum and at a lesser degree, for F. graminearum.
Thermal optima for mycelial growth of Fusarium spp. are specific for every specie. In fact, for F. sambucinum and F. graminearum, the optimum is comprised between 20 and 25°C and this last specie also grows at 15°C.
F. oxysporum f. sp. tuberosi seems to have its thermal optimum of in vitro growth between 25 and 30°C whereas that of F. solani is at 30°C. Figure 1 also shows that at temperatures inferior to 25°C, F. sambucinum and F. graminearum grow faster but at temperatures superior to 25-30°C, F. solani and F. oxysporum f. sp. tuberosi are most favoured.
Effect of temperature of incubation on dry rot development on tubers inoculated by Fusarium spp.: Development of tuber dry rot is assessed on cv. Spunta depending on Fusarium species and temperatures of incubation. Variance analyses of mean pathogen penetration into tubers revealed existence of a significant interaction (at p = 0.05) between both fixed factors. Figure 2 showed that aggressivity of different Fusarium species varies upon temperatures of incubation and their thermal optimum for dry rot induction seems to be specific. In fact, that of F. graminearum is comprised between 25 and 30°C, but this pathogen occasioned a lesser rot at lower temperatures such as 5 and 10°C. However, at 10-15°C, F. sambucinum seems to be more aggressive but at these temperatures, rots observed tend to develop in large then in depth (Fig. 3).
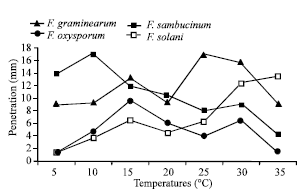 | |
| Fig. 2: | Effect of temperatures of incubation on dry rot development on potato tubers cv. Spunta inoculated by Fusarium spp. (after 21 days) LSD (Fusarium spp.xTemperatures) = 2.779 mm (at p≤0.05). |
It is important to note that majority of Fusarium species showed both thermal pics one at lower temperatures and a second at higher temperatures. In fact, F. sambucinum showed a pic of aggressivity at 10°C and a second but less important at 30°C. F. graminearum has a first pic at 15°C and a second at 25-30°C.
At temperatures inferior to 25°C, F. sambucinum and F. graminearum seem to be more aggressive than F. solani and F. oxysporum f. sp. tuberosi. Furthermore, for these both agents, none dry rot development was observed at 5°C, contrary to that noted for F. sambucinum and F. graminearum. For F. oxysporum f. sp. tuberosi, maximum of aggressivity was reached at 15°C whereas at higher temperatures such as 30 and 35°C, it was F. solani that occasioned the most important dry rot (Fig. 4).
When aggressivity of Fusarium spp. was observed depending on incubation temperatures, two groups can be distinguished, one containing F. sambucinum and F. graminearum that are more aggressive at temperatures inferior or equal to 20°C and the second group includes F. oxysporum f. sp. tuberosi and F. solani.
At a temperature such as 30°C, the difference between Fusarium spp. aggressivity is well distinguished. In fact, all Fusarium species occasioned dry rot which importance was determined by the aggressivity of the pathogens. F. graminearum and F. sambucinum are more aggressive than the other species. However at 35°C, F. graminearum showed decrease of their development in vivo and F. solani becomes more aggressive.
It is important to note that at 40°C, latent infections due to Erwinia spp. and interaction with other tuber rot agents inhibits assessment of dry rot development at this temperature.
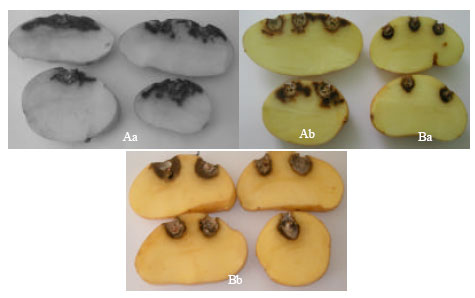 | |
| Fig. 3: | Dry rot occasioned by F. sambucinum on potato tubers cv. Spunta incubated for 21 days at 10°C (Aa) and 5°C (Ab) and by F. graminearum at 5°C (Ba) and 15°C (Bb) |
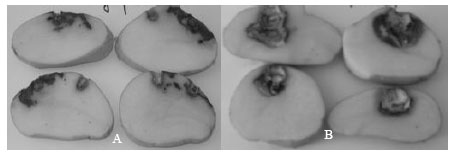 | |
| Fig. 4: | Dry rot occasioned by F. oxysporum f. sp. tuberosi on potato tubers cv. Spunta incubated for 21 days at 15°C (A) and by F. solani at 35°C (B) |
DISCUSSION
As temperature has a significant role on pathogen development and tuber susceptibility to dry rot, a range of temperatures of incubation was tested on mycelial growth and aggressivity of Fusarium complex involved in this disease. Obtained results showed that thermal optima for in vitro and in vivo growth are specific for each Fusarium specie as observed by Tivoli and Jouan (1981) who found that necrosis future is not the same for all Fusarium species.
Furthermore, this is the first study in Tunisia revealing interaction of the Fusarium complex with temperatures of incubation showing, consequently, variable mycelial growth and aggressivity. This result joins on the whole that of Theron and Holz (1991) reporting that Fusarium spp. incidence depends considerably upon temperatures of incubation of inoculated tubers.
F. sambucinum and F. graminearum, tested in the current study, are aggressive at lower temperatures; this result explains presence of mixed infections occasioned by F. sambucinum and F. graminearum in refrigerated storage in Tunisia and consequently their ability to cause dry rot at temperatures 5 and 10°C. These both Fusarium species are also able to induce disease even at higher temperatures, joining findings of Seppanen (1981) showing that F. sambucinum and F. culmorum have a double thermal optimum one at 10-12°C and a second at 24-27°C; this phenomenon was explained by optimised tuber defence reactions between 15 and 25°C.
Present study showed that optimum temperature for F. sambucinum in vivo development is reported to be 10°C however this pathogen induced rot of lesser importance under other thermal conditions whereas it was reported to be, respectively 10-15°C, 17°C, 15-20°C and even 25°C according to Seppanen (1982), Boyd (1972), Tivoli and Jouan (1981) and Theron and Holz (1990); this fact demonstrated that this pathogen has a large thermal spectrum suggesting presence of an important ability of adaptation. Furthermore, F. sambucinum isolated in France induces symptoms between 5 and 30°C (Tivoli and Jouan, 1981).
Aptitude of in vivo development of F. sambucinum at lower temperatures was also observed by several authors such as Corsini and Pavek (1986) who found that 10°C, is more favourable for this pathogen growth than for that of F. coeruleum and according to Liu and Kushalappa (2002), when infection occurs, it is difficult to slow down lesion expansion even at lower temperatures mainly for F. sambucinum. This is also the case for F. graminearum as Ali et al. (2005) recently reported that, in North Dakota (USA), this pathogen was detected on cultivars Shepody and Russet Burbank stored at lower temperatures and Secor (personal communication) detailed that induced rots occurred on tuber lots stored at 10°C. However, F. graminearum occasioning, in the current study, a dry rot significantly important at 15°C, comparatively to other species, seems to have its thermal optimum of aggressivity comprised between 25 and 30°C. This result joins findings of Tivoli and Jouan (1981) who classified French isolates of F. graminearum, with F. culmorum, within a group of higher thermal optimum situated between 25 and 30°C.
F. solani seems to have its thermal optimum of in vivo development situated between 30 and 35°C contrary to 17, 15-20, 15 and 35°C reported, respectively by Boyd (1972), Tivoli and Jouan (1981), Seppanen (1982) and Theron and Holz (1990). In the same way, Carnegie et al. (2001) found that incubation of tubers for several weeks at lower temperatures, followed by a higher temperature, is more favourable for dry rot development by F. solani var. coeruleum than their incubation at a constant temperature of 15°C. However, at 5°C, our data showed none dry rot induction on tubers inoculated by F. solani and F. oxysporum f. sp. tuberosi. This result joins that obtained by Theron and Holz (1990) showing that F. oxysporum, F. solani, F. equiseti and F. scripi are the lesser active pathogens at 5°C but at lower temperatures, they found that minor species such F. acuminatum and F. crookwellense become more pathogenic than F. oxysporum and F. solani (Theron and Holz, 1991). Hide and Cayley (1985) concluded, via lower temperature effect on in vivo Fusarium spp. development, that a chemical treatment is more efficient when tubers are stored at 5°C than at 15°C, suggesting that inoculum introduced via wounds infected tubers slowly, at lower temperatures, being more accessible to fungicide.
Following our present results, Tunisian Fusarium species causing potato tuber dry rot have their specific thermal optimum of aggressivity different from previous findings. Furthermore, the presence of a double thermal pics for major tested Fusarium species clarifies presence of mixed infections on rotten tubers and occurrence of F. graminearum and F. sambucinum in refrigerated stores. Finally, for the further studies and specially differential susceptibility of local potato cultivars and clones to tunisian Fusarium species, temperature effect should be implicated for the screening of tuber resistance to dry rot.
ACKNOWLEDGMENT
Authors thank High School of Horticulture and Breeding of Chott-Mariem (ESHE-CM), Technical Potato Centre of Tunisia (CTPT), Technical Centre of Biological Agriculture (CTAB) and Interprofessional Group of Vegetables (GIL) for their partially financial contribution.
Many thanks for Aymen Youssef for his excellent technical assistance.
REFERENCES
- Bjor, T., 1987. Laboratory screening for resistance to gangrene and to Fusarium dry rot in potato. Potato Res., 30: 517-524.
CrossRefDirect Link - Carnegie, S.F., A.M. Cameron and P. Haddon, 2001. The effect of date of haulm destruction and harvest on the development of dry rot caused by Fusarium solani var. coeruleum on potato tubers. Ann. Applied Biol., 139: 209-216.
Direct Link - Daami-Remadi, M. and M. El Mahjoub, 2006. Presence in Tunisia of Fusarium sambucinum isolates resistant to Benzimidazoles: In vitro growth and aggressiveness on potato tubers. Biotechnol. Agron. Soc. Environ., 10: 7-16.
Direct Link - Desjardins, A.E., H.W. Gardner and K.M. Weltring, 1992. Detoxification of sesquiterpene phytoalexins by Gibberella pulicaris (Fusarium sambucinum) and its importance for virulence on potato tubers. J. Ind. Microbiol., 9: 201-211.
CrossRefDirect Link - Jellis, G.J. and N.C. Starling, 1983. Resistance to powdery dry rot (Fusarium sulphureum) in potato tubers. Potato Res., 26: 295-301.
CrossRefDirect Link - Lapwood, D.H., P.J. Read and J. Spokes, 1984. Methods for assessing the susceptibility of potato tubers of different cultivars to rotting by Erwinia carotovora subsp. atroseptica and carotovora. Plant Pathol., 33: 13-20.
Direct Link - Lui, L.H. and A.C. Kushalappa, 2002. Response surface models to predict potato tuber infection by Fusarium sambucinum from duration of wetness and temperature and dry rot lesion expansion from storage time and temperature. Int. J. Food Microbiol., 76: 19-25.
CrossRefDirect Link - Priou, S. and M. El Mahjoub, 1999. Bacterial and fungal diseases in the major potato-growing areas of Tunisia. EPPO Bull., 29: 167-171.
CrossRefDirect Link - Theron, D.J. and G. Holz, 1989. Fusarium species associated with dry and stem-end rot of potatoes in South Africa. Phytophylactica, 21: 175-181.
Direct Link - Theron, D.J. and G. Holz, 1990. Effect of temperature on dry rot development of potato tubers inoculated with different Fusarium spp. Potato Res., 33: 109-117.
CrossRefDirect Link - Theron, D.J. and G. Holz, 1991. Prediction of potato dry rot based on the presence of Fusarium in soil adhering to tubers at harvest. Plant Dis., 75: 126-130.
Direct Link - Triki, M.A., S. Priou and M. El Mahjoub, 2001. Effects of soil solarization on soil-borne populations of Pythium aphanidermatum and Fusarium solani and on the potato crop in Tunisia. Potato Res., 44: 271-279.
Direct Link