
A. Prabhu Britto
Center for Medical Electronics, Department of Electronics and Communication Engineering,
Dr. G. Ravindran
Faculty of Information and Communication Engineering, Anna University, Chennai 600 025, India
Information Technology Journal
Year: 2006 | Volume: 5 | Issue: 6 | Page No.: 1122-1126







ABSTRACT
The realm of medical imaging has always been fascinating because of its capability to yield visual clinical information about the human body. It was found that genetic information carried in the chromosomes also carries information about certain clinical abnormalities or diseases. Medical imaging and cytogenetics, when working in conjunction give us a valuable diagnostic avenue. This study is expected to help both the layman and the professional by providing them a good insight into this fascinating interdisciplinary area. Further, this study will help to understand the promising role of medical imaging in the field of human cytogenetics and disease diagnosis.
PDF Abstract XML References Citation
How to cite this article
A. Prabhu Britto and Dr. G. Ravindran, 2006. Medical Imaging-A Promising Technology in the field of Human Cytogenetics and Disease Diagnosis. Information Technology Journal, 5: 1122-1126.
DOI: 10.3923/itj.2006.1122.1126
URL: https://scialert.net/abstract/?doi=itj.2006.1122.1126
DOI: 10.3923/itj.2006.1122.1126
URL: https://scialert.net/abstract/?doi=itj.2006.1122.1126
INTRODUCTION
Medical imaging and its associated technologies have been rmdergoing a rapid and revolutionary phase of development in the past few decades. Scientists and physicians can gather potentially life-saving information by peering non-invasively into the human body enabled by recent technological breakthroughs in the reahn of medical imaging.
Medical imaging has evolved into a tool for surgical planning and simulation, intra-operative navigation, radiotherapy planning, accurate diagnosis and also to investigate susceptibility to various diseases. Modem imaging devices provide exceptional views of internal anatomy, yet computers are not used independently to quantify and analyze the embedded structures in the human body without physician intervention. Accurate and repeatable quantitative data should be efficiently extracted in order to support the spectnun of biomedical investigations and clinical activities.
A variety of image processing techniques are available for processing medical images. But some of these techniques used to obtain significant useful information from biomedical images are limited by the sheer size of the datasets and the complexity and variability of the anatomic shapes of interest. Also, the shortcomings typical of sampled data such as sampling artifacts, spatial aliasing and noise may cause difficulties in the processing of medical images.
CHROMOSOMES
A chromosome is a very long continuous piece of DNA, which contains many genes, regulatory elements and other intervening nucleotide sequences. Chromosomes are present in every nucleated cell of all living organisms and carry the genetic information that is used as a blueprint in reproduction of cells and organisms. The chromosomes are responsible for the determination and transmission of hereditary characteristics. The name Chromosome is derived from Greek chroma for color and soma for body, because they can be stained with certain dyes.
In 1865, Gregor Mendel traced inheritance patterns of certain traits in pea plants and showed that they obeyed simple statistical rules. From his statistical analysis, Mendel defined a concept described as an allele, which was the frmdamental mrit of heredity. The significance of Mendel's work was re-discovered after his death by other scientists working on similar problems.
It is now known that genetic information is normally carried on a nucleic acid called DNA (deoxyribonucleic acid), which is capable of carrying genetic instructions for the biological development of all cellular forms of life. During reproduction, DNA is replicated and transmitted to offspring. DNA is also referred to as the molecule of heredity, as it is inherited and used to propagate traits.
In bacteria and other simple cell organisms, DNA is not separated by a nuclear envelope from the cytoplasm. In higher organisms, chromosomes are formd in the nucleus of eukaryotic cells.
DNA, histone and nonhistone proteins are the components of chromatin of which these bodies are made. Hwnan being has 23 pairs of chromosomes, 22 pairs being autosomes for body structure and function and a pair of sex chromosomes which determine whether the child is male or female and other characteristics (Trask, 2002; Therman and Susman, 1993; Tjio and Levan, 1956).
Each chromosome is made up of two chromatids held together by the centromere, which separates the chromosome into upper and lower arms. The shorter upper arm is called p and the longer lower arm is referred to as q. The location of the centromere varies depending on the number of the chromosome which is associated to the chromosome in accordance with ISCN (1978, 1981, 1985).
CHROMOSO:ME STRUCTURE AND CONDENSATION
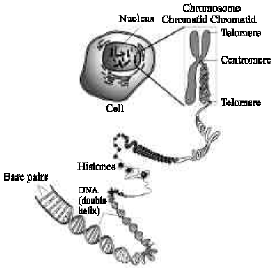
Watson and Crick (1953) discovered the double helix DNA geometrical structure (Fig. 1). Measured along the helicoids axis the length of the DNA molecules in one hwnan cell is between 1.5 and 2m. This enormous length is packed into the cell by coiling (the 2 nm-wide) helix twice; once into a fiber or 11 nm thick, which in turn is coiled into a chromatin fiber of 36 nm thick, visible only by electron microscopy.
In the chromosomes of eukaryotes (including hwnan beings) the uncondensed DNA exists in a quasi-ordered structure inside the nucleus, where it wraps around structural proteins called histones. During metaphase of mitosis the chromosomes are condensed, which is the only natural context in which individual chromosomes are visible with an optical microscope.
 | |
| Fig. 1: | Chromosome |
(Courtesy:http://www.accessexcellence.org/RC/VL/GG/chromosome.htrnl)
 | |
| Fig. 2: | Different levels of DNA condensation (1) Single DNA strand. (2) Chromatin strand (DNA with histones). (3) Chromatin during interphase with centromere. (4) Condensed chromatin during prophase. (Two copies of the DNA molecule are now present) (5) Chromosome during metaphase. |
Courtesy:http:I/en.wikipedia.org/wiki/Image:Chro matin_chromosome.png
{kind=link}
In the latter stages of prophase (Fig. 2) and early stages of metaphase, the chromatin strands become more and more condensed and appear as separate entities and move toward the equatorial plate to get attached to the spindle fibers. They cease to function as accessible genetic material and become a compact form. It is at this time that the chromosomes are most condensed and the least active. At this stage, the cells are harvested for cytogenetic studies.
A typical chromosome has one centromere, with arms projecting from the centromere. The ends of the chromosomes are special structures called telomeres. The condensed chromatin strands called chromatids, become visible as a chromosome, linked at the centromere. Long microtubules are attached at the centromere and two opposite ends of the cell. During mitosis, the microtubules pull the chromatids apart, so that each daughter cell inherits one set of chromatids. Once the cells have divided, the chromatids are uncoiled and can function again as chromatin. In spite of their appearance, chromosomes are highly structured.
Metaphase chromosomes are those that are frequently studied with optical microscopy. In metaphase chromosomes the chromatin fiber in turn is folded onto an acidic protein scaffold with coils of0.6 J.liD wide that form the chromatids, the smallest threads visible in optical microscopy. The total length of the chromatids is dependent on the preparation technique, but in general it is in the order of 300 Jlm for metaphase chromosomes, so the coiling and folding accounts for a length contraction by a factor of 5000. As a result, the smallest detail-if assumed 0.3 J.liD large-will be in the order of one thousandth of the total amount of material, i.e., in the order of six million base pairs Vossepoel, 1987).
CYTOGENETICS
Cytogenetics is the study of the structure and the frmction of chromosomes. It is also useful in the study of human chromosomes in health and disease. Chromosome specimen from the living cells of an organism can be examined rmder a light microscope and the images can be enhanced with stains and fluorescing probes. Chromosomes are basically identified by size, centromere location and the visible banding pattern. Defects in chromosomes can lead to abnormalities in the structure or frmction of that particular organism.
Chromosome studies are an important laboratory diagnostic procedure in prenatal diagnosis; in patients with mental retardation and multiple birth defects; in patients with abnormal sexual development and in some cases of infertility or multiple miscarriages. Cytogenetic analysis is also useful in the study and treatment of patients with malignancies and hematologic disorders.
The cytogeneticist does not actually look at the molecular level of the chromosome. By observing the banding and centromere locations, the cytogeneticist identifies chromosomes, ascertains the appropriate number and type of chromosomes; detects abnormalities m banding such as deletions, mverswns and translocations.
The field of human Cytogenetics was launched by Tjio and Levan (1956). A suspension of human mitotic cells (Hsu. 1979). when added to a serendipitous addition of water (before the cells were fixed and dropped onto glass microscope slides) caused the chromosomes to spread apart from each other so that Tjio and Levan (1956) could accurately cmmt the full complement of 46 human chromosomes. Ford and Hamerton (1956) confirmed the number 46 by independent investigations.
Some of the 22 homologous pairs and two sex chromosomes that make up the normal human complement of 46 chromosomes can be identified based on their sizes and position of the Centromere, while others can only be classified into groups, as they are similar (Graselli, 1969). These findings set off many discoveries that associated specific chromosomal abnormalities with disease in the late 1950s and quickly established the central role of Cytogenetics in medicine.
The erstwhile prevailing dogma had held the chromosome cormt at 48 for more than 30 years till the geneticist Painter (1923) had reported his observations of testicular cells. In the ensuing years, technological advances that have combined innovations in molecular biology, chemistry and instnunentation have transformed human cytogenetics.
Fleming and Arnold first observed human chromosomes in the 1880s (Trask. 2002). Only three years after Watson and Crick (1953) elucidated the structure of the DNA helix was the chromosome number established as 46. Vogel and Motulsky (1997) ascribe this delay to both technological and politico-social causes. However, soon after the number 46 was firmly established, scientists readily applied the new cytogenetic technique to the investigation of phenotype-genotype correlations in humans and began to tap useful information from naturally occming chromosomal rearrangements.
In human chromosomes, it is known that the short arm is designated p (petite) and the long arm q (the next letter in the alphabet). Chromosomes are usually characterized by one of the two parameters. The arm ratio (qlp) is the length of the long arm divided by that of the short ann. The centromere index expresses the length of the short ann as a percentage of the total chromosome length [(p/p+q) x 1 00]. The length of a particular chromosome relative to others in the same metaphase cell, together with the ann ratio or centromere index, is sometimes sufficient to permit identification of the chromosome. Equally important is the length of the chromosome; this shows wide variation, the largest being about five times as long as the shortest.
At the Paris Conference (1972), a system was proposed for identifying human chromosome bands and indicating various chromosome abnormalities. Telomeres, centromeres and a number of prominent bands are used as landmarks. A section of a chromosome between two landmarks is called a region and these regions are numbered 1, 2, 3 and so on in both directions, starting from the centromere. The bands within the regions are numbered according to the same rule. Thus, the first band in the second region of the short arm of chromosome 1 is 1 p21 (ISCN. 1978).
Yunis (1980) has presented a corresponding system for prophase bands. For instance, 14q32 indicates chromosome 14, long ann 3, band 2. For a sub-band, a decimal point is placed after this followed by the number of the sub-band (they are numbered sequentially from the centromere). The last sub-band in chromosome 14 is thus 14q32.3. When the sub-band is fwi.her sub-divided, an additional digit is used, the last sub-sub-band thus being 14q32.33. An extra or missing chromosome is denoted with a plus or a minus sign respectively, which is indicated before the number of the chromosome. Thus, the chromosome constitution of a female with trisomy for chromosome 13 would be 47,XX,+13 and a male with monosomy for 21 would have the formula 45,XY,-21.
A plus sign after the symbol of a chromosome arms means that a segment is added to it, while a minus sign indicates that a segment is missing from it. For example, a female with the cri du chat syndrome caused by a deletion in the short ann of chromosome 5, would have the syndrome formula 46,XX,5p- and a male with an abnormally long 4q would be designated as 46. XY. 4q+. The karyotype of a female with a Robertsonian translocation (centric fusion) between chromosomes 13 and 14would be 45.XX.t(l3ql4q). The formula for a male carrier in whom chromosome arms 3p and 6q have exchanged segments. the breakpoints being 3pl2 and 6q34. would be 46.XY.t(3;6)(pl2;q34).
MEDICAL IMAGING IN lllJMAN CYTOGENETICS
Wang et al.(2005) point out that since the 1980s. a large number of research groups armmd the world have been working on developing computer-assisted chromosome detection and classification systems based on digitally acquired microscopic images. Initial tests based on the limited datasets demonstrated promising results in algoritlnn and system development. Despite the progress made over the last two decades, there are several limitations of the current computer-assisted chromosome classification systems.
First, these systems depend heavily on slide preparation, image enhancement process and the optical system that captures the images. The performance of the systems can be improved when the slides are well-prepared, the microscope has good optical quality and the camera can digitize the image with sufficient clarity and resolution.
Second, the performance of these systems is affected by the results of chromosome segmentation. Early studies fmmd that a number of automated classification systems were somewhat successful in karyotyping the chromosomes rmder favorable imaging conditions. The typical case error rate was approximately 20% (Wu et oZ .,1989). If the chromosomes were touching. overlapping or deformed as sho\Vll in the majority of Images acquired in the clinical laboratories, the classification error rate was substantially increased (Pantaleao et oZ.,2002).
Third, although there are several publicly available databases of chromosome Images (i.e., Copenhagen dataset, discussed in Delshadpour. 2003; Sweeney and Musavi, 1993), current systems have not been tested and evaluated using a large and diverse independent dataset. Many of the tests were performed using simulated data (Delshadpour. 2003) or a round-robin (leave-one-out) validation method (Sweeney and Husavi, 1993). Hence the robustness of these systems IS largely unknown.
Fourth, although several commercialized software and systems have been developed, they are mostly semi-automatic products. To identify metaphase spreads, the scanning speed and accuracy of current commercial systems do not match the clinical service demand. Once metaphase spreads are visually identified, current computerized systems can help to partially pair the rmtouched chromosomes. The interaction of a skilled laboratory technician is required to check the results and manually complete the karyotyping (including correcting the mismatch of chromosome pairs).
Recent studies by Hoffbrand et ol. (2000) have found that consistent chromosomal changes led to isolation of the genes involved in the pathogenesis of leukemia and other cancers. Detection of these consistent, recurrent chromosomal changes has allowed the division of patients into clinical groups which define their duration of remission and mean survival time (LeBeau and Rowley, 1984). Hence, using the chromosome imaging technique to detect chromosomal disorder provides a much more sensitive approach than other medical imaging techniques (i.e., x-ray radiographs and computer tomography) not only in detecting cancers or other genetic disorders at an early stage but also in monitoring the efficacy of patient treatment. Without a computer-assisted and reliable semi- or fully-automated detection and classification system, visually searching for and detecting these subtle chromosomal changes (disorders) is a labor-intensive and time-consuming process because of difficulties involved in the culture of primary tumor cells, low mitotic indices and poor chromosomal morphology. Moreover, only a small proportion of total acquired testing cells (metaphase spreads) are associated with cancers or tumors in each patient. The subjective evaluation of subtle chromosomal changes can also lead to errors and substantial inter or intra-reader variations.
Wang et al. (2005) emphasize that further research is needed to develop and evaluate computer assisted systems for detection and classification of banded chromosomes in order to facilitate diagnosis of cancers (or other genetic diseases) and reduce diagnostic errors.
CONCLUSIONS
This study has elucidated the promising role of the medical imaging in the realm of human cytogenetic studies. It is formd that the area of Automated Human Cytogenetic Studies is relatively recent. Still, there has a lot of contributory research work done, as rmderstood from the various publications that have arisen in this area. Yet the observations of Wang et al. (2005) help us to rmderstand that still more significant research has to be carried out. Hence, it is concluded that the area of automated human cytogenetics needs further significant research exhaustive research output that will further facilitate diagnosis of genetic diseases and cancers, thus contributing to better health for mankind.
REFERENCES
- Delshadpour, S., 2003. Reduced size multi-layer perceptron neural network for human chromosome classification. Proc. 25th Annual Int. Conf. IEEE Eng. Med. Biol. Soc., 3: 2249-2252.
CrossRefDirect Link - Pantaleao, C.H.Z., F.M. Azevedo, E.T. Pereira, M.C.M. Ribeiro and J.L.B. Marques, 2002. Development of a computerized system for cytogenetic analysis and classification. Proceedings of the 2nd Joint 24th Annual Conference and the Annual Fall Meeting of the Biomedical Engineering Society, October 23-26, 2002, IEEE Xplore, pp: 2211-2212.
CrossRef - International Congress of Human Genetics, 1971. Paris Conference (1971): Standardization in Human Cytogenetics. Birth Defects Original Article Series Vol. 8, The National Foundation, New York.
Direct Link - Trask, B.J., 2002. Human cytogenetics: 46 chromosomes, 46 years and counting. Nat. Rev. Genet., 3: 769-778.
Direct Link - Wang, X., B. Zheng, M. Wood, S. Li, W. Chen and H. Liu, 2005. Development and evaluation of automated systems for detection and classification of banded chromosomes: Current status and future perspectives. J. Phys. D: Applied. Phys., 38: 2536-2542.
Direct Link