
H. Pirdashti
Faculty of Agriculture, University of Tarbiat Modarres,
P.O. Box 14155-4838, Tehran, Iran
Faculty of Agriculture, University of Mazandaran, Sari, Iran
Z. Tahmasebi Sarvestani
Faculty of Agriculture, University of Tarbiat Modarres,
P.O. Box 14155-4838, Tehran, Iran
GH. Nematzadeh
Faculty of Agriculture, University of Mazandaran, Sari, Iran
A. Ismail
Crop, Soil and Water Science Division, International Rice Research Institute, DAPO P.O. Box 7777, Metro Manila, Philippines
Journal of Agronomy
Year: 2003 | Volume: 2 | Issue: 4 | Page No.: 217-222







ABSTRACT
An experiment was conducted to compare fifteen lowland rice (Oryza sativa L.) genotypes for drought tolerance based on some physiological parameters in germination stage. Five levels of water stress (0, -3.0, -5.0, -7.5 and B10 bars) treatments were given with polyethylene glycol (PEG 6000). The water solutions were replaced with fresh solutions regularly. Seed germination percentage and seedling growth decreased with increasing water stress in all the genotypes. Among the genotypes, Tarom, Khazar, Fajr and Nemat recorded better germination and seedling growth in terms of radicle and plumule length under stress conditions than other genotypes.
PDF Abstract XML References
How to cite this article
H. Pirdashti, Z. Tahmasebi Sarvestani, GH. Nematzadeh and A. Ismail, 2003. Effect of Water Stress on Seed Germination and Seedling Growth of Rice
(Oryza sativa L.) Genotypes. Journal of Agronomy, 2: 217-222.
DOI: 10.3923/ja.2003.217.222
URL: https://scialert.net/abstract/?doi=ja.2003.217.222
DOI: 10.3923/ja.2003.217.222
URL: https://scialert.net/abstract/?doi=ja.2003.217.222
INTRODUCTION
Rice is the most important crop at the global level, as it used as a staple food in the most countries of the world (Dowling et al., 1998). In Iran, according to the area, rice has third place among cereals that is after wheat and barley. On the other hands, rice is the second main consumed in Iran, with wheat being the main staple for most of the population (Kaveh, 1997). One of the main problems of rice cultivation and production is the lack of water resources, especially during periods of low rainfall. Irrigation dominates water use in Iran and surface water storage has been increased by construction of numerous multi-purpose dams and reservoirs along rivers flowing from the Zagros and Elburz Mountains. Iran major rice producing region within Gilan and Mazandaran provinces (which produce 80 to 85% of the country’s rice) is located between the Elburz Mountain and the Caspian Sea. Drought, a period of no rainfall or irrigation that affects plant growth, is a major constraint for about 50% of the world production area of rice (Oryza sativa L.) (Mitcell et al., 1998). Iran’s rice production has been dawn for the 1999-2001 years, as a shortage of irrigation water reduced planted area. In the past year drought affected 20 of 28 provinces in Iran, with the southeastern, central and eastern parts of the county being most severely affected (Reynolds, 2001). Thus, identifying stress-resistance mechanisms is essential for genetic improvement of stress resistance in crop plants. The use of suitable methodology for measuring stress resistance in large breeding populations is also very important. Some genotypes are more drought resistance than others, out-yielding those exposed to the same degree of water stress (Mitcell et al., 1998). Germinating seeds in solutions of different water potentials is a convenient method for establishing the response of germination to low water potentials (Naylor, 1992). The germination of seeds consists of a number of phased events. The processes of imbibition, radicle emergence, plumule emergence, radicle growth and plumule growth, each respond to the availability of water and each has a critical water potential at which the process ceases (Naylor, 1985). The first phase of water uptake by the seeds involves movement of water into the free space (apoplast) and dose not depends on the osmotic potential of the surrounding solution. The second slower linear phase of water uptake involves the movement of water across cell membranes into the cells of the seeds and is determined by the difference between the osmotic potential of the seed and that of the medium (Simon, 1984; Bewley and Chattopadhyay, 1994). Clearly the solute chosen should not exert a phytotoxic effect; for this reason NaC1 is little used. Sugars, e.g. mannitol, may be absorbed and utilized by plants, resulting in nutritional effects. The most widely used solute is the polymer, polyethylene glycol (PEG) and this has been used with monocots, dicots, gymnosperms, fungi and yeasts. Polyethylene glycole is available as a range of products with different avarage molecular weight (Naylor, 1992). Polyethylene glycol (PEG) compounds have been used to simulate water stress effects in plants (Murillo- Amador et al., 2002). There is evidence that as water potential decreases (i.e. as water becomes less availabele), the first plumule emergence and growth are reduced, then radicle emergence and growth and finally imbibition mat be prevented. In addition, the critical values may differ between crop species, between cultivars of a single species and also between seed lots of different quality in a single cultivar (Naylor, 1992). The decrease in both rate and percentage of germination and seedling growth as a result of the decrease in osmotic potentials in seed environment has been shown by several authors (Manabendra et al., 1998; Dighe and Rajurkar, 1981; Jha and Singh, 1997; Babu et al., 1986; Reddy and Vajaranabhiah, 1993; Choi et al., 2000). This paper includes the effect of polyethylene glycol on the germination and seedling growth of different rice genotypes to find out how different genotypes (which cultivated now or recently introduced to Iran) behave under water stress conditions and screening of them for pot and field experiments.
MATERIALS AND METHODS
This study was conducted at the Rice Research Institute of Iran-Deputy of Mazandaran (Amol) located in north of Iran (52° 22` E; 36° 28` N). Germination and seedling growth of different rice genotypes were studied using different osmotic potentials. The experiment was conducted in a factorial arranged in a completely randomized design with four replications of 40 seed per replicate. The first factor (genotype) had 15 levels (Mahali Tarom, Hashemi Tarom, Nemat, Neda, Fajr, Sahel, Khazar, Sephidroud, Dasht, 7165 Line, 7602 Line, 7604 Line, 77-44 Line, IR 24 Line and IR 56 Line). These genotypes represent a wide range of germplasm from traditional to improved lowland types. The second factor (osmotic potential) had five levels (0, -0.3, -0.5, -7.5 and –1.0 MPa). Distilled water was used as a control (0 MPa) and osmotic potentials (-0.3, -0.5, -7.5 and –1.0 MPa) were prepared adding polyethylene glycol 6000 (Sigma Chemicals) to distilled water according to Michel and Kaufmann’s equation (Michel and Kaufmann, 1973). Seeds of each genotype were previously disinfected by immersion in a sodium hypochlorite solution, containing 10% active chlorine, for 1 min. The seeds were then washed three times with sterilized distilled water. Germination tests were carried out in sterilized Petri dishes [(150 * 15 mm) covered at the bottom with two sheets of whatman No. 1 filter paper] that had been autoclaved. Each dish was moistened with 10 ml distilled water or uniform amounts of desired osmotic solutions (-0.3, -0.5, -7.5 and –1.0 MPa). This experiment was carried out in a dark growth chamber (Grouc, controlled environment, Tehran, Iran) at 25±0.5°C and 80%±1 of relative humidity. The number of germinated seeds (the final germination percentage) was determined after seven days. Not all-germinable seeds necessarily produce normal seedlings the germinated percentage is expressed on the basis of normal seedlings only (Agrawal and Dadlani, 1992). Seeds were considered germinated when the radicle was at least 2-mm long. Ten seedling were chosen randomly and seedling growth was measured by dry and fresh weights of different parts (radicle and plumule) of the seedling on the seventh day. Dry weight was determined after drying each organ in a forced-air dryer at 70°C for 48 h. the lenghts of radicle (hypocotyle) and plumule (epicotyle) were measured with a ruler. Data for final germination percentage after arcsine transformation was analyzed. All statistical tests were carried out using the Statistical Analysis System (SAS Institute, 1996).
RESULTS AND DISCUSSION
Germination
The ANOVA for germination percentage showed that all factors and interaction of factors were significant at P=0.01. The germination percentage decreased with the decrease in osmotic potential in PEG solutions so that germination stopped completely in –1.0 MPa (Table 1).
| Table 1: | Germination and seedling growth traits as affected by different osmotic potentials |
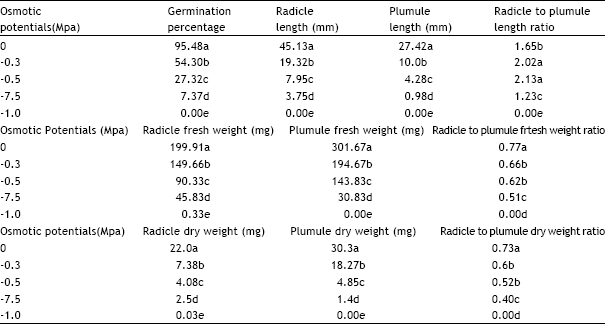 | |
| In a column, means followed by a common letter are not significantly different at the 5% level by Duncan’s Multiple Range Test | |
| Table 2: | Comparison of germination and seedling growth traits among different rice genotypes |
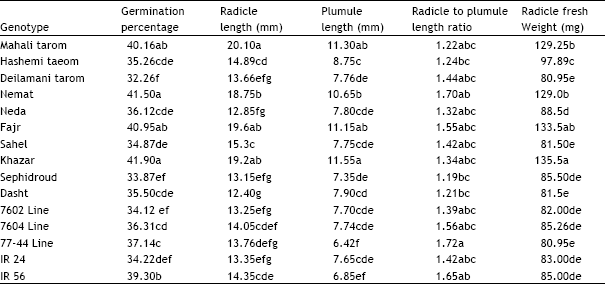 | |
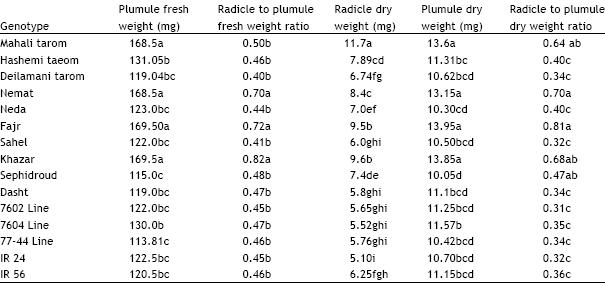 | |
| In a column, means followed by a common letter are not significantly different at the 5% level by Duncan’s Multiple Range Test | |
Results indicated that the highest amount of germination percentage was recorded for the Khazar, Nemat, Mahali Tarom and Fajr genotypes (Table 2). Babu et al. (1986), Manabendra et al. (1998, 2000) and Jha and Singh (1997) reported similar findings for the upland genotypes. It seems that lowering the osmotic potential with PEG decrease water availability for seeds and then caused low germination. The physical process of water uptake leads to activation of metabolic process, as the dormancy of the seed is broken following hydration. Elevated drought stress slows dawn water uptake by seeds, thereby inhibiting their germination and root elongation. At the low osmotic potential, PEG inhibited the process of inhibition, germination and root elongation of genotypes (Murillo-Amador et al., 2002; Simon, 1984).
Seedling growth
The analysis of variance showed significant difference between the two main factors, genotype and osmotic potential and interactions for the all variables except for radicle to plumule length ratio, which did not show any differences between genotypes. Differences among genotypes showed that radicle and plumule length of Tarom, Fajr and Khazar were higher than other genotypes (Table 2). Under water stress, it has been shown that the inhibition of radicle emergence is mainly because of a decrease in water potential gradient between the external environment and the seed (Murillo- Amador et al., 2002). The radicle and plumule length and their fresh and dry weight decreased with the decrease in osmotic potential in PEG (Table 1). Among the genotypes, Mahali Tarom showed higher radicle dry weight in all the treatments (Table 2). The high correlation coefficient (r= 0.96**) between radicle length and dry weight indicate that lowering the osmotic potentials with PEG be caused a decrease in the weight of radicle in addition to its length. On other hands, coefficient of variation for plumule length (15.39) was greater than radicle length (12.96) that indicate plumule growth is more sensitive than radicle growth to water stress. The reasons, as described by Matsuo et al. (1995), may be that radicle is the first organ emerged from the seed, therefore its growth is faster than plumule growth and also, plumule have not any direct contact with water resource because of its lately emergence and its location on the seed (Matsuo et al., 1995).
Results of this paper showed that (a) although these genotypes (except IR 24 and IR 56) cultivated in the same geographic area, but display distinct responses to drought stress. In this sense, genetic variability within a species offers a valuable tool for studying mechanisms of drought tolerance. One of these mechanisms depends on the capacity for osmotic adjustment, which allows growth to continue under drought conditions. Under water stress, osmotic adjustment is achieved by synthesis and accumulation of organic compatible solutes (Murillo- Amador et al., 2002; Babu et al., 1996). However, in this study these ions and organic solutes were not measured; (b) This work is useful for plant breeders by proposing early tests for screening new or existing genotypes for their tolerance to drought stress and © Mahali Tarom, Khazar, Fajr and Nemat appear to be more tolerant to drought stress than other genotypes at germination and early seedling growth stage.
ACKNOWLEDGMENTS
The assistance of Mr. Heshmatollah Pirdashti and Mr. M. Rahimi in the conduct of the experiment is gratefully acknowledged. Financial support by the Tarbiat Modarres University and the Rice Research Institute of Iran- Deputy of Mazandaran (Amol) was greatly appreciated.
REFERENCES
- Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Murillo-Amador, B., R. Lopez-Aguilar, C. Kaya, J. Larrinaga-Mayoral and A. Flores-Hernandez, 2002. Comparative effects of NaCl and polyethylene glycol on germination, emergence and seedling growth of cowpea. J. Agron. Crop Sci., 188: 235-247.
CrossRefDirect Link