
Carly Ballinger-Boone
Department of Biology, University of Kentucky, Kentucky, United State of America
Ogechi Anyagaligbo
Department of Biology, University of Kentucky, Kentucky, United State of America
Jate Bernard
University of Virginia, Charlottesville, Virginia, United State of America
Sonya M. Bierbower
Department of Biology, William Paterson University, Wayne, New Jersey, United State of America
Esther E. Dupont-Versteegden
Division of Physical Therapy, Department Rehabilitation Sciences, College of Health Sciences, University of Kentucky, Kentucky, United State of America
Adam Ghoweri
Department of Pharmacology and Nutritional Sciences, University of Kentucky, Kentucky, United State of America
Abigail Greenhalgh
Department of Biology, University of Kentucky, Kentucky, United State of America
Doug Harrison
Department of Biology, University of Kentucky, Kentucky, United State of America
Oscar Istas
Department of Biology, University of Kentucky, Kentucky, United State of America
Micaiah McNabb
Department of Biology, University of Kentucky, Kentucky, United State of America
Christa Saelinger
Department of Biology, University of Kentucky, Kentucky, United State of America
Alex Stanback
Department of Biology, University of Kentucky, Kentucky, United State of America
Maddie Stanback
Department of Biology, University of Kentucky, Kentucky, United State of America
Olivier Thibault
Department of Pharmacology and Nutritional Sciences, University of Kentucky, Kentucky, United State of America
Robin L. Cooper
Department of Biology, University of Kentucky, Kentucky, United State of America
LiveDNA: 1.523
International Journal of Zoological Research
Year: 2020 | Volume: 16 | Issue: 1 | Page No.: 33-62







ABSTRACT
Gram-negative bacteria produce endotoxins in the form of lipopolysaccharides (LPS). The direct effect of LPS itself has not been well studied. Peptidoglycan recognition proteins (PGRPs) are potential receptors for LPS in Drosophila and likely other arthropods whereas in mammals the Toll receptors mediate a response by LPS. In larval Drosophila the myogenic heartbeat is dampened by LPS. The LPS exposure at larval Drosophila glutamatergic neuromuscular junctions (NMJ) depresses the amplitude of evoked and spontaneous release, potentially blocking glutamate receptors on the muscle. In contrast, LPS increases evoked response but not quantal events at the crayfish glutamatergic NMJ. In both mentioned invertebrate NMJs, the muscle transiently hyperpolarizes to exposure of LPS and effects on evoked release are reversible with short exposures to LPS. No effects of LPS on primary sensory neurons of crab or crayfish occur. Evoked responses at the cholinergic frog NMJs were depressed but spontaneous quantal events were not. No hyperpolarization of the muscle was observed for frog or rat skeletal muscle from exposure to LPS. The hippocampus of the rodent displayed a rapid depression in evoked responses but is reversible with short exposures. This was also mimicked for sensory-CNS-motor nerve evoked responses in crayfish and larval Drosophila. The RNAi expression for PGRP-LC and PGRP-LE in Drosophila did not alter the responses to LPS. The same dosage of LPS (500 μg mL–1) from Serratia marcescens was used in all animal models. Thus, the effects of LPS on synaptic function and skeletal/cardiac muscle is species and receptor subtype specific but not transmitter subtype specific.
PDF Abstract XML References Citation
How to cite this article
Carly Ballinger-Boone, Ogechi Anyagaligbo, Jate Bernard, Sonya M. Bierbower, Esther E. Dupont-Versteegden, Adam Ghoweri, Abigail Greenhalgh, Doug Harrison, Oscar Istas, Micaiah McNabb, Christa Saelinger, Alex Stanback, Maddie Stanback, Olivier Thibault and Robin L. Cooper, 2020. Effects of Bacterial Endotoxin (LPS) on Cardiac and Synaptic Function in Various Animal Models: Larval Drosophila, Crayfish, Crab and Rodent. International Journal of Zoological Research, 16: 33-62.
DOI: 10.3923/ijzr.2020.33.62
URL: https://scialert.net/abstract/?doi=ijzr.2020.33.62
DOI: 10.3923/ijzr.2020.33.62
URL: https://scialert.net/abstract/?doi=ijzr.2020.33.62
INTRODUCTION
Bacteria infect both animals and plants alike; thus, organisms have developed various mechanisms to protect themselves from infection and combat bacteria once beyond the initial protective barriers or the first line of defence of the body. The gram-negative bacterial strains, such as Escherichia coli, Pseudomonas aeruginosa and Serratia marcescens are linked to septicemia (i.e., blood or hemolymph borne infections) in animals from invertebrates to humans1-5. One common mode of infection is consumption of plants and animals, which can introduce various types of bacteria into a diet; thus, due to the widespread presence of bacteria in nature, various strains can be amplified and passed through the food chain6. Different forms of bacteria have different effects on a host’s immune system. Gram-negative bacteria produce and release endotoxins in the form of lipopolysaccharides (LPS)7. The different forms of LPS elicit varying immune responses from a host. In mammals, the induced immune responses results in raising levels of circulating cytokines. The higher levels of cytokines amplify the immune response and generate abnormal neural and cardiac function that are almost always harmful to the host8-11.
The LPS-induced response of sepsis is well established and results in degradation of skeletal muscles of mammals12. This is mediated by the release of cytokines into the systemic circulation, as well as activation of cellular responses within muscle exposed to endotoxins. Understanding these cellular responses could potentially lead to treatments of pathological aliments associated with systemic infection. Blocking pathways, such as for NF-κB expression, in cultured muscle, as well as muscles in rodents decreases the induced atrophy13. Activation of NF-κB and TNF-α induced by sepsis also inhibits myogenic differentiation, which could impact muscle growth and repair14. In addition to skeletal muscle being impacted by sepsis, cardiac function can be severely impaired, with effects ranging from muscle atrophy to electrical dysfunction in mammals15-20.
Not only do the induced cytokines act on tissues, but LPS itself can act directly on cells independently of the secondary immune induced responses. The induction of the immune response from gram-negative bacteria is caused by LPS, as well as other associated peptidoglycans21. Considering that commercially obtained LPS is a mixture of LPS and associated peptidoglycans, the effects of this mixture parallels the effects induced on a tissue or in whole animal responses that are exposed to gram-negative bacteria. Thus, it is essential to understand the action of LPS and the associated peptidoglycans released by gram-negative bacteria on cellular function.
The LPS binds to a Toll-like receptor 4 (TLR4) known as the CD14/TLR4/MD2 receptor complex in mammals22,23. These receptors are conserved from insects to mammals24. Despite Toll receptors being discovered in Drosophila melanogaster, the immune response does not appear to be mediated by these receptors, but instead by the Immune deficiency (Imd) signaling pathway25-27. The LPS and the peptidoglycan layer in gram-negative bacteria trigger these Imd receptors, yet the expression levels, locations and density of these receptors in insects have not been fully identified. Likewise, the physiological actions of activating these receptors on tissues (heart, body wall muscles and nervous system) have not been comprehensively investigated. The LPS has recently been examined for direct effects on the larval heart of Drosophila melanogaster and medicinal blow fly (Phaenicia sericata) in a dose-dependent manner28. However, responses varied between these two species of insects to the same type of LPS29.
Recent studies have examined the acute effects of LPS on cardiac function in insects and crayfish, as well as on sensory-CNS-motor neuron responses in crustaceans and Drosophila. Studies on the direct focus of LPS have targeted the evoked synaptic transmission and spontaneous quantal responses at the neuromuscular junctions of amphibian, crayfish and larval Drosophila with opposite effects. Thus, if one model preparation is emphasized for the effects of LPS on synaptic transmission at the NMJs, an incorrect generalization of responses may occur.
This comparative review highlights not only the effects on synaptic transmission at NMJs, but also the odd occurrence of hyperpolarizing the membrane potential on the skeletal muscles of some animals (i.e., invertebrates) but not others (i.e., rodent). In addition, the effects on central circuits of invertebrate and vertebrate models are examined and discussed. Since the IMD receptors are assumed to be the targeted mediators of LPS in Drosophila, preliminary investigations embarked on using RNAi transgenic lines to address the mechanism of action for the direct effects on physiological functions. The novel results of these studies are presented to help in addressing future studies. Lastly, this review discusses potential future topics of interest to advance this field of study.
AMPHIBIAN NMJ: A CHOLINERGIC JUNCTION
The NMJs of the frog skeletal muscle have been used as a model for understanding synaptic transmission30. An additional benefit is that the synaptic properties mimic mammalian skeletal NMJs in pharmacological and electrophysiological properties31-34, although there are synaptic structural differences35-37. The neurotransmission utilizes acetylcholine (Ach) as the transmitter and nicotinic receptors on muscle. The commonly used frog preparations (Rana ‘genus’, Lithobates pipiens: Xenopus laevis) are viable at a variety of temperatures since the frogs are ectothermic and live in temperate environments, making them easy for laboratory experimentation. In addition, a very basic physiological saline maintains the synaptic preparation and muscle function for up to 8 h.
Direct application of LPS from E. coli and Salmonella typhimurium has been investigated for altering synaptic function by Person38-40 on frog NMJs (species not identified). LPS of different strains of E. coli (10 mg mL–1) causes different effects on the frequency of spontaneous quantal events, also known as miniature excitatory junction potentials (mEJPs). The effects are dependent of the presence of extracellular Ca2+. Two of the three E. coli strains cause an increase in the frequency of the mEJPs. Over time the frequency is severely reduced38. The LPS from Salmonella typhimurium (10 and 50 mg mL–1) increases the frequency of mEJPs and is followed by a reduction in the frequency of mEJPs by 45% within an hour. The amplitude of the evoked EJPs is rapidly reduced with exposure to LPS, but the mEJPs are still present. Over an hour, the frequency of the mEJPs did decrease substantially. Thus, LPS from Salmonella typhimurium did not block the Ach receptors. No effect of the resting membrane potential was reported39,40. Given the evoked EJPs decrease gradually in amplitude, it is unlikely there is a failure in blocking the induction of action potentials in the motor nerve. Person39,40 proposed that LPS is sticky and cannot be washed off the preparation since he could not get recovery of the evoked EJPs by exchanging the saline. Person39,40therefore, suggested that the voltage-gated Ca2+ channels may be compromised by LPS in the motor neurons of the frog, but at the same time there maybe be a leak of Ca2+ across the membrane resulting in an increase in the frequency of the mEJPs.
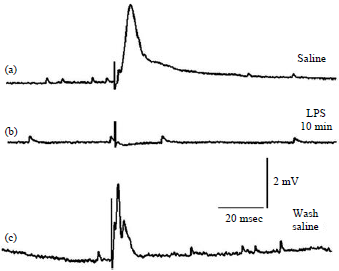 | |
| Fig. 1(a-c): | Intracellular recording of evoked excitatory junction potentials (EJPs) and spontaneous miniature EJPs (mEJPs) in the cutaneous pectoris muscle before, during and after exposure to 500 μg mL–1 lipopolysaccharides (LPS) endotoxin, (a) Evoked EJP and mEJPs are readily observed. The stimulus artifact precedes the EJP, (b) Evoked EJP does not occur with exposure to LPS after 10 min, however, the mEJPs are still present at a low frequency and (c) After exchanging the saline and removing LPS the evoked EJPs partially return and mEJPs are still present |
| Source: Reproduced from McNabb et al.42 | |
Repeating these studies with Northern leopard frogs (Lithobates pipiens) and using the cutaneous pectoris muscle41 with LPS from Serratia marcescens (500 μg mL–1), the evoked EJP amplitude is reduced rapidly to not being measurable42. However, even after 10 min of incubation in LPS, the responses return after exchanging the bathing media with a saline not containing LPS (Fig. 1). Thus, it is unlikely this strain of LPS is sticky. In addition, the frequency of the mEJPs is not altered over the 10 min of LPS exposure42. Perhaps a longer incubation time with LPS would have resulted in a decrease in the frequency in the mEJPs, however, one must also account for rundown in the frequency in the occurrence with time in control preparations not exposed to LPS. Thus, McNabb et al.42 propose that LPS may directly block voltage-gated Ca2+ channels on the presynaptic motor nerve terminal. The evoked responses can recover with short exposure to LPS. The high concentration of 500 μg mL–1 does not result in a leaky presynaptic membrane for Ca2+, as no substantial alteration on the frequency of the mEJPs was observed within the 10 min exposure.
CRAYFISH NMJ: A GLUTAMATERGIC JUNCTION
Crustacean NMJs are still relevant as model synaptic preparations to address structure-function and physiological properties of synapses. The availability of freshwater crayfish in most locations and ease in maintaining the isolated in situ synaptic preparations at room temperature with minimal salts to maintain physiological viable preparations are likely the reasons for the early use of crayfish for teaching physiology and conducting experiments43-46. The opener muscle in the walking leg of the crayfish has been a focus for many years due to the graded nature in EJPs and inhibitory junction potentials (IJPs) within the muscle. Many discoveries occurred using this preparation, such as presynaptic inhibition and demonstrating short-and long-term facilitation synaptic facilitation47-52. In fact, the neural innervation of the muscle was described very early in identified NMJs as compared to other preparations53. As with the frog NMJ, synaptic transmission on the opener NMJ was recognized to be quantal in nature54,55. An interesting aspect of the opener muscle is that it is innervated by a single excitatory motor neuron; however, there are regional differences in muscle fiber type and synaptic properties56-60. Since the evoked EJPs on the opener muscle are not spiking, as they do not generate action potentials on the muscle, the graded nature of synaptic transmission can be examined without having to reduce the Ca2+ or raising the Mg2+ levels in the saline to remain below a threshold of the action potential as for frog preparations50.
As for the effect of endotoxins on synaptic physiology using a crayfish NMJ, only one report has been identified61. In this report the muscle used was a deep abdominal extensor which is a high-output phasic like NMJ and is innervated by multiple excitatory motor nerve terminals62. In this study, LPS from Serratia marcescens (2 μg mL–1) was exposed to the NMJ. In contrast to the frog NMJ, the amplitude of the EJPs increase within 3 min and continued to increase for up to 10 min, at which time the preparation was rinsed with fresh saline. The amplitude of the EJPs decrease back to the initial amplitude prior to exposure to LPS. In addition, the mEJPs increased in frequency from 0.21-0.73 over 75 sec of exposure to LPS. The mEJPs were obtained from the claw closer muscle. It appears that the reported effects of LPS are from a single preparation in the abdomen and a single closer muscle preparation in the claw as no sample size or statistical analysis is reported in this study61. By the shape of the EJP in the trace shown in Parnas et al.61, it appears a second EJP is riding on the increased amplitude of the initial EJP after the exposure to LPS. Thus, one cannot be sure that an additional excitatory motor neuron was recruited or if a delay in vesicular recruitment occurred, promoting latency jitter of the evoked response of the initial motor neurons recruited.
In addressing the effects of LPS on the amplitude of evoked EJPs and the frequency of mEJPs with a preparation innervated by a single excitatory motor neuron, the well-defined opener NMJ of the walking was utilized in a report by Saelinger et al.41. Exposure to LPS of Serratia marcescens (500 μg mL–1) increases the amplitude of the evoked EJPs on average by 100% (p<0.05, Paired t-test, n = 12) within 10 min of exposure. However, not all the preparations demonstrate an increase in amplitude. Oddly, there is no significant increase in facilitation of the evoked EJPs within a train of 25 EJPs delivered at 40 Hz (Fig. 2a, b). In addition, no significant change in the frequency of mEJPs or amplitude of the mEJPs occur overall. Only one preparation of six showed an increase in the amplitude of the mEJPs. Thus, for the opener muscle LPS increases the evoked EJP amplitude without increasing synaptic facilitation or having any effect on the frequency or amplitude of the mEJPs within 10 min of exposure to LPS (Fig. 2c). It would appear LPS promotes Ca2+ entry with evoked transmission but not enough to result in a facilitation or an increase in spontaneous quantal events.
Saelinger et al.41 reported a novel finding for crustacean muscles regarding the effects of LPS exposure on the resting membrane potential (Fig. 3). Upon exposure to LPS, the membrane potential rapidly hyperpolarizes and slowly depolarizes back to the initial value over the 10 min of exposure. Thus, the hyperpolarization of the muscle increases the driving gradient for the evoked EJP and is likely partially responsible for the increased amplitude of the EJPs. This is not necessarily due to solely increased vesicular fusion events, but more likely there is a combination of effects. The large variation in the amplitudes of the mEJPs before and during LPS exposure may have masked slight increases in the driving gradient for producing larger quantal events. Flushing away the LPS recovers the resting membrane potential and this the driving gradient. This could also be a reason for the amplitude of the evoked EJPs to return to their initial values.
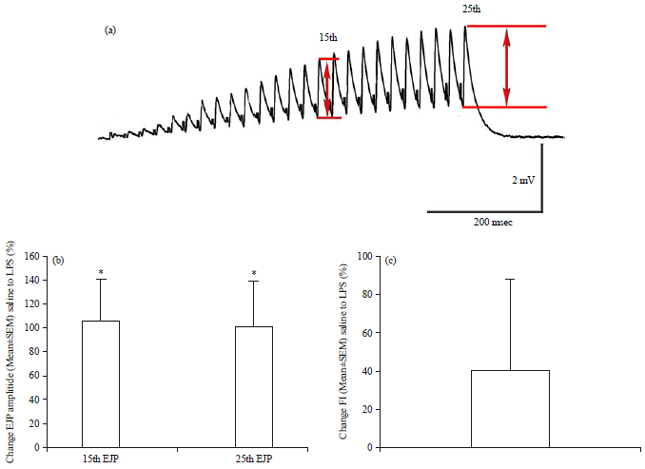 | |
| Fig. 2(a-c): | Effect of LPS exposure on the amplitude of the excitatory junction potentials (EJPs) measured in the opener muscle of the crayfish first walking leg, (a) Stimulation at 40 Hz of the opener excitatory motor nerve induces an EJP train of responses which facilitate, (b) Amplitudes of the 15th and 25th EJPs within a train increase with exposure to LPS (500 μg mL–1) and (c) LPS exposure did not significantly further enhance facilitation |
Average percent increase in the 15th and 25th EJP amplitudes from saline to LPS exposure are shown (p<0.05, paired t-test), No significant effect on the facilitation index were noted by exposure to LPS, source: Reproduced from Saelinger et al.41 | |
 | |
| Fig. 3: | Membrane potential and spontaneous quantal responses measured in the opener muscle of the crayfish before and during LPS exposure |
A representative intracellular muscle recording depicting the effects of switching the bathing media to one containing 500 μg mL–1 LPS, Membrane shows a transit hyperpolarization, Miniature EJPs (mEJPs) or quantal responses are present before and during LPS exposure, note the enlarge section of the trace to illustrate the quantal responses and varied sizes measured, three quantal responses are shown, source: Reproduced from Saelinger et al.41
DROSOPHILA NMJ: A GLUTAMATERGIC JUNCTION
The relative recent surge in use of the Drosophila NMJ for investigating synaptic transmission has come about with development of physiological salines and learning how to expose and record from the NMJs in larvae and adults63-68. The powerful amenable genetic attributes of Drosophila have increased the drive to utilize this preparation69-71. Interestingly, the pharmacology of the glutamatergic NMJ is similar to that of the crayfish and the general anatomical properties of low and high-output synapses (number of active zones and proximity to each other) are similar to those of the crayfish72-76. The larval NMJ is readily accessible for measures in synaptic transmission and like the crayfish opener NMJ, the muscle is non-spiking in normal physiological conditions. Thus, quantal analysis of synaptic transmission in measures of evoked EJPs and mEJPs is obtained with intracellular recording or loose patch recordings over defined regions of the nerve terminals to relate to synaptic function with wild type and strains containing mutational synaptic relevant proteins or ones with reductions or over expression of normally functional proteins60,66,77,78.
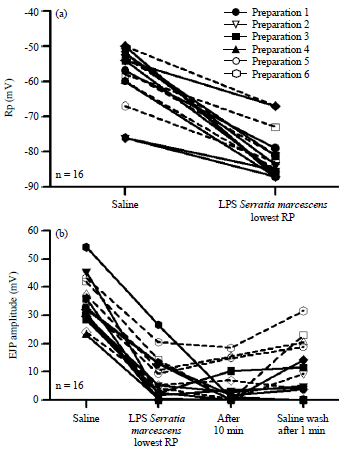
The exposure of LPS to the larval Drosophila was first reported by Cooper et al.79. Even though there are many similarities in synaptic properties to the crayfish NMJ the synaptic responses to LPS are in stark contrast. Exposure to LPS from Serratia marcescens (500 μg mL–1) rapidly (within 1-2 sec) decreases the amplitude of evoked EJPs and mEJPs (Fig. 4). Upon removal of the LPS, within 10 min of exposure, the responses recover. Like for the crayfish muscle, the larval muscle rapidly hyperpolarizes and slowly regain the potential over 10 min of LPS exposure. The membrane potential recovers quickly with removal of the LPS. The rapid hyperpolarization can be as much as an additional 20 mV from resting membrane potential of-60 mV (Fig. 5a). Thus, there is an increase in the ionic driving gradient for the producing larger amplitude evoked EJPs and mEJPs, but the rapid yet graded decrease in evoked EJPs and mEJPs would imply that the glutamate receptors are being blocked by LPS while the muscle is hyperpolarizing (Fig. 5b). It is not likely the presynaptic voltage-gated calcium channels are being blocked, as the evoked and mEJPs gradually decrease in amplitude. As for the explaining the mechanism for the hyperpolarization there is no definitive answer except possibly the Na-K pump or by a yet unknown mechanism. The cell membrane potential slowly recovers by use of pumps and exchangers to the initial state in the presence of LPS.
In addressing potential mechanisms, such as an induction of a calcium-activated potassium conductance, a Ca2+ free saline was used in the presence of LPS and the same response occurred. Considering that the known TEA sensitive channel in larval muscle might be activated, a 20 mM TEA incubation and then exposure to LPS with TEA was provided.
 | |
| Fig. 4(a-c): | Representative trace for exposure to LPS from Serratia marcescens at 500 μg mL–1 and enlarged segments to highlight the changes in amplitudes of the EJPs, (a) Overview of the evoked amplitude of the EJPs and the muscle membrane potential before, during and after removal of LPS, (b) Initial EJP amplitudes and the spontaneous mEJPs are readily observed and measurable and (c) During the exposure to LPS the amplitude of both the EJPs and mEJPs decrease and the mEJPs became so small they are lost in the background noise and (d) After removal of LPS the mEJPs and EJPs gradually reappear upon removing the LPS |
| Source: Reproduced from Cooper et al.79 | |
Again, no difference in the response to LPS was noted. Lastly, given LPS can induce nitric oxide synthase (NOS) and induce hyperpolarization in the membrane potential of rodent cardiac muscle, L-NAME incubation, to inhibit NOS at 1 mM along with TEA (20 mM) for 20 min prior to treatment with saline containing L-NAME+TEA+LPS (500 μg mL–1) was used, but the response to LPS remain the same79.
 | |
| Fig. 5(a-b): | Representative effects on resting membrane potential and EJP amplitude to exposure of LPS from Serratia marcescens at 500 μg mL–1, (a) Resting membrane potential of the body wall muscle hyperpolarizes with LPS exposure and (b) LPS depresses the EJP amplitudes rapidly during the hyperpolarization and is maintained depressed even after 10 min of exposure |
During the saline wash, removing LPS, the EJP amplitude starts to increase, p<0.001, paired t-test to the initial depressed EJP amplitude, Source: Reproduced from Cooper et al.79 | |
If a chloride channel were to be opened in the muscle, the membrane would depolarize as the equilibrium potential for Cl– in larval Drosophila body wall muscles is more depolarized than the resting membrane potential80,81. Whatever the mechanism is to account for the hyperpolarization of the body wall muscle in larval Drosophila it may also be a similar mechanism to account for the same response in the crayfish muscles described above as the responses are just as rapid and the slowly recovering of the potential during the exposure of LPS is similar. Oddly, though the glutamate receptors are similar pharmacologically in the crayfish and larval Drosophila NMJ, the same concentration and batch of LPS from the supplier (Sigma-Aldrich) appears to block the receptors at the Drosophila NMJ but not the crayfish NMJ.
RODENT SKELETAL MUSCLE-EFFECT OF LPS ON MEMBRANE POTENTIAL
Considering the crayfish and Drosophila muscle had a pronounced hyperpolarization but the frog skeletal muscle did not upon exposure to LPS, it was important to examine what would happen upon exposure of LPS to the membrane potential of rat skeletal muscles. Since these have not been previously reported, the methodology in the experimentation needs to be presented.
Methods and results: All animal procedures were conducted in accordance with guidelines for the care and use of laboratory animals as approved by the Institutional Animal Care and Use Committee of the University of Kentucky. Rats were housed in a temperature and humidity-controlled room and maintained on a 14:10 h light-dark cycle with food and water ad libitum. Brown Norway/Fischer 344 rats (10 months/450-550 g) were used. The extensor digitorum longus were rapidly removed after the animals were euthanized and maintained in physiological saline (Lily’s solution82). The membrane potential was measured with sharp glass microelectrodes (30 megaOhm resistance) filled with 3 M potassium acetate. A 1% agar bridge with the physiological saline was constructed using a micropipette plastic tip. This prevented DC changes on the ground wire located within the agar when changing the bathing media. The membrane potentials were obtained within the mid-region (i.e., endplate region) in the length of the fibers before and during exposure to LPS from Serratia marcescens (500 μg mL–1). The membrane potentials did not show any significant differences before and during LPS exposure (p>0.3, n = 6, paired t-test).
The values for the resting membrane potentials, before exposure to LPS (Mean±SEM, 80.2±2.4 mV, n = 6), obtained are similar to those reported previously (-76.6±0.4 mV) for recording under the endplate of soleus muscle from male Wistar rats (180-230 g)83. Thus, it appears the rodent and frog skeletal muscle do not undergo hyperpolarization, unlike the body muscles of crayfish and larval Drosophila, when exposed to LPS from Serratia marcescens (500 μg mL–1) within 5 min of exposure.
RODENT CNS-SLICE PREPARATION OF THE HIPPOCAMPUS
Previous literature has demonstrated the role of LPS as a potent pro-inflammogen with the ability to induce neuroinflammation and recruit proinflammatory cytokines like interleukin-1 (IL-1) and tumor necrosis factor alpha (TNF-α)84-88. A high density of IL-1 receptors has been observed in the hippocampus, where activation inhibits long-term potentiation (LTP) and reduces associated calcium influx89,90. These authors, as well as others, have proposed a mechanism whereby reduction in calcium flux through ryanodine receptors91, NMDA and voltage-gated receptors92 may mediate the impact of LPS under chronic cerebroventricular exposure. Other mechanisms proposed to mediate the negative impact of LPS on hippocampal function have unmasked the role of astrocytic calcium oscillations93 and microglial activation94-98. While neuroinflammation has been recognized as a key promoter of neurodegeneration, almost all previous work has been conducted using chronic exposures to neurotoxins in vivo. Thus, it was important to characterize the impact of acute LPS exposure on rodent central neurons using the hippocampal slice preparation. The same LPS was utilized (at concentrations used in the experiments presented throughout this review) to determine the effects of LPS on synaptic activation using field recordings.
MATERIALS AND METHODS
Slice preparation and sharp electrode electrophysiology: Eight adult (10 months) male and female wildtype mice (C57BL6.5XFAD-/- and C57BL6.ALDH2-/-) were used in this study. Brains were removed and incubated in ice cold low calcium (0.1 mM), high magnesium (8 mM) artificial cerebral spinal fluid (ACSF). The 350 μm thick coronal slices from the dorsal hippocampus were sectioned and incubated in normal calcium ACSF of the following composition (in mM): 114 NaCl, 3 KCl, 10 glucose, 1.25 KH2PO4, 26 NaHCO3, 2 CaCl2 and 2 MgCl2. Incubation was accomplished at 32°C in an interface-type chamber providing humidified oxygenation (95% O2/5% CO2) for at least 2 h. Each slice was placed in a recording chamber (RC-22, Warner Instrument) capable of maintaining the tissue under static or flow (perfusion) conditions. A bipolar stimulating electrode and an SD9 stimulator (Grass) were used to stimulate the Schaffer collaterals and a recording electrode was placed in Stratum pyramidale to record population spikes. The recording electrode was filled with 1 M potassium methyl sulfate (KMeSO4) and 10 mM HEPES. Signals were amplified and digitized (~5-10 KHz), AxoClamp-2BAxonTM; Digidata® 1550B. Data was processed and analyzed through Clampex V10.7.
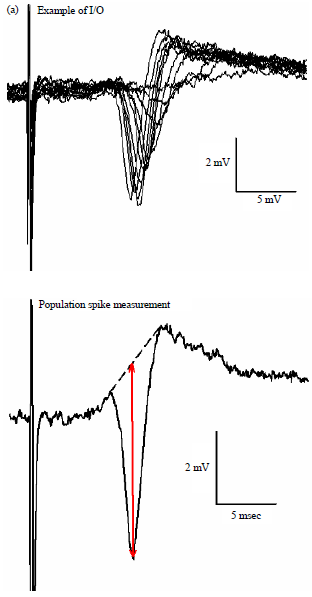 | |
| Fig. 6(a-b): | Example of population spikes and measurement, (a) Increases in stimulation intensity give rise to larger population spikes and (b) Amplitude of the population spike for each trace was measured from the middle of the top diagonal dashed line (top of EPSP) to the apex of the peak |
A total of 15 hippocampal slices were recorded and used to conduct these experiments. The n’s reported reflect the number of slices recorded under each condition. Two slices were removed from the analysis (based on>2SD) due to hyperexcitability.
Synaptic excitability and I/O measures: To determine the synaptic excitability of each slice, synaptic activation was elicited with increasing voltage (I/O) ranging from 5-36 V (every 10 sec). This protocol produces a reliable increase in the EPSP and the population spike amplitude as more axons are recruited (Fig. 6a).
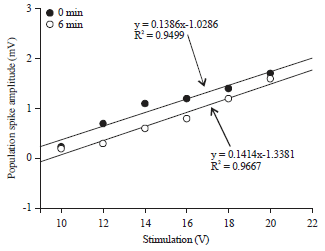 | |
| Fig. 7: | Comparison of population spike amplitude over time (before drug exposure) |
| I/O of a representative voltage recording from a slice is shown at 0 and 6 min later, slope and goodness of fit (R2) values suggest no difference with time | |
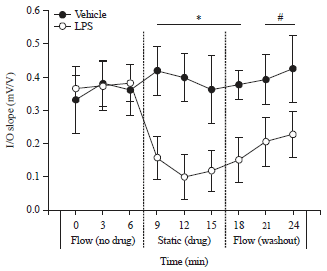 | |
| Fig. 8: | Effect of vehicle and LPS on synaptic activation |
| Means of I/Oslopes across both groups (vehicle, n = 6, LPS, n = 7) were compared every three minutes over 3 recording conditions (flow, static and flow). Synaptic activation was significantly attenuated in the LPS group. Impact of LPS on synaptic communication was relatively reversible in the time frame studied Mean±SEM are shown, *Significant differences between the control and LPS groups at p<0.001, pound signs indicate trends at p = 0.0536 | |
The I/O curves were repeated once every 3 min for 24 min (9 measurements). All data were acquired under 3 different conditions: (1) During ACSF perfusion (1 mL min–1) with no drug added, (2) with either LPS or vehicle administration under static condition and (3) During ACSF washout. The LPS (Serratia marcescens) was made daily in fresh oxygenated ACSF at 50 mg mL–1 and diluted 100X in the recording chamber to yield a final concentration of 500 μg mL–1. The amplitude of each population spike was measured (Fig. 6b) for each voltage and the slope of the resulting I/O from the different voltages used was extracted using a linear fit (Fig. 7). Only the linear portion of the I/O was fit to derive the R2 and the slope of the relationship for each slice. To investigate the stability of our recordings across time, given that each experiment lasted approximately 30 min, I/O slopes were measured of the population spike at time 0 and 6 min later (Fig. 7). All values of R2 were greater than 0.9 and no difference in the slopes were noted.
Results: I/O slopes determined across ~25 min of recording under perfusion and static conditions were averaged for each treatment group (Fig. 8). A strong and rapid synaptic depression (~65-75%) occurred upon LPS administration and persisted throughout the duration of the LPS exposure [F(1,125) = 19.820, p<0.001, asterisk Fig. 8]. Neither time, nor flow conditions altered the I/O slope across groups. Washout experiments provide evidence for small recovery of the synaptic communication (indicated as a trend in Fig. 8).
Discussion: In summary, LPS depressed the evoked field potentials within 9 min of exposure and a trend for recovery upon removal of the LPS was seen. This partial recovery is similar to that seen at the NMJs of the crayfish, Drosophila and frog models (chapters 2-4). The mechanisms underlying the synaptic depression seen at the hippocampal synapse may well depend on a glial involvement (i.e., astrocytes, oligo) including the release of cytokines (TNF-α and IL-1) through activated nuclear factor kappa B (NF-κB)99-102.
Unfortunately, few, if any, studies have investigated the impact of acute LPS on synaptic excitability and communication in the hippocampus. Using synapse-associated protein levels to infer on synaptic function (e.g., SYN and PSD-95), several groups have found that LPS could reduce SYN and PSD-95, perhaps highlighting the phenomenon reported here. This effect could be rescued by pretreatment with the cholinergic drug galantamine103. Similar findings were also shown when the anti-inflammatory/anti-oxidative drug juglanin was administered in a mouse model of parkinsonian-like symptoms induced by LPS and an increase in synaptic markers SYP, PSD-95 and SNAP-25 was observed104. Further, the anti-inflammatory drugs anthocyanins and ferulic acid appear capable of upregulating pre-and post-synaptic proteins that were initially reduced by LPS exposure105,106. It remains to be determined if the acute reduction in synaptic communication reported here relied on alterations in similar synaptic proteins.
It should be noted, however, that other reports have indicated opposing actions of TNF-α in models of neuronal injury associated with cell death. In these studies, TNF-α appears as a potent up-regulator of AMPA receptor trafficking, perhaps initiating excitotoxicity following injury88,107-108. Similar inflammation-mediated mechanisms have been proposed to interact with glial and neuronal excitotoxicity in models of clinical depression109. Whether these proposed mechanisms are direct effects of TNF-α on neurons or glia, or are dependent on both cell types is still unknown. However, it seems unlikely that LPS may have had a direct effect on neurons independently from glial cells in our experiments, as both cell types express various associated inflammatory cytokine receptors108. In fact, TLR4 receptors are located on microglia, astrocytes and oligodendrocytes, as well as neurons110-114. It should be noted that direct effects on neurons, independent from those on glial cells, have been reported in hippocampal neurons in culture115. In this work, the authors showed that LPS (1 μg mL–1, Escherichia coli) increases neuronal cytosolic [Ca2+] and promote cell death in older cultures without alterations in surrounding glial cells. These responses were only partially reduced by blocking NMDA receptors with high concentrations of MK801, suggesting NMDA receptors may not be critical to LPS-mediated cell death.
In conclusion, while it cannot be directly addressed the mechanism of the synaptic depression presented here, the acute and semi-reversible effect of LPS appears to be independent of excitotoxicity and cell death. The results associated with acute exposure of the hippocampal slice to LPS could help elucidate new signaling pathways and highlight worthwhile techniques for understanding the mechanism underlying clinical depression.
DROSOPHILA AND CRAYFISH CNS-SENSORY TO CNS TO MOTOR NEURONS
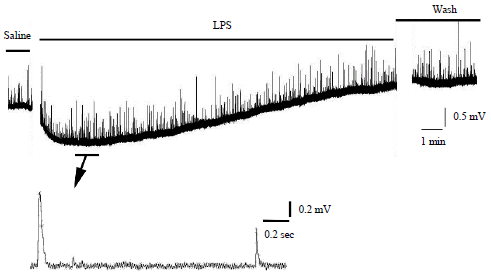
In addressing the sluggish sensory responses in larval Drosophila consuming food tainted with LPS, an evoked sensory-CNS-motor neuron circuit was examined before and during exposure of the circuit acutely to LPS116. Exposure of LPS (Serratia marcescens) at 100 or 500 μg mL–1 rapidly depressed the evoked neural circuit (Fig. 9a). This action potentially explains the reduced responsiveness in the intact larvae to sensory stimuli when fed LPS. The effects on the in situ central circuit are reversible with short (10 min) exposures and flushing with fresh saline. Thus, this suggests some direct actions on neurons and synaptic transmission within the neural circuit. It is known the sensory neurons in larval Drosophila are cholinergic with both muscarinic and nicotinic receptors residing in the larval CNS but exactly where the receptors are in the circuitry used in this study are unknown117-121. The nicotinic-cholinergic receptors at the frog neuromuscular junction showed no responsiveness to LPS, as spontaneous quantal responses were present while evoked responses where depressed (see above section on frog NMJ). This indicates nicotinic-cholinergic receptors are spared by direct influence by LPS during acute exposures. However, the presynaptic nerve terminals may not respond to depolarization as it appears for the frog NMJ.
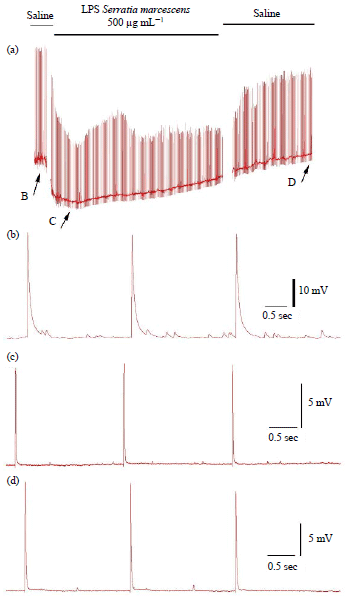
Since the neural circuitry of Drosophila share some similarities with the crayfish in that sensory neurons are also cholinergic and there are local segmental circuits, an established sensory-CNS-motor neuron circuit122,123 was also examined in the crayfish model to access the effects of LPS exposure41. Like for the larval Drosophila CNS preparation, the direct exposure to LPS (Serratia marcescens) on an in situ sensory-CNS-motor circuit produces a decrease in recruiting of the motor neurons at 500 μg mL–1 (Fig. 9b, c). No effect was noted at 100 μg mL–1 over a 10 min exposure. The depressed neural circuit continued to be reduced with longer exposure of 20 min. Activity does not recover with removal of the LPS and extensive flushing with fresh saline41. It is not established where the site of action for LPS is within this sensory-CNS-motor neuron circuit. However, given the reduced activity of the motor nerve over the 10 and 20 min, it is likely not blocking cholinergic receptors within the circuit or blocking the ability to evoke action potentials in the stimulated sensory neurons. Potentially, there may be alterations in membrane potential of the neurons or slowly mediated responses to LPS altering membrane resistance of the various neurons within the circuit.
PRIMARY SENSORY NEURONS OF CRAYFISH AND CRAB
Given that the sensory-CNS-motor neuron circuits examined in the larval Drosophila and crayfish mentioned above demonstrated a depressed response with acute exposure to LPS it was important to examine if primary sensory neuron are affected by LPS. Stanback et al.124 addressed, in well-established electrophysiological models of primary sensory neurons of a freshwater crayfish and a marine crab, if LPS altered sensory transduction and electrical activity. The blue crab (Callinectes sapidus) offered another crustacean model to examine for the potential effects of LPS on sensory neurons.
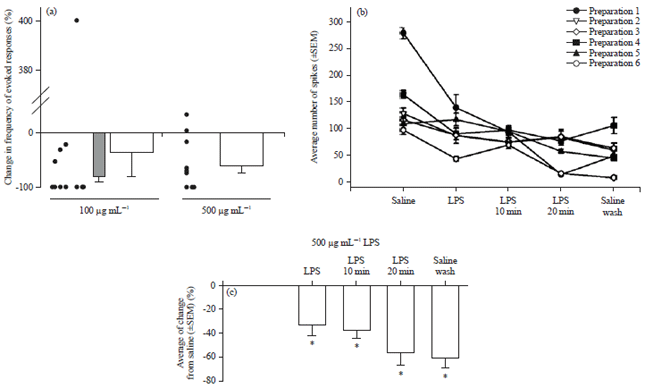 | |
| Fig. 9(a-c): | Effect of LPS exposure on the evoked sensory-CNS-motor neural circuit and synaptic transmission at the neuromuscular junction in 3rd instar Drosophila larvae and of a crayfish, (a) A stimulus train delivered to sensory roots of larval Drosophila every 10 sec before and during LPS exposure evoked a motor nerve recruitment which was indicated by the evoked EJPs measured in muscle 6. Average percent change in EJP frequency in response to 10 min of LPS exposure for 100 and 500 μg mL–1. The mean (±SEM) percent changes are shown as bars along with the changes in the individual preparations. The individual preparations are offset from each other in order to prevent overlap. The outlier at 400% increase for the 100 μg mL–1 was removed for additional analysis of a mean (±SEM) as seen in the hatched bar graph, (b) 500 μg mL–1 exposure to the neural circuit in the crayfish abdomen while providing sensory stimulation resulted in depressed neural activity. The average number of spikes recorded for each preparation for each condition over 10 min is shown and (c) Percent changes in sensory to motor nerve activity prior and after to LPS exposure. The 500 μg mL–1 resulted in depressed activity 10 min after flushing of the preparation with saline not containing LPS (p<0.05, ANOVA) |
The sensory neurons utilized in the crab were of the joint proprioceptor in the distal walking leg segment. This proprioceptive chordotonal organ, referred to as the propoditedactylopodite (PD) chordotonal organ, monitors joint activity by the rate of movement and static position of the joint between the propodite and dactylopodite leg segments. This proprioceptive organ contains both dynamically sensitive neurons, meaning that they fire only throughout the primary movement and statically sensitive neurons, which fire during the held various joint positions125-129. In the crayfish, the abdominal muscle receptor organ (MRO), which is analogous to the human muscle spindle, was utilized. In the MRO there are also two main types of sensory neurons, similarly to the crab PD organ. There are rapidly adapting and slow adapting sensory neurons, but each are associated with their separate muscle fibers130,131. In contrast, the sensory endings for the crab PD organ are embedded in a single chordotonal strand. The cell bodies are in close proximity to the sensory endings in these preparations. Thus, the sensory endings, cell bodies and axons, along with the locations where the site of spike initiation occur, are exposed to the bathing saline. The activity of the neurons was measured with suction electrodes of the cut nerves while moving and holding the various joints in positions before and during exposure to LPS.
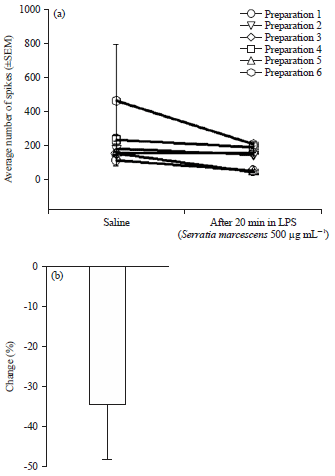 | |
| Fig. 10(a-b): | Activity of the PD organ before and during exposure to LPS from Serratia marcescens |
| (a) Activity of the initial displacement within 1 sec. The dynamic displacement was significant n = 7, p<0.05 non-parametric sign test, the effect of 500 μg mL–1 for 6 preparations are shown. The activity decreased as a result of LPS exposure. (b) Overall percent changes of the averaged (±SEM) values for activity in saline to LPS, source: Reproduced from Stanback et al.124 | |
In these set of experiments conducted by Stanback et al.124, two form of LPS were examined. Application of LPS from Serratia marcescens (500 μg mL–1) and Pseudomonas aeruginosa (500 μg mL–1) were examined individually as well as in a cocktail of the two combined (each at 500 μg mL–1).
There was no effect of either strain of LPS or the cocktail on sensory transduction and overall activity for the crayfish MRO preparation over 10 min of exposure. However, the activity of the dynamic sensory neurons of the crab PD organ did decrease in activity for Serratia marcescens (Fig. 10), but not for Pseudomonas aeruginosa or the cocktail of the two. The sample size of these studies was relatively low (n = 6 for each condition) and the effects were not drastic, but still convincing due to the trends. Longer incubation time and inducing evoked activity during exposure may relieve a different outcome. The discovery that LPS has a greater effect on the dynamic sensing neurons than the static position sensitive neurons may have to do with the slight changes in the bursting activity during the one second movement as compared to the static neurons firing over 8 sec. Minor changes in a smaller number of recorded extracellular spikes may be accentuated in the calculated percent changes to the effects of LPS.
CARDIOVASCULAR EFFECTS-DROSOPHILA AND CRAYFISH
The heart of Drosophila is used as a developmental and physiological model in research as it is myogenic and electrically paces as well as the organism is amenable to genetic manipulation132-140. Interestingly, the larval Drosophila heart tube is myogenic in a late stage of embryogenesis and by the late 3rd instar becomes neurally innervated141. In the adult, the heart is neurally influenced. Due to the ability to regulate what is expressed in cardiac tissue in the larval heart, it has served as the experimental model to understand mechanisms that control physiological function and provide a unique opportunity to evaluate novel experimental techniques, such as with expression of light sensitive ion channels and pumps to provide optogenetic techniques to be implemented and tested142,143. Since the myogenic heart of mammals is also altered in its electrical function in septicemia, it is of interest to investigate how the larval heart rate and physiology is altered by bacterial infection and exposure to LPS directly144-146. The larval heart tube circulates hemocytes, which act as the innate immune defense system, as well as bacteria and LPS present147. A recent study29 compared the effects of application of LPS from two common bacterial stains (Pseudomonas aeruginosa and Serratia marcescens), known to be a cause septicemia in humans and other mammals148-151, directly on the exposed heart tube. In addition to investigating Drosophila melanogaster, the larvae of a blow fly (Phaenicia sericata) was used in comparison since it is used in medical therapy for debriding dead tissue, which can be exposed to high levels of bacterial endotoxins. There is a dose-dependent effect with LPS exposure on heart rate in Drosophila, but not for the blow fly. Additionally, the effects of a cardiac modulator (i.e., serotonin152,153) was examined for differential function of serotonin alone or in combination with LPS29. Since the heart rate in larval Drosophila is very sensitive to the extracellular free calcium concentration lowering the concentration reduces heart rate. If LPS exposure resulted in a leaky membrane to Ca2+ ions a change in the heart rate would be more apparent with a lower rate than when it is beating at a fast rate with a higher concentration of calcium levels. This experimental approach was also examined in the study by Anyagaligbo et al.29.
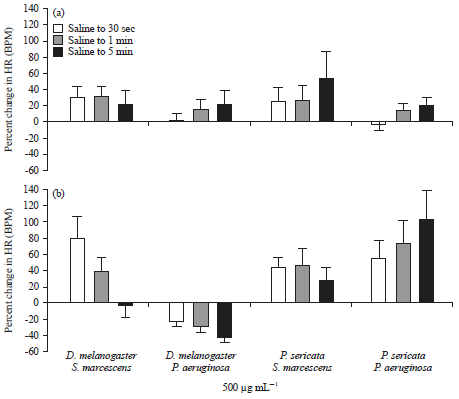 | |
| Fig. 11(a-b): | Average percent change in the Drosophila melanogaster and Phaenicia sericata preparations to the exposure of LPS from Serratia marcescens and Pseudomonas aeruginosa. The average percent change (±SEM) for each condition is made to the initial saline condition, (a) Percent changes for the exposure and (b) 500 μg mL–1 for both forms of LPS |
| Heart rate (HR) is reported as beats per minute (BPM), source: Reproduced from Anyagaligbo et al.29 | |
In this past study, it was demonstrated that direct exposure to LPS from two common strains of Gram-negative bacteria is able to induce changes on the heart rate of the fruit fly (Drosophila melanogaster) and blowfly (Phaenicia sericata) larvae. The effects of LPS on HR are dosage and time dependent (Fig. 11). In addition, the extracellular [Ca2+] alters the responsiveness to LPS. The HR of Drosophila melanogaster is very sensitive to very low extracellular [Ca2+] and cannot be electrical induced to contract; however, the blow fly heart continues to function in very low extracellular [Ca2+] and can be electrical stimulated to beat. The LPS induces a small transient hyperpolarization in cardiac muscle. The heart of larval Drosophila melanogaster remains sensitive to modulation by 5-HT with exposure to high levels of LPS, indicating that LPS does not likely deplete internal stores of Ca2+ from the ER and that the LPS does not block the 5-HT receptors or the 5-HT activated second messenger cascade (Fig. 12).
Since serotonin activates phospholipase Cβ (PLC), which results in the release of internal Ca2+ from the sarcoplasmic/endoplasmic reticulum137,154, it does not appear the exposure to LPS blocked this response as the exposure to serotonin, in the presence of LPS, still lead to substantial increases in the heart rate. The LPS exposure in a bathing media low in free Ca2+ ions, lead only to slight further reductions in heart rate as compared to the presences of a higher extracellular Ca2+ concentration. Since only a slight reduction in heart rate occurred with exposure to LPS with lowered Ca2+, as compared to a substantial decrease in rate with higher extracellular Ca2+, this implies that LPS may block Ca2+ entry as a mechanism to maintain the higher heart rate or that LPS promotes a Cl-influx or a K+efflux from the heart muscle. It is not known if the equilibrium potential for Cl– is more depolarized than the resting membrane potential in the larval heart as it is for the larval skeletal muscle80. In general, a hyperpolarization or blocking Ca2+ entry induced by direct action of LPS would result in a slowing of the heart rate.
As for investigating the in situ sensory-CNS-motor unit function mentioned above for the crayfish, addressing intact cardiac function and survival of the crayfish allows for an integrative approach on the whole animal to the effects of LPS exposure. Since the heart of the adult crayfish is neurogenic, requiring neural control for beating, any alterations in the neural circuitry involved with pacing the heart will be noted in recording of the heartbeat. The heart rate in crayfish is relatively easy to measure with an impedance technique155 and the influence of an injection of LPS into the hemolymph on heart rate acutely or over days can be obtained.
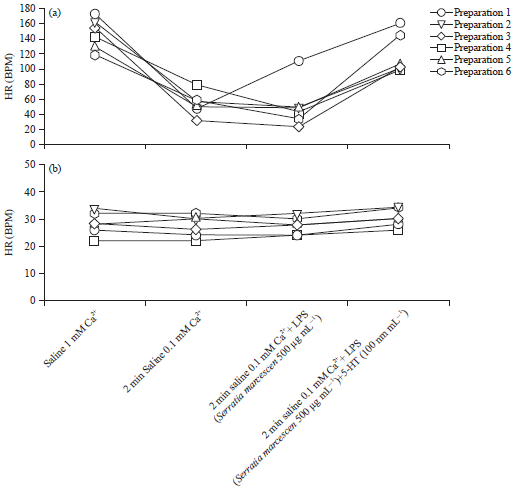 | |
| Fig. 12(a-b): | Effect of LPS on heart rate and a combination of LPS with serotonin in two species of larvae, (a) Drosophila melanogaster and (b) Phaenicia sericata. In both species a saline with reduced Ca2+ from 1-0.1 mM is used to slow down heart rate but only Drosophila melanogaster showed a reduction. In the presences of low Ca2+ LPS continued to reduce the Heart rate for the most of the Drosophila larvae but had little effect on Phaenicia sericata. With 0.1 mM Ca2+ combined with serotonin (5-HT, 100 nM) and LPS, heart rate increased for (a) Drosophila melanogaster but not (b) Phaenicia sericata |
| Source: Reproduced from Anyagaligbo et al.29 | |
Saelinger et al.41 conducted such experiments on the red swamp crayfish with two concentrations (500 and 5,000 μg mL–1) of estimated circulating levels of LPS from Serratia marcescens.
The injection of the vehicle control (saline only) and 500 μg mL–1 did not have a greater effect on heart rate, however, the 5,000 μg mL–1 produce a rapid increase in heart rate within 5 min and was maintained at a high rate over41 the next 12 h. The crayfish injected to a level of 5,000 μg mL–1 died within 24 h, whereas, the saline and lower level LPS-injected crayfish were still alive after 2 weeks at which point, they were not observed any longer (Fig. 13).
Since the effect of LPS exposure of the in situ sensory-CNS-motor unit in crayfish (mentioned above) resulted in a depression in activity, it does not appear that the neural circuit controlling the heartbeat was depressed. However, it is feasible that an inhibitor branch of the cardiac neural circuit was depressed more than the excitatory input which would lead to a heighten heart rate. A necropsy of the crayfish would have been beneficial to examine if the hemolymph may have clotted due to the innate immunity of crustaceans and horseshoe crabs1. If this were the case, then potentially the gills could be clogged resulting in a reduction in oxygen carrying capacity. If there is a cardiovascular compensation to potentially deliver oxygenated hemolymph then heart rate may increase. Crayfish have well-established neural control of the autonomic nervous system for integrating respiratory and cardiac function in various physiological states155-159. It remains unknown at this time why the crayfish exposed to high concentration of LPS exposure died but it is possible that other systemic effects induced by endotoxins such as cytokines, not yet identified in crustaceans, could be a factor.
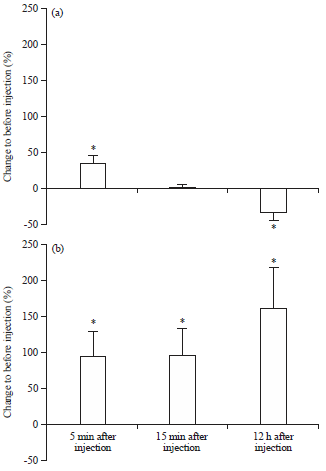 | |
| Fig. 13a-b): | Effects of LPS on heart rate in whole animals. The rates before and after injection of LPS (estimated circulating concentration) were used to calculate a percent change for, (a) 500 μg mL–1 and (b) 5,000 μg mL–1 (p<0.05, ANOVA compared to rate before injection) |
| Source: Reproduced from Saelinger et al.41 | |
CURRENT STUDIES UNDERWAY TO ADDRESS THE MECHANISM OF ACTION BY LPS
Genome analysis of Drosophila revealed 13 PGRPs with only three of the identified linked with the immune response160,161. Of these three PGRPs (PGRP-SA, PGRP-LC and PGRP-LE), PGRP-LC and PGRP-LE have been shown to respond to Gram negative bacteria162,163. The location and function of these proteins in various tissues of larval Drosophila has not yet been fully characterized. However, RNAi expressing lines have been developed by the Drosophila Transgenic RNAi Project (TRiP) at Harvard Medical School to knock down the protein expression of both PGRP-LC and PGRP-LE164. This will allow for the examination of physiological tissues of interest and their response or lack of response to direct exposure to LPS to determine functional significance of these PGRPs to LPS. The signaling pathways of these PGRP receptors are being addressed specifically in how they potentially alter function in the presynaptic terminal to effect synaptic transmission165. It is known that the immune deficiency (IMD) pathway is involved in the downstream cellular cascade activated by PGRP-LE and PGRP-LC receptors166.
Research findings of this study examined the effects of knocking down expression of peptidoglycan recognition proteins (PGRPs) PGRP-LC and PGRP-LE of body wall muscle, cardiac muscle, as well as, motor neurons via RNAi-mediated genetic approach. This will help to determine if this will block, in part or all, the direct acute actions of LPS and associated peptidoglycans from Serratia marcescens as previously reported. This present study is significant in aiding to identify the direct actions of LPS on targeted tissues in understand the actions independent of initiating an acquired or innate immune response. This may lead to potentially better approaches to combat bacterial infections in a wide range of organisms as well as understanding the basic physiological responses induced by LPS. Since this is a novel set of experiments and results, detailed methodology and results sections follow.
METHODS
Larval Drosophila melanogaster: The RNAi knockdowns of PGRPs were accomplished in muscle by crossing homozygous virgin females bearing the mesoderm driver 24B-Gal4 (Bloomington Drosophila Stock Centre or BDSC stock # 1767) to the males of dsRNA for RNAi of PGRP-LC ({TRiP.HMS00259} BDSC #33383) or PGRP-LE ({TRiP.HMC05031} BDSC # 60038). The motor neuron driverD42-Gal4 (BDSC # 8816 was crossed with the same dsRNA stocks to address the potential actions of LPS on motor neurons. These studies were conducted in May-August, 2019 at University of Kentucky, USA.
Electrophysiology in 3rd instar larvae: Fly saline hemolymph-like 3 (HL3)68,167 was used: (in mmol L–1) 70 NaCl, 5 KCl, 20 MgCl2, 10 NaHCO3, 1 CaCl2, 5 trehalose, 115 sucrose, 25 N,N-bis(2-hydroxyethyl)-2-aminoethanesulfonic acid (BES) and pH at 7.1
The early 3rd instar larval body wall muscle m6 was used to monitor the transmembrane potentials with sharp intracellular electrode (30-40 megaOhm resistance) filled with 3 M K-acetate. An Axoclamp 2B (Molecular Devices, Sunnyvale, CA, USA) amplifier and 1 X LU head stage was used. The EJPs and spontaneous mEJPs were collected and analyzed with LabChart 7.0 (ADInstruments, USA) as previously detailed79.
The LPS was dissolved in saline prior to use and was readily exchanged over the dissected preparations during the recording of evoked EJPs and mEJPs. Exposure to LPS for 10 min was used followed by changing the bathing saline 3 or 4 times with fresh saline not containing LPS. The total volume of the chamber is only 1 mL which is fully exchanged when switching the media. The LPS from Serratia marcescens was used. The LPS and chemicals used for saline were obtained from Sigma-Aldrich (St. Louis, MO, USA). Acute 10 min direct exposure of in situ NMJs to saline containing LPS (Serratia marcescens) at 500 μg mL–1 was used to compare to previous published reports for comparison in the action of LPS. Data were recorded as percent change from a saline solution to a saline solution containing the compound to normalize among preparations.
Heart rate measures in dissected larvae: Only early 3rd instar larvae were used (50-70 h post hatching). This stage was easily identified due to the small size of the Drosophila larvae (~5 mm) and the dissection to expose the heart tube is relatively feasible without damage. All larvae were maintained at room temperature ~21°C in vials partially filled with a cornmeal-agar-dextrose-yeast medium.
The general larval dissection technique to expose the larval heart tubes has been previously reported168. In brief, the larvae were dissected ventrally and pinned on four corners. The visceral organs were removed keeping the heart tube intact. This dissection technique was previously used to directly assess pharmacological agents on the heart of Drosophila larvae137,169,170. The dissection time was roughly 3-6 min and the muscles were allowed to relax while bathed in saline for 3-5 min after dissection. The heart rate was monitored and recorded after the initial dissection, after bathing 30 sec in LPS, after 1 min in LPS, after 5 min in LPS and lastly after 10 min in a saline wash as previously performed for monitoring actions of LPS on D. melanogaster, Canton-S strains29.
Statistical analysis: SigmaPlot (version 13.0) was used for graphing and statistical analysis. The electrophysiological analysis is presented as raw values and percent change from control (saline) or from LPS exposure to washout. There is considerable variation among baseline heart rates from preparation to preparation. Thus, raw values as well as percent changes are used. The paired t-test and non-parametric sign test were used for statistical analysis. A p<0.05 was considered as statistically significant.
RESULTS
Effects of LPS on heart rate: The transit increases in HR noted in Canton S with exposure to LPS also occurs for RNAi of PGRP-LE and PGRP-LC in the crosses with 24B (Fig. 14). The increase is short lived with just 1-2 min before the rate decreases. Control experiments of exchanging saline with saline as well as a dose-response (1, 100 and 500 μg mL–1) of LPS from Serratia marcescens exposure to the Canton-S strain were reported earlier29. As previously reported, an exchange of saline to saline does not result in a rapid rise in heart rate. Individual preparations are shown over time, as well as, an average (±SEM) of all preparations for each strain. As shown, there was individual variation in the HRs. However, a general trend in the response to LPS exposure was seen to be the same among individual preparations and indicated by significant increases in HR within the initial 1 sec after changing the bathing saline to one containing LPS for Canton-S (Fig. 14a), RNAi-PGRC-LE (Fig. 14b) and RNAi-PGRC-LC (Fig. 14c) (paired t-test, p<0.05, n = 10).
Effects of LPS on synaptic transmission and membrane potential with RNAi expressed in muscle: The wild type Canton S strain is known to show a transient hyperpolarization in the membrane potential of the body wall muscle and a decrease in the amplitude of the evoked EJP79. To examine if the PGRC-LC and PGRC-LE receptors mediate the hyperpolarization of the body wall muscles, RNAi was targeted for these receptors in the body wall muscles using the 24B line. Just as LPS (500 μg mL–1) results in the transient hyperpolarization in the body wall muscle (Fig. 15a, b, paired t-test p<0.05, n = 6) and depression of the evoked EJP amplitude (Fig. 15c, paired t-test p<0.05, n = 6) for Canton S (Fig. 15a), the same trends are also observed in the transit hyperpolarization in the body wall muscle (Fig. 16a, b (paired t-test p<0.05, n = 6) and depression of the evoked EJP amplitude (Fig. 16c, paired t-test p<0.05, n = 6) for RNAi-PGRC-LE (Fig. 3a) and transit hyperpolarization in the body wall muscle (Fig. 17a, b (paired t-test, p<0.05, n = 6) and depression of the evoked EJP amplitude (Fig. 17c, paired t-test p<0.05, n = 6) for RNAi-PGRC-LC (Fig. 17a).
Effects of LPS on synaptic transmission and membrane potential with RNAi expressed in motor neurons: In examining if the decrease in the evoked EJP amplitude induced by LPS was a result of a response mediated by the PGRC-LC and PGRC-LE receptors on the motor neurons, the RNAi lines were expressed in the D42 strain which targeted motor neurons.
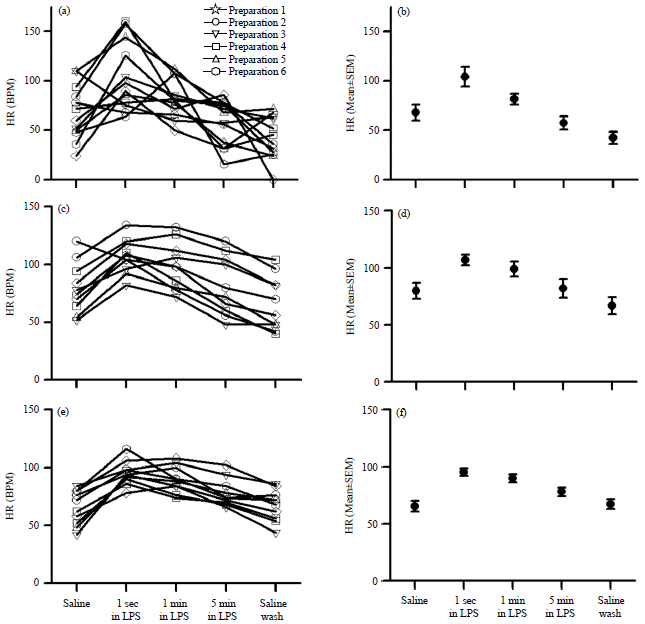 | |
| Fig. 14(a-f): | Changes in heart rate with exposure to LPS (500 μg mL–1) over time. (a, b) Individual preparations are shown with a line and different symbols in the left panels and a mean value (±SEM) of HR among all preparations in the right panel for Canton-S, (c, d) RNAi-PGRC-LE and (e, f) RNAi-PGRC-LC. In all Drosophila strains LPS resulted in a rapid increase in heart rate upon adding LPS and over time heart rate decreased. (*Paired t-test p<0.05, n = 10 for the saline to the first second of LPS exposure) |
Just as for the RNAi lines targeted in body wall muscles, LPS (500 μg mL–1) results in similar responses with a transit hyperpolarization in the body wall muscle and depression of the evoked EJP amplitude (Fig. 18a, 19a). Transit hyperpolarization in the body wall muscle (Fig. 18a, b, paired t-test p<0.05, n = 6) and depression of the evoked EJP amplitude (Fig. 18c, paired t-test p<0.05, n = 6) occurred for the RNAi-PGRC-LE and for RNAi-PGRC-LC (Fig. 19a-c, paired t-test p<0.05, n = 6).
Since there was some variation among preparations, a percent change in the initial values in saline to the maximal response by LPS was obtained. The average percent changes (±SEM) among the strains are shown (Fig. 20). There were no significant differences among the strains for changes in membrane potential or the changes in the amplitude of the EJPs (One way analysis of variance p>0.05).
Discussion: The use of the RNAi lines for the peptidoglycan recognition proteins (PGRPs) PGRP-LC and PGRP-LE in body wall muscle, cardiac muscle and motor neurons did not reveal any differences in responses to the commercial LPS from the RNAi lines for PGRP-LC and PGRP-LE expressed in either 24B or D42 or responses in the Canton S strain.
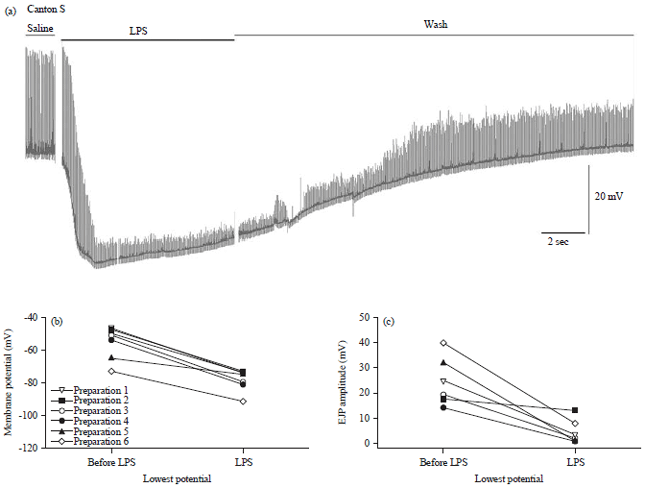 | |
| Fig. 15(a-c): | LPS (500 μg mL–1) depresses the amplitude of evoked EJPs and transiently hyperpolarizes the membrane potential of the larval body wall muscle in Canton S, (a) A representative trace illustrating the transit hyperpolarization of the muscle and reduction in the EJP amplitudes with exposure to LPS, (b) Transit hyperpolarization in the body wall muscle is shown for each preparation with different lines before and after application of LPS (Paired t-test p<0.05, n = 6) and (c) Depression of the evoked EJP amplitude before and acutely after LPS when the membrane potential is the most negative (Paired t-test p<0.05, n = 6) |
From the genomic analysis 13 PGRPs receptors were identified, but in previous studies only three (ie. PGRP-SA, PGRP-LC and PGRP-LE) demonstrated an immune response160,161. It was established that only the PGRP-LC and PGRP-LE responded to exposure of Gram-negative bacteria162,163. The RNAi expressing lines constructed by the Drosophila Transgenic RNAi Project for both PGRP-LC and PGRP-LE were used for specific expression in mesoderm (body wall and cardiac muscle) and motor neurons. Since it was shown that there were not any differences in the acute responses to LPS, it is feasible that the peptidoglycans associated with Serratia marcescens used in this study are not acting through PGRP-LC and PGRP-LE receptor subtypes. Alternative explanations for the lack of differences in response are that the RNAi knockdown larvae may retain sufficient activity to mediate a normal response or that the two receptors are redundant in interacting with LPS.
The depressed amplitudes from evoked EJPs and quantal responses (i.e. single vesicular events) on the body wall muscles are indicative that LPS exposure is blocking the glutamate receptors. The gradual decrease in the evoked EJPs and single quantal responses is strong evidence for this explanation. Furthermore, washing out the LPS containing saline partially recovers the evoked EJPs and quantal responses in a gradual manner, which aids in support of this notion. The glutamate receptor subtype at the NMJs of larval Drosophila melanogaster and crayfish are similar in their pharmacological profile75,76. The glutamate subunits partially resemble vertebrate AMPA and kainate receptors at the amino acid sequence level but do not share the same pharmacological profile as they do not respond well to AMPA, kainate or NMDA but are highly sensitive to quisqualate74-76.
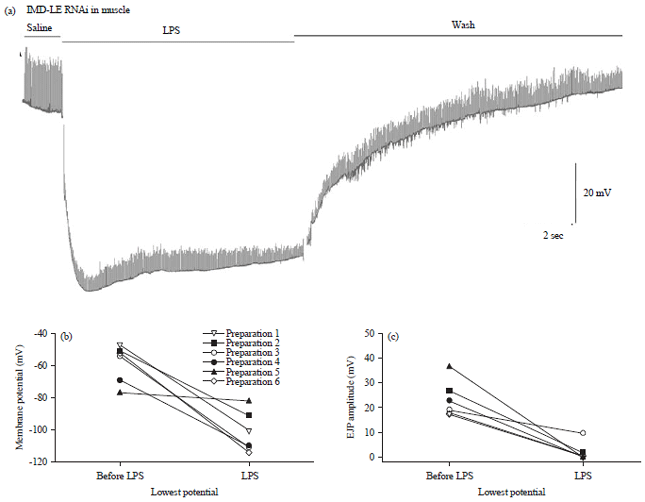 | |
| Fig. 16(a-c): | LPS (500 μg mL–1) depresses the amplitude of evoked EJPs and transiently hyperpolarizes the membrane potential of the larval body wall muscle of the RNAi-PGRC-LE x 24B strain, (a) A representative trace illustrating the transit hyperpolarization of the muscle and reduction in the EJP amplitudes with exposure to LPS, (b) Transit hyperpolarization in the body wall muscle is shown for each preparation with different lines before and after application of LPS (Paired t-test p<0.05, n = 6) and (c) Depression of the evoked EJP amplitude before and acutely after LPS when the membrane potential is the most negative (Paired t-test p<0.05, n = 6) |
It also appears the LPS induced transient hyperpolarization of the body wall muscles, which is not mediated by the PGRP-LC and PGRP-LE subtypes since there was still a large degree of hyperpolarization induced in the 24B-RNAi lines. The rationale to explain the mechanism behind the hyperpolarization remains elusive. The suggestive mechanisms of immune responses on tissues were previously examined. The effect does not appear to be due to calcium-activated potassium channels or activated nitric oxide synthase (NOS) or opening of Cl-channels79. It was postulated that if the Na-K ATPase pumps were transitory hyperactivated, this could explain the phenomenon in the large hyperpolarization phase of the LPS response79. However, no experimental evidence has been forth coming to substantiate this suggestion, so experiments are currently underway to address this by inhibiting the pump and examining the effects of LPS exposure.
The D42 strain was used to express the RNAi for PGRP-LC and PGRP-LE to examine if the effects induced by LPS might also be accompanied, in part, to the actions within the motor nerve terminal on vesicle docking and fusion machinery known to be linked with the IMD pathway70,165. However, if the blocking of the glutamate receptors on the muscle is a more pronounced effect than any slight actions in altering vesicular fusion kinetics, the actions would be masked. Given the induced LPS depression and effects of washing out LPS in depressing the amplitude of the quantal events and reappearance are so rapid, the acute actions do not appear to be long lasting on the vesicle docking or fusion machinery.
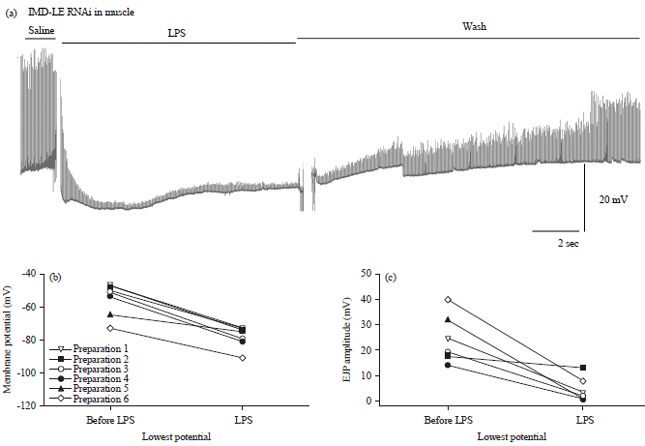 | |
| Fig. 17(a-c): | LPS (500 μg mL–1) depresses the amplitude of evoked EJPs and transiently hyperpolarizes the membrane potential of the larval body wall muscle of the RNAi-PGRC-LC x 24B strain, (a) A representative trace illustrating the transit hyperpolarization of the muscle and reduction in the EJP amplitudes with exposure to LPS, (b) Transit hyperpolarization in the body wall muscle is shown for each preparation with different lines before and after application of LPS (Paired t-test p<0.05, n = 6) and (c) Depression of the evoked EJP amplitude before and acutely after LPS when the membrane potential is the most negative (Paired t-test p<0.05, n = 6) |
The effects of LPS on the larval heart rate in these two RNAi lines were the same as previously reported for CS lines. As with the hyperpolarization in the body wall muscles, the mechanisms that causes a transient change in heart rate are not known. Although, it appears it is not from the release of internal stores of Ca2+ from the ER since even after the heart rate is depressed with low external bathing Ca2+, the heart rate did not increase with exposure to LPS and remained sensitive to serotonin. As with the body wall muscles, there is a slight hyperpolarization of the cardiac tissue with LPS exposure29.
Current studies are underway to image the nerve terminals with Ca2+ indicators, which will address if the motor nerve terminals and/or body wall muscles show any transient differences in Ca2+ loading with exposure to LPS. Future studies should continue to address the potential mechanisms to explain the presented acute observations, as it appears the PGRP-LC and PGRP-LE are not responsible for the effects. However, these receptors likely account for chronic responses previously reported due to exposures of Gram-negative bacteria171. It would be of interest to know if the acute actions presented in this study play some role in the chronic immune responses or if they are independent. Specifically, a basic understanding may help in treatment of septicemia in humans and other mammals.
In considering the NMJ, muscle and CNS preparations of Drosophila, crayfish and rodents preparations are bathed in a saline with a much larger volume of saline than the localized environments would not provide support for trophic factors being released from surrounding supportive cells, activated by LPS to influence the cell of interest. Also given that many the responses presented are within seconds of LPS application (i.e., muscle hyperpolarization and blocking of larval glutamate receptors at the NMJ) it would appear the effects are directly due to LPS on the muscle. If PGRPs are activated by LPS and the IMD pathway is used for nuclear translocation of Relish (a cytoplasmic protein) to trigger a response in the cell of contact this would to require minutes rather than seconds27,172.
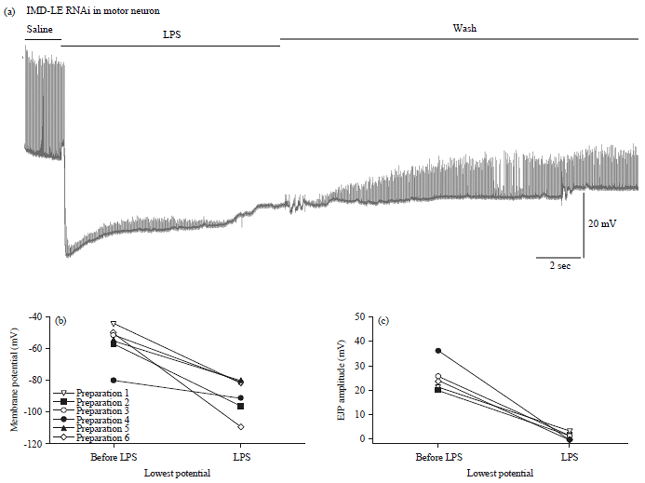 | |
| Fig. 18(a-c): | LPS (500 μg mL–1) depresses the amplitude of evoked EJPs and transiently hyperpolarizes the membrane potential of the larval body wall muscle of the RNAi-PGRC-LE x D42 strain, (a) A representative trace illustrating the transit hyperpolarization of the muscle and reduction in the EJP amplitudes with exposure to LPS, (b) Transit hyperpolarization in the body wall muscle is shown for each preparation with different lines before and after application of LPS (Paired t-test p<0.05, n = 6) and (c) Depression of the evoked EJP amplitude before and acutely after LPS when the membrane potential is the most negative (Paired t-test p<0.05, n = 6) |
It is feasible within the CNS models presented herein (above) that other cell types besides neurons are responsive to LPS and mediate some signal to influencing the neuronal activity, but with the speed of action it is unlikely to involve protein synthesis via IMD pathway used for nuclear translocation and excretion of a product. There are several antimicrobial peptides (AMPs) in the hemolymph of insects and crustaceans27. These AMPs may also reside between cells within the CNS and effect function if they rapidly aggregate, but the recovery of the responses would not be expected to be so rapid upon washing away the LPS response if this were the case.
Conclusion: The comparative studies with the same concentrations, types and stock of LPS have provided an informative approach for addressing commonalties in the direct effects of LPS. However, there are many puzzling effects in the different experimental models. One of the most puzzling findings, unknown before last year, is the fact that LPS in skeletal muscles of Drosophila and crayfish have a large transient hyperpolarization (i.e., 10-20 mV) with an unidentified underlying mechanism. A second result presented within this past year is the apparent blocking of glutamate receptors at the Drosophila NMJ, but not for the glutamatergic receptors at the crayfish NMJ. Enhancing synaptic transmission at the crayfish NMJ may be due to the hyperpolarization of the muscle and/or enhancing evoked release within the presynaptic neuromuscular junction. Considering the Drosophila and crayfish skeletal muscle contract due to Ca2+ entry through plasma membrane voltage gated calcium channels, the hyperpolarization might be related in these 2 species to this commonality.
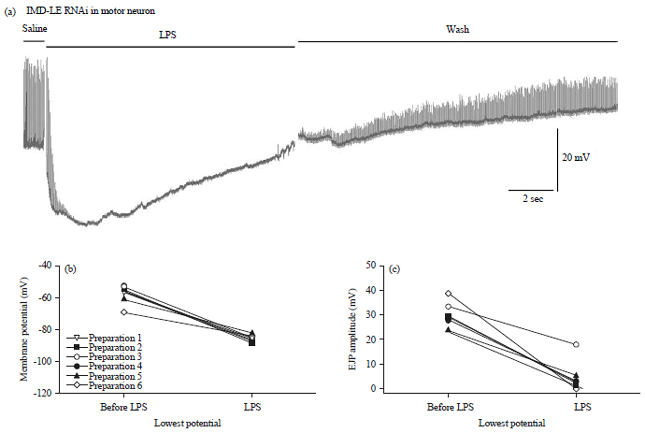 | |
| Fig. 19(a-c): | LPS (500 μg mL–1) depresses the amplitude of evoked EJPs and transiently hyperpolarizes the membrane potential of the larval body wall muscle of the RNAi-PGRC-LC x D42 strain, (a) A representative trace illustrating the transit hyperpolarization of the muscle and reduction in the EJP amplitudes with exposure to LPS, (b) Transit hyperpolarization in the body wall muscle is shown for each preparation with different lines before and after application of LPS (Paired t-test p<0.05, n = 6) and (c) Depression of the evoked EJP amplitude before and acutely after LPS when the membrane potential is the most negative (Paired t-test p<0.05, n = 6) |
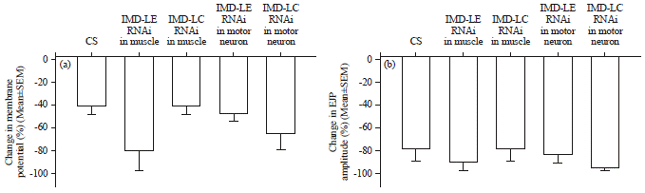 | |
| Fig. 20(a-b): | Averages in the percent change of the initial values in saline to the maximal response by LPS for the strains used. The average percent changes (±SEM) among the strains illustrated no significant differences among the strains for changes in, (a) Membrane potential (top panel) and (b) Changes in the amplitude of the EJPs (top panel) |
| One way analysis of variance p>0.05 | |
The frog and rodent muscle do not demonstrate the hyperpolarization with exposure to LPS. Contraction of these muscles are not dependent on extracellular entry of Ca2+ during contraction, but instead, intracellular release from sarcoplasmic/endoplasmic reticulum. Likewise, the Drosophila larval heart is very dependent on extracellular Ca2+ and is also sensitive to direct exposure to LPS. Perhaps there is a link with Ca2+ entry and the effect of LPS.
The rapid effects on the larval Drosophila and crayfish sensory-CNS-motor neuron evoked neural circuits, as well as the hippocampal slice, is also likely due to direct effects on neurons. It is not likely that these fast effects are due to activating microglial or invading cells that then release factors in the saline bath to alter synaptic transmission as the actions are very rapid and upon upon exchanging the bathing media to remove LPS the effects are rapidly reversed. Knowing that LPS did not have an effect of the primary sensory neurons of the crayfish and only slight effects on the a subset of sensory responses on the crab PD sensory organ, this would suggest that these sensory neurons may have a low density of receptors or ion channels that LPS effects on these other tissues.
POTENTIAL FUTURE STUDIES TO UNDERSTAND THE DIRECT ACTIONS OF LPS ON PHYSIOLOGICAL FUNCTIONS
Considering that gram negative bacteria also are known to release other toxins besides LPS, such as repeats-in-toxin (RTX)173 and it has been demonstrated that a RTX toxin form from Bordetella pertussis, which interacts with adenylate cyclase, can result in a rapid K+efflux from sheep erythrocytes and Jurkat cells, a human T cell leukemia line174 it could be possible that with the commercially obtained LPS that some RTX toxin maybe a contaminate in the preparation. It is known that Serratia marcescens does possess an RTX toxin175. This possibility needs to be addressed by chemical analysis. However, since the same batch of LPS and concentration was used in the studies presented herein, there is still the mystery why the crayfish and insect muscle hyperpolarized while the frog and rodent muscle did not. This might mean that a possible contaminant of RTX works differently on frog, rodent and invertebrate muscles.
Suggestions for future investigations in order to learn more about the direct effects of LPS on tissues are to examine the commercially obtained LPS from the same manufacturer and screen for any repeats-in-toxin (RTX) contaminates.
It would be of use to express IMD receptors in some insect line (i.e. S2) at high levels to see electrophysiological effects when exposed to LPS. RNAi expression in the cultured cells to depress protein expression of any the endogenous IMD receptors or to demonstrate if there are non-IMD effects by LPS on these cell lines would be beneficial. Use bacteria or other cells to express high levels of IMD receptor proteins, in order to purify and develop antibodies in host animals would provide a means of staining the receptors on various tissues in insects and potentially crustaceans as well as to be used in Western blots. Isolation of rodent/frog heart in a perfusion system to examine the influence of LPS on evoked vagal or sympathetic neural drive, as well as the effect of LPS on the electrical properties of the myogenic properties of the hearts in the absence of circulating body cytokines from non-cardiac tissue.
It would also be helpful to know if LPS has an effect on force development in rodent/frog muscle independent of the synaptic input. Indirectly electrically stimulating the muscle with cholinergic blockers would allow one to determine if LPS has and effects on the contractile properties of skeletal muscle independent of neural input and to examining cultured primary mammalian neurons without a glial substrate would provide a means to assessing more of the direct effects of LPS on them such as membrane potential and ionic fluxes.
SIGNIFICANCE STATEMENT
This review and primary research is significant to illustrate a better understanding in the potential effects of LPS in humans and indicates that one animal model or tissue type is not necessarily representative of the effects of LPS in other animal and tissue models.
ACKNOWLEDGMENTS
This work was in part supported by a Student Chellgren Fellowship by the University of Kentucky (C.B.B.), a summer fellowship by the Neuroscience Program at the University of Kentucky (M.N.), by a Gertrude Flora Ribble Scholarship from the Department of Biology at the University of Kentucky (C.S), personally funds (R.L.C.), Department of Biology at University of Kentucky teaching fund for Bio 446 in supplies, National Institutes of Health R01AG033649 and T32AG057461 (A.G. and O.T.), work was partly supported by the William Paterson University Center for Research Summer Faculty Award (S.M.B). We thank Zachary R. Hettinger (University of Kentucky) for preparing muscle myotubes for initial studies with rodent muscle. We also thank Dr. Melody Danley (University of Kentucky) for help with the earlier studies at the frog NMJ referred to in this review.
REFERENCES
- Lorenzon, S., S. De Guarrini, V.J. Smith and E.A. Ferrero, 1999. Effects of LPS injection on circulating haemocytes in crustaceans in vivo. Fish Shell Immunol., 9: 31-50.
Direct Link - Pien, B.C., P. Sundaram, N. Raoof, S.F. Costa and S. Mirrett et al., 2010. The clinical and prognostic importance of positive blood cultures in adults. Am. J. Med., 123: 819-828.
CrossRefDirect Link - Carl, M.A., I.M. Ndao, A.C. Springman, S.D. Manning and J.R. Johnson et al., 2014. Sepsis from the gut: The enteric habitat of bacteria that cause late-onset neonatal bloodstream infections. Clin. Infect. Dis., 58: 1211-1218.
CrossRefDirect Link - Kim, S.B., Y.D. Jeon, J.H. Kim, J.K. Kim and H.W. Ann et al., 2015. Risk factors for mortality in patients with Serratia marcescens bacteremia. Yonsei Med. J., 56: 348-354.
CrossRefDirect Link - Palavutitotai, N., A. Jitmuang, S. Tongsai, P. Kiratisin and N. Angkasekwinai, 2018. Epidemiology and risk factors of extensively drug-resistant Pseudomonas aeruginosa infections. PloS One, Vol. 13, No. 2.
CrossRefDirect Link - Osborn, M.J., S.M. Rosen, L. Rothfield, L.D. Zeleznick and B.L. Horecker, 1964. Lipopolysaccharide of the gram-negative cell wall. Science, 145: 783-789.
CrossRefDirect Link - Eidelman, L.A., D. Putterman, C. Putterman and C.L. Sprung, 1996. The spectrum of septic encephalopathy: Definitions, etiologies and mortalities. J. Am. Med. Assoc., 275: 470-473.
CrossRefDirect Link - Wilson, J.X. and G.B. Young, 2003. Progress in clinical neurosciences: Sepsis-associated encephalopathy: Evolving concepts. Can. J. Neurol. Sci., 30: 98-105.
CrossRefDirect Link - Friedrich, O., M.B. Reid, G. van den Berghe, I. Vanhorebeek, G. Hermans, M.M. Rich and L. Larsson, 2015. The sick and the weak: Neuropathies/myopathies in the critically ill. Physiol. Rev., 95: 1025-1109.
CrossRefDirect Link - Tong, D.M. and Y.T. Zhou, 2017. No awakening in supratentorial intracerebral hemorrhage is potentially caused by sepsis-associated encephalopathy. Med. Sci. Monit., 23: 4408-4414.
CrossRefPubMedDirect Link - Al-Nassan, S. and H. Fujino, 2018. Exercise preconditioning attenuates atrophic mediators and preserves muscle mass in acute sepsis. Gen. Physiol. Biophys., 37: 433-441.
CrossRefDirect Link - Hahn, A., M. Kny, C. Pablo-Tortola, M. Todiras and M. Willenbrock et al., 2019. Serum amyloid A1 mediates myotube atrophy via Toll-like receptors. J. Cachexia, Sarcopenia Muscle.
CrossRefDirect Link - Ono, Y. and K. Sakamoto, 2017. Lipopolysaccharide inhibits myogenic differentiation of C2C12 myoblasts through the Toll-like receptor 4-nuclear factor-κB signaling pathway and myoblast-derived tumor necrosis factor-α. PloS One, Vol. 12, No. 7.
CrossRefDirect Link - Cimolai, M., S. Alvarez, C. Bode and H. Bugger, 2015. Mitochondrial mechanisms in septic cardiomyopathy. Int. J. Mol. Sci., 16: 17763-17778.
CrossRefDirect Link - Gao, M., X. Wang, X. Zhang, T. Ha and H. Ma et al., 2015. Attenuation of cardiac dysfunction in polymicrobial sepsis by microRNA-146a is mediated via targeting of IRAK1 and TRAF6 expression. J. Immunol., 195: 672-682.
CrossRefDirect Link - Liu, Y.C., M.M. Yu, S.T. Shou and Y.F. Chai, 2017. Sepsis-induced cardiomyopathy: Mechanisms and treatments. Front. Immunol., Vol. 8.
CrossRefDirect Link - Shashikumar, S.P., M.D. Stanley, I. Sadiq, Q. Li, A. Holder, G.D. Clifford and S. Nemati, 2017. Early sepsis detection in critical care patients using multiscale blood pressure and heart rate dynamics. J. Electrocardiol., 50: 739-743.
CrossRefDirect Link - Walley, K.R., 2018. Sepsis-induced myocardial dysfunction. Curr. Opin. Crit. Care, 24: 292-299.
CrossRefDirect Link - Wang, Z., L. Bu, P. Yang, S. Feng and F. Xu, 2019. Alleviation of sepsis-induced cardiac dysfunction by overexpression of Sestrin2 is associated with inhibition of p-S6K and activation of the p-AMPK pathway. Mol. Med. Rep., 20: 2511-2518.
CrossRefDirect Link - Kaneko, T., W.E. Goldman, P. Mellroth, H. Steiner and K. Fukase et al., 2004. Monomeric and polymeric gram-negative peptidoglycan but not purified LPS stimulate the Drosophila IMD pathway. Immunity, 20: 637-649.
CrossRefDirect Link - Yoshida, H., K. Kinoshita and M. Ashida, 1996. Purification of a peptidoglycan recognition protein from hemolymph of the silkworm, Bombyx mori. J. Biol. Chem., 271: 13854-13860.
CrossRefDirect Link - Steiner, H., 2004. Peptidoglycan recognition proteins: On and off switches for innate immunity. Immunol. Rev., 198: 83-96.
CrossRefDirect Link - Levin, T.C. and H.S. Malik, 2017. Rapidly evolving Toll-3/4 genes encode male-specific Toll-like receptors in Drosophila. Mol. Biol. Evol., 34: 2307-2323.
CrossRefDirect Link - Coscia, M., S. Giacomelli and U. Oreste, 2011. Toll-like receptors: an overview from invertebrates to vertebrates. Invert. Survival J., 8: 210-226.
Direct Link - Loker, E.S., C.M. Adema, S.M. Zhang and T.B. Kepler, 2004. Invertebrate immune systems-not homogeneous, not simple, not well understood. Immunol. Rev., 198: 10-24.
CrossRef - Kleino, A. and N. Silverman, 2014. The Drosophila IMD pathway in the activation of the humoral immune response. Dev. Compa. Immunol., 42: 25-35.
CrossRefDirect Link - Myllymäki, H., S. Valanne and M. Rämet, 2014. The Drosophila IMD signaling pathway. J. Immunol., 192: 3455-3462.
CrossRefDirect Link - Anyagaligbo, O., J. Bernard, A. Greenhalgh and R.L. Cooper, 2019. The effects of bacterial endotoxin (LPS) on cardiac function in a medicinal blow fly (Phaenicia sericata) and a fruit fly (Drosophila melanogaster). Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 217: 15-24.
CrossRefDirect Link - Augustine, G.J. and H. Kasai, 2007. Bernard Katz, quantal transmitter release and the foundations of presynaptic physiology. J. Physiol., 578: 623-625.
CrossRefDirect Link - Eccles, J.C., B. Katz and S.W. Kuffler, 1941. Nature of the "endplate potential" in curarized muscle. J. Neurophysiol., 4: 362-387.
CrossRefDirect Link - Feng, T.P. and Z.S. Dai, 1990. The neuromuscular junction revisited: Ca2+ channels and transmitter release in cholinergic neurones in Xenopus nerve and muscle cell culture. J. Exp. Biol., 153: 129-140.
Direct Link - Rudolf, R., M.M. Khan and V. Witzemann, 2019. Motor endplate-anatomical, functional and molecular concepts in the historical perspective. Cells, Vol. 8, No. 5.
CrossRefDirect Link - Grinnell, A.D., 1995. Dynamics of nerve-muscle interaction in developing and mature neuromuscular junctions. Physiol. Rev., 75: 789-834.
CrossRefDirect Link - Harlow, M.L., D. Ress, A. Stoschek, R.M. Marshall and U.J. McMahan, 2001. The architecture of active zone material at the frog's neuromuscular junction. Nature, 409: 479-484.
CrossRefDirect Link - Heuser, J.E., T.S. Reese and D.M.D. Landis, 1974. Functional changes in frog neuromuscular junctions studied with freeze-fracture. J. Neurocytol., 3: 109-131.
CrossRefDirect Link - Person, R.J., 1977. Endotoxin alters spontaneous transmitter release at the frog neuromuscular junction. J. Neurosci. Res., 3: 63-72.
CrossRefDirect Link - Person, R.J., 1979. Bacterial lipopolysaccharide depresses spontaneous, evoked and ionophore-induced transmitter release at the neuromuscular junction. J. Neurosci. Res., 4: 105-114.
CrossRefDirect Link - Person, R.J., 1979. Depression of spontaneous and ionophore-induced neurotransmitter release by Salmonella. Experientia, 35: 801-802.
CrossRefDirect Link - Saelinger, C.M., M.C. McNabb, R. McNair, S. Bierbower and R.L. Cooper, 2019. Effects of bacterial endotoxin on regulation of the heart, a sensory-CNS-motor nerve circuit and neuromuscular junctions: Crustacean model. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., Vol. 237.
CrossRefDirect Link - McNabb, M.C., C.M. Saelinger, M. Danley and R.L. Cooper, 2019. Effects of bacterial endotoxin lipopolysaccharides (LPS) on synaptic transmission at neuromuscular junction in an amphibian. Int. J. Zool. Res., 15: 38-42.
CrossRefDirect Link - Van Harreveld, A., 1936. A physiological solution for freshwater crustaceans. Proc. Soc. Exp. Biol. Med., 34: 428-432.
Direct Link - Atwood, H.L., 1973. An attempt to account for the diversity of crustacean muscles. Integr. Compa. Biol., 13: 357-378.
CrossRefDirect Link - Sherman, R.G. and H.L. Atwood, 1971. Synaptic facilitation: long-term neuromuscular facilitation in crustaceans. Science, 171: 1248-1250.
CrossRefDirect Link - Van Harreveld, A. and C.A.G. Wiersma, 1936. The triple innervation of the crayfish muscle. Proc. Natl. Acad. Sci. USA., 22: 667-667.
CrossRefPubMedDirect Link - Katz, B. and S.W. Kuffler, 1946. Excitation of the nerve-muscle system in Crustacea. Proc. R. Soc. Lond. B, 133: 374-389.
CrossRefDirect Link - Katz, B., 1949. Neuro‐muscular transmission in invertebrates. Biol. Rev., 24: 1-20.
CrossRefDirect Link - Wiersma, C.A.G., 1949. Synaptic facilitation in the crayfish. J. Neurophysiol., 12: 267-276.
CrossRefDirect Link - Dudel, J. and S.W. Kuffier, 1961. The quantal nature of transmission and spontaneous miniature potentials at the crayfish neuromuscular junction. J. Physiol., 155: 514-529.
PubMedDirect Link - Dudel, J. and S.W. Kuffler, 1961. Mechanism of facilitation at the crayfish neuromuscular junction. J. Physiol., 155: 530-542.
CrossRefDirect Link - Bittner, G.D., 1968. Differentiation of nerve terminals in the crayfish opener muscle and its functional significance. J. Gen. Physiol., 51: 731-758.
CrossRefDirect Link - Bittner, G.D., 1968. The differentiation of crayfish muscle fibers during development. J. Exp. Zool., 167: 439-456.
CrossRefDirect Link - Iravani, J., 1965. Membrandepolarisation der Muskelfasern des öffnermuskels des Flusskrebses auf Nervenreiz und Kaliumapplikation. Experientia, 21: 609-610.
CrossRefDirect Link - Cooper, R.L., L. Marin and H.L. Atwood, 1995. Synaptic differentiation of a single motor neuron: Conjoint definition of transmitter release, presynaptic calcium signals and ultrastructure. J. Neurosci., 15: 4209-4222.
CrossRefPubMedDirect Link - Cooper, R.L., B.A. Stewart, J.M. Wojtowicz, S. Wang and H.L. Atwood, 1995. Quantal measurement and analysis methods compared for crayfish and Drosophila neuromuscular junctions and rat hippocampus. J. Neurosci. Methods, 61: 67-78.
CrossRefDirect Link - Parnas, I., R. Reinhold and J. Fine, 1971. Synaptic transmission in the crayfish: increased release of transmitter substance by bacterial endotoxin. Science, 171: 1153-1155.
CrossRefDirect Link - Baierlein, B., A.L. Thurow, H.L. Atwood and R.L. Cooper, 2011. Membrane potentials, synaptic responses, neuronal circuitry, neuromodulation and muscle histology using the crayfish: Student laboratory exercises. J. Vis. Exp., Vol. 18.
CrossRefDirect Link - Budnik, V., M. Gorczyca and A. Prokop, 2006. Selected methods for the anatomical study of Drosophila embryonic and larval neuromuscular junctions. Int. Rev. Neurobiol., 75: 323-365.
CrossRefDirect Link - Jan, L.Y. and Y.N. Jan, 1976. Properties of the larval neuromuscular junction in Drosophila melanogaster. J. Physiol., 262: 189-214.
CrossRefDirect Link - Kurdyak, P., H.L. Atwood, B.A. Stewart and C.F. Wu, 1994. Differential physiology and morphology of motor axons to ventral longitudinal muscles in larval Drosophila. J. Compa. Neurol., 350: 463-472.
CrossRefDirect Link - Stewart, B.A., H.L. Atwood, J.J. Renger, J. Wang and C.F. Wu, 1994. Improved stability of Drosophila larval neuromuscular preparation in haemolymph-like physiological solutions. J. Comp. Physiol. A, 175: 179-191.
CrossRefPubMedDirect Link - Hales, K.G., C.A. Korey, A.M. Larracuente and D.M. Roberts, 2015. Genetics on the fly: A primer on the Drosophila model system. Genetics, 201: 815-842.
CrossRefDirect Link - Martin, C.A. and D.E. Krantz, 2014. Drosophila melanogaster as a genetic model system to study neurotransmitter transporters. Neurochem. Int., 73: 71-88.
CrossRefDirect Link - Mohr, S.E., Y. Hu, K. Kim, B.E. Housden and N. Perrimon, 2014. Resources for functional genomics studies in Drosophila melanogaster. Genetics, 197: 1-18.
CrossRefDirect Link - Atwood, H.L. and R.L. Cooper, 1996. Assessing ultrastructure of crustacean and insect neuromuscular junctions. J. Neurosci. Methods, 69: 51-58.
CrossRefDirect Link - Atwood, H.L. and R.L. Cooper, 1996. Synaptic diversity and differentiation: Crustacean neuromuscular junctions. Invert. Neurosci., 1: 291-307.
CrossRefDirect Link - Bhatt, D. and R.L. Cooper, 2005. The pharmacological and physiological profile of glutamate receptors at the Drosophila larval neuromuscular junction. Physiol. Entomol., 30: 205-210.
CrossRefDirect Link - Lee, J.Y., D. Bhatt, D. Bhatt, W.Y. Chung and R.L. Cooper, 2009. Furthering pharmacological and physiological assessment of the glutamatergic receptors at the Drosophila neuromuscular junction. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 150: 546-557.
CrossRef - Titlow, J.S. and R.L. Cooper, 2018. Glutamatergic Synthesis, Recycling and Receptor Pharmacology at Drosophila and Crustacean Neuromuscular Junctions. In: Biochemical Approaches for Glutamatergic Neurotransmission, Parrot, S. and L. Denoroy (Eds.). Humana Press, New York, USA., ISBN: 978-1-4939-7227-2, pp: 263-291.
- Badre, N.H. and R.L. Cooper, 2008. Reduced calcium channel function in drosophila disrupts associative learning in larva and behavior in adults. Int. J. Zool. Res., 4: 152-164.
CrossRefDirect Link - Xing, B., A.A. Long, D.A. Harrison and R.L. Cooper, 2005. Developmental consequences of NMJs with reduced presynaptic calcium channel function. Synapse, 57: 132-147.
Direct Link - Cooper, R.L., M. McNabb and J. Nadolski, 2019. The effects of a bacterial endotoxin LPS on synaptic transmission at the neuromuscular junction. Heliyon, Vol. 5, No. 3.
CrossRefDirect Link - Rose, U., C. Derst, M. Wanischeck, C. Marinc and C. Walther, 2007. Properties and possible function of a hyperpolarisation-activated chloride current in Drosophila. J. Exp. Biol., 210: 2489-2500.
CrossRefDirect Link - Stanley, C.E., A.S. Mauss, A. Borst and R.L. Cooper, 2019. The effects of chloride flux on Drosophila heart rate. Methods Protocols, Vol. 2, No. 3.
CrossRefDirect Link - De Nooij, J.C., C.M. Simon, A. Simon, S. Doobar and K.P. Steel et al., 2015. The PDZ-domain protein Whirlin facilitates mechanosensory signaling in mammalian proprioceptors. J. Neurosci., 35: 3073-3084.
CrossRefDirect Link - Kravtsova, V.V., A.M. Petrov, V.V. Matchkov, E.V. Bouzinova and A.N. Vasiliev et al., 2016. Distinct α2 Na, K-ATPase membrane pools are differently involved in early skeletal muscle remodeling during disuse. J. Gen. Physiol., 147: 175-188.
CrossRefDirect Link - Rothfield, L. and B.L. Horecker, 1964. The role of cell-wall lipid in the biosynthesis of bacterial Lipopolysaccharide. Proc. Natl. Acad. Sci. USA., 52: 939-946.
CrossRefPubMedDirect Link - Park, B.S. and J.O. Lee, 2013. Recognition of lipopolysaccharide pattern by TLR4 complexes. Exp. Mol. Med., Vol. 45, No. 12.
CrossRefDirect Link - Perry, V.H., M.D. Bell, H.C. Brown and M.K. Matyszak, 1995. Inflammation in the nervous system. Cur. Opin. Neurobiol., 5: 636-641.
CrossRefDirect Link - Allan, S.M. and N.J. Rothwell, 2001. Cytokines and acute neurodegeneration. Nat. Rev. Neurosci., 2: 734-744.
CrossRefDirect Link - Khairova, R.A., R. Machado-Vieira, J. Du and H.K. Manji, 2009. A potential role for pro-inflammatory cytokines in regulating synaptic plasticity in major depressive disorder. Int. J. Neuropsychopharmacol., 12: 561-578.
CrossRefDirect Link - Cunningham, A.J., C.A. Murray, L.A.J. O'Neill, M.A. Lynch and J.J. O'Connor, 1996. Interleukin-1β (IL-1β) and Tumour Necrosis Factor (TNF) inhibit long-term potentiation in the rat dentate gyrus in vitro. Neurosci. Lett., 203: 17-20.
CrossRefDirect Link - Golia, M.T., S. Poggini, S. Alboni, S. Garofalo and N.C. Albanese et al., 2019. Interplay between inflammation and neural plasticity: Both immune activation and suppression impair LTP and BDNF expression. Brain Behav. Immunity, 81: 484-494.
CrossRefDirect Link - Hopp, S.C., H.M. D'Angelo, S.E. Royer, R.M. Kaercher, A.M. Crockett, L. Adzovic and G.L. Wenk, 2015. Calcium dysregulation via L-type voltage-dependent calcium channels and ryanodine receptors underlies memory deficits and synaptic dysfunction during chronic neuroinflammation. J. Neuroinflam., Vol. 12, No. 1.
CrossRefDirect Link - Min, S.S., H.Y. Quan, J. Ma, J.S. Han, B.H. Jeon and G.H. Seol, 2009. Chronic brain inflammation impairs two forms of long-term potentiation in the rat hippocampal CA1 area. Neurosci. Lett., 456: 20-24.
CrossRefDirect Link - Velasco-Estevez, M., S.O. Rolle, M. Mampay, K.K. Dev and G.K. Sheridan, 2020. Piezo1 regulates calcium oscillations and cytokine release from astrocytes. Glia, 68: 145-160.
CrossRefDirect Link - Abudara, V., L. Roux, G. Dallérac, I. Matias and J. Dulong et al., 2014. Activated microglia impairs neuroglial interaction by opening Cx43 hemichannels in hippocampal astrocytes. Glia, 63: 795-811.
CrossRefDirect Link - Wu, X., Y.G. Lv, Y.F. Du, M. Hu and M.N. Reed et al., 2019. Inhibitory effect of INT-777 on lipopolysaccharide-induced cognitive impairment, neuroinflammation, apoptosis and synaptic dysfunction in mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry, 88: 360-374.
CrossRefDirect Link - Cox, F.F., D. Carney, A.M. Miller and M.A. Lynch, 2012. CD200 fusion protein decreases microglial activation in the hippocampus of aged rats. Brain Behav. Immun., 26: 789-796.
CrossRefDirect Link - Denieffe, S., R.J. Kelly, C. McDonald, A. Lyons and M.A. Lynch, 2013. Classical activation of microglia in CD200-deficient mice is a consequence of blood brain barrier permeability and infiltration of peripheral cells. Brain Behav. Immun., 34: 86-97.
CrossRefDirect Link - Lynch, M.A., 2020. Can the emerging field of immunometabolism provide insights into neuroinflammation? Prog. Neurobiol., Vol. 184.
CrossRefDirect Link - Pérez‐Otaño, I., M.K. McMillian, J. Chen, G. Bing, J.S. Hong and K.R. Pennypacker, 1996. Induction of NF‐kB‐like transcription factors in brain areas susceptible to kainate toxicity. Glia, 16: 306-315.
CrossRefDirect Link - Friedman, W.J., S. Thakur, L. Seidman and A.B. Rabson, 1996. Regulation of nerve growth factor mRNA by interleukin-1 in rat hippocampal astrocytes is mediated by NFκB. J. Biol. Chem., 271: 31115-31120.
CrossRefDirect Link - Hartlage-Rubsamen, M., R. Lemke and R. Schliebs, 1999. Interleukin-1β, inducible nitric oxide synthase and nuclear factor-κB are induced in morphologically distinct microglia after rat hippocampal lipopolysaccharide/interferon-γ injection. J. Neurosci. Res., 57: 388-398.
CrossRefPubMedDirect Link - Nomura, Y., 1998. A transient brain ischemia- and bacterial endotoxin-induced glial iNOS expression and NO-induced neuronal apoptosis. Toxicol. Lett., 102: 65-69.
CrossRefDirect Link - Liu, Y., Y. Zhang, X. Zheng, T. Fang and X. Yang et al., 2018. Galantamine improves cognition, hippocampal inflammation and synaptic plasticity impairments induced by lipopolysaccharide in mice. J. Neuroinflamm., Vol. 15.
CrossRefDirect Link - Zhang, F.X. and R.S. Xu, 2018. Juglanin ameliorates LPS-induced neuroinflammation in animal models of Parkinson's disease and cell culture via inactivating TLR4/NF-κB pathway. Biomed. Pharmacother., 97: 1011-1019.
CrossRefDirect Link - Khan, M.S., T. Ali, M.W. Kim, M.H. Jo, J.I. Chung and M.O. Kim, 2019. Anthocyanins improve hippocampus-dependent memory function and prevent neurodegeneration via JNK/Akt/GSK3β signaling in LPS-treated adult mice. Mol. Neurobiol., 56: 671-687.
CrossRefDirect Link - Rehman, S.U., T. Ali, S.I. Alam, R. Ullah and A. Zeb et al., 2019. Ferulic acid rescues LPS-induced neurotoxicity via modulation of the TLR4 receptor in the mouse hippocampus. Mol. Neurobiol., 56: 2774-2790.
CrossRefDirect Link - Beattie, M.S., A.R. Ferguson and J.C. Bresnahan, 2010. AMPA-receptor trafficking and injury-induced cell death. Eur. J. Neurosci., 32: 290-297.
CrossRefDirect Link - Beattie, M.S., G.E. Hermann, R.C. Rogers and J.C. Bresnahan, 2002. Cell death in models of spinal cord injury. Progr. Brain Res., 137: 37-47.
CrossRefDirect Link - McNally, L., Z. Bhagwagar and J. Hannestad, 2008. Inflammation, glutamate and glia in depression: A literature review. CNS Spectrums, 13: 501-510.
CrossRefDirect Link - Aravalli, R.N., P.K. Peterson and J.R. Lokensgard, 2007. Toll-like receptors in defense and damage of the central nervous system. J. Neuroimmune Pharmacol., 2: 297-312.
CrossRefDirect Link - Bowman, C.C., A. Rasley, S.L. Tranguch and I. Marriott, 2003. Cultured astrocytes express toll-like receptors for bacterial products. Glia, 43: 281-291.
CrossRefDirect Link - Okun, E., K.J. Griffioen and M.P. Mattson, 2011. Toll-like receptor signaling in neural plasticity and disease. Trends Neurosci., 34: 269-281.
CrossRefDirect Link - Olson, J.K. and S.D. Miller, 2004. Microglia initiate central nervous system innate and adaptive immune responses through multiple TLRs. J. Immunol., 173: 3916-3924.
CrossRefDirect Link - Tang, S.C., T.V. Arumugam, X. Xu, A. Cheng and M.R. Mughal et al., 2007. Pivotal role for neuronal toll-like receptors in ischemic brain injury and functional deficits. Proc. Natl. Acad. Sci. USA., 104: 13798-13803.
CrossRefDirect Link - Calvo-Rodriguez, M., C. de la Fuente, M. Garcia-Durillo, C. Garcia-Rodriguez, C. Villalobos and L. Nunez, 2017. Aging and amyloid β oligomers enhance TLR4 expression, LPS-induced Ca2+ responses and neuron cell death in cultured rat hippocampal neurons. J. Neuroinflamm., Vol. 14.
CrossRefDirect Link - Istas, O., A. Greenhalgh and R. Cooper, 2019. The effects of a bacterial endotoxin on behavior and sensory-CNS-motor circuits in Drosophila melanogaster. Insects, Vol. 10, No. 4.
CrossRefDirect Link - Buchner, E., 1991. Genes expressed in the adult brain of Drosophila and effects of their mutations on behavior: A survey of transmitter- and second messenger-related genes. J. Neurogenet., 7: 153-192.
CrossRefDirect Link - Gorczyca, M.G., V. Budnik, K. White and C.F. Wu, 1991. Dual muscarinic and nicotinic action on a motor program in Drosophila. J. Neurobiol., 22: 391-404.
CrossRefDirect Link - Malloy, C.A., E. Somasundaram, A. Omar, U. Bhutto, M. Medley, N. Dzubuk and R.L. Cooper, 2019. Pharmacological identification of cholinergic receptor subtypes: Modulation of locomotion and neural circuit excitability in Drosophila larvae. Neuroscience, 411: 47-64.
CrossRefDirect Link - Rohrbough, J. and K. Broadie, 2002. Electrophysiological analysis of synaptic transmission in central neurons of Drosophila larvae. J. Neurophysiol., 88: 847-860.
CrossRefDirect Link - Su, H. and D.K. O'Dowd, 2003. Fast synaptic currents in Drosophila mushroom body Kenyon cells are mediated by α-bungarotoxin-sensitive nicotinic acetylcholine receptors and picrotoxin-sensitive GABA receptors. J. Neurosci., 23: 9246-9253.
CrossRefDirect Link - Strawn, J.R., W.S. Neckameyer and R.L. Cooper, 2000. The effects of 5-HT on sensory, central and motor neurons driving the abdominal superficial flexor muscles in the crayfish. Comp. Biochem. Physiol. B: Biochem. Mol. Biol., 127: 533-550.
PubMedDirect Link - Bierbower, S.M. and R.L. Cooper, 2013. The mechanistic action of carbon dioxide on a neural circuit and NMJ communication. J. Exp. Zool., 319: 340-354.
CrossRef - Stanback, M., A.E. Stanback, S. Akhtar, R. Basham and B. Chithrala et al., 2019. The effect of lipopolysaccharides on primary sensory neurons in crustacean models. IMPULSE, Vol. 16.
Direct Link - Hartman, H.B. and E.G. Boettiger, 1967. The functional organization of the propus-dactylus organ in Cancer irroratus Say. Comp. Biochem. Physiol., 22: 651-663.
CrossRefDirect Link - Cooper, R.L. and H.B. Hartman, 1999. Quantification of responses from proprioceptive neurons in the limbs of the crab, Cancer magister. J. Exp. Zool., 284: 629-636.
CrossRef - Cooper, R.L., 2008. Proprioceptive neurons of chordotonal organs in the crab, Cancer magister Dana (Decapoda, Brachyura). Crustaceana, 81: 447-475.
CrossRefDirect Link - Dayaram, V., C.A. Malloy, S. Martha, B. Alvarez and I. Chukwudolue et al., 2017. The effect of CO2, intracellular pH and extracellular pH on mechanosensory proprioceptor responses in crayfish and crab. Am. J. Undergrad. Res., 14: 85-99.
Direct Link - Malloy, C., V. Dayaram, S. Martha, B. Alvarez and I. Chukwudolue et al., 2017. The effects of potassium and muscle homogenate on proprioceptive responses in crayfish and crab. J. Exp. Zool. Part A: Ecol. Integr. Physiol., 327: 366-379.
CrossRefDirect Link - Kuffler, S.W., 1954. Mechanisms of activation and motor control of stretch receptors in lobster and crayfish. J. Neurophysiol., 17: 558-574.
CrossRefDirect Link - Rydqvist, B., J.H. Lin, P. Sand and C. Swerup, 2007. Mechanotransduction and the crayfish stretch receptor. Physiol. Behav., 92: 21-28.
CrossRefDirect Link - Akasaka, T. and K. Ocorr, 2009. Drug discovery through functional screening in the Drosophila heart. Methods Mol. Biol., 577: 235-249.
CrossRefDirect Link - Badre, N.H., M.E. Martin and R.L. Cooper, 2005. The physiological and behavioral effects of carbon dioxide on Drosophila melanogaster larvae. Comp. Biochem. Physiol. A Mol. Integr. Physiol., 140: 363-376.
CrossRef - Bellen, H.J., C. Tong and H. Tsuda, 2010. 100 years of Drosophila research and its impact on vertebrate neuroscience: A history lesson for the future. Nat. Rev. Neurosci., 11: 514-522.
CrossRefDirect Link - Bier, E. and R. Bodmer, 2004. Drosophila, an emerging model for cardiac disease. Gene, 342: 1-11.
CrossRefDirect Link - Cripps, R.M. and E.N. Olson, 2002. Control of cardiac development by an evolutionarily conserved transcriptional network. Dev. Biol., 246: 14-28.
CrossRefDirect Link - Desai-Shah, M., A.R. Papoy, M. Ward and R.L. Cooper, 2010. Roles of the sarcoplasmic/endoplasmic reticulum Ca2+-ATPase, plasma membrane Ca2+-ATPase and Na+/Ca2+ exchanger in regulation of heart rate in Larval drosophila. Open Physiol. J., 3: 16-36.
CrossRefDirect Link - Ocorr, K., L. Perrin, H.Y. Lim, L. Qian, X. Wu and R. Bodmer, 2007. Genetic control of heart function and aging in Drosophila. Trends Cardiovasc. Med., 17: 177-182.
CrossRefDirect Link - Pandey, U.B. and C.D. Nichols, 2011. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev., 63: 411-436.
CrossRef - Wessells, R.J. and R. Bodmer, 2004. Screening assays for heart function mutants in Drosophila. Biotechniques, 37: 58-66.
CrossRefDirect Link - Johnstone, A.F.M. and R.L. Cooper, 2006. Direct innervation of the Drosophila melanogaster larval aorta. Brain Res., 1083: 159-163.
CrossRefDirect Link - Zhu, Y.C., E. Yocom, J. Sifers, H. Uradu and R.L. Cooper, 2016. Modulatory effects on Drosophila larva hearts: Room temperature, acute and chronic cold stress. J. Comp. Physiol. A., 186: 829-841.
CrossRefPubMedDirect Link - Zhu, Y.C., H. Uradu, Z.R. Majeed and R.L. Cooper, 2016. Optogenetic stimulation of Drosophila heart rate at different temperatures and Ca2+ concentrations. Physiol. Rep., Vol. 4, No. 3.
CrossRefDirect Link - Shankar-Hari, M., G.S. Phillips, M.L. Levy, C.W. Seymour and V.X. Liu et al., 2016. Developing a new definition and assessing new clinical criteria for septic shock: For the Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). J. Am. Med. Assoc., 315: 775-787.
CrossRefDirect Link - Singer, M., C.S. Deutschman, C.W. Seymour, M.S. Hari and D. Annane et al., 2016. The third international consensus definitions for sepsis and septic shock (Sepsis-3). J. Am. Med. Assoc., 315: 801-810.
CrossRefDirect Link - Jayaprakash, N., O. Gajic, R.D. Frank and N. Smischney, 2018. Elevated modified shock index in early sepsis is associated with myocardial dysfunction and mortality. J. Crit. Care, 43: 30-35.
CrossRefDirect Link - Myers, A.L., C.M. Harris, K.M. Choe and C.A. Brennan, 2018. Inflammatory production of reactive oxygen species by Drosophila hemocytes activates cellular immune defenses. Biochem. Biophys. Res. Commun., 505: 726-732.
CrossRefDirect Link - Mahlen, S.D., 2011. Serratia infections: From military experiments to current practice. Clin. Microbiol. Rev., 24: 755-791.
CrossRefDirect Link - Olexy, V.M., T.J. Bird, H.G. Grieble and S.K. Farrand, 1979. Hospital isolates of Serratia marcescens transferring ampicillin, carbenicillin and gentamicin resistance to other gram-negative bacteria including Pseudomonas aeruginosa. Antimicrob. Agents Chemother., 15: 93-100.
CrossRefDirect Link - Piening, B.C., C. Geffers, P. Gastmeier and F. Schwab, 2017. Pathogen-specific mortality in very low birth weight infants with primary bloodstream infection. PLoS ONE, Vol. 12.
CrossRefDirect Link - Villalon, N., N. Farzan and K. Freeman, 2018. Rate of bacteremia in the hemodialysis patient presenting to the emergency department with fever: A retrospective chart review. Int. J. Emerg. Med., Vol. 11.
CrossRefDirect Link - Dasari, S. and R.L. Cooper, 2006. Direct influence of serotonin on the larval heart of Drosophila melanogaster. J. Comp. Physiol. B, 176: 349-357.
CrossRefDirect Link - Johnson, E., T. Sherry, J. Ringo and H. Dowse, 2002. Modulation of the cardiac pacemaker of Drosophila: Cellular mechanisms. J. Compa. Physiol. B, 172: 227-236.
CrossRefPubMedDirect Link - Sanyal, S., T. Jennings, H. Dowse and M. Ramaswami, 2006. Conditional mutations in SERCA, the Sarco-endoplasmic reticulum Ca2+-ATPase, alter heart rate and rhythmicity in Drosophila. J. Comp. Physiol. B, 176: 253-263.
CrossRefDirect Link - Listerman, L.R., J. Deskins, H. Bradacs and R.L. Cooper, 2000. Heart rate within male crayfish: Social interactions and effects of 5-HT. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., 125: 251-263.
CrossRefPubMedDirect Link - Li, H., L.R. Listerman, D. Doshi and R.L. Cooper, 2000. Heart rate measures in blind cave crayfish during environmental disturbances and social interactions. Compa. Biochem. Physiol. Part A: Mol. Integr. Physiol., 127: 55-70.
CrossRefDirect Link - Schapker, H., T. Breithaupt, Z. Shuranova, Y. Burmistrov and R.L. Cooper, 2002. Heart rate and ventilatory correlative measures in crayfish during environmental disturbances and social interactions. Comp. Biochem. Physiol. A, 131: 397-407.
CrossRef - Shuranova, Z.P., Y.M. Burmistrov, J.R. Strawn and R.L. Cooper, 2006. Evidence for an autonomic nervous system in decapod crustaceans. Int. J. Zool. Res., 2: 242-283.
CrossRefDirect Link - Shuranova, Z.P., Y.M. Burmistrov and R.L. Cooper, 2003. Bioelectric field potentials of the ventilatory muscles in the crayfish. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., 134: 461-469.
CrossRefDirect Link - Leclerc, V. and J.M. Reichhart, 2004. The immune response of Drosophila melanogaster. Immunol. Rev., 198: 59-71.
CrossRefDirect Link - Werner, T., G. Liu, D. Kang, S. Ekengren, H. Steiner and D. Hultmark, 2000. A family of peptidoglycan recognition proteins in the fruit fly Drosophila melanogaster. Proc. Natl. Acad. Sci. USA., 97: 13772-13777.
CrossRefDirect Link - Gottar, M., V. Gobert, T. Michel, M. Belvin and G. Duyk et al., 2002. The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature, 416: 640-644.
CrossRefDirect Link - Takehana, A., T. Katsuyama, T. Yano, Y. Oshima, H. Takada, T. Aigaki and S. Kurata, 2002. Overexpression of a pattern-recognition receptor, peptidoglycan-recognition protein-LE, activates imd/relish-mediated antibacterial defense and the prophenoloxidase cascade in Drosophila larvae. Proc. Natl. Acad. Sci. USA., 99: 13705-13710.
CrossRefDirect Link - Perkins, L.A., L. Holderbaum, R. Tao, Y. Hu and R. Sopko et al., 2015. The transgenic RNAi project at Harvard Medical School: Resources and validation. Genetics, 201: 843-852.
CrossRefDirect Link - Harris, N., R.D. Fetter, D.J. Brasier, A. Tong and G.W. Davis, 2018. Molecular interface of neuronal innate immunity, synaptic vesicle stabilization and presynaptic homeostatic plasticity. Neuron, 100: 1163-1179.
CrossRefDirect Link - Martin, M., A. Hiroyasu, R.M. Guzman, S.A. Roberts and A.G. Goodman, 2018. Analysis of Drosophila STING reveals an evolutionarily conserved antimicrobial function. Cell Rep., 23: 3537-3550.
CrossRefDirect Link - De Castro, C., J. Titlow, Z.R. Majeed and R.L. Cooper, 2014. Analysis of various physiological salines for heart rate, CNS function and synaptic transmission at neuromuscular junctions in Drosophila melanogaster larvae. J. Comp. Physiol. A, 200: 83-92.
CrossRefDirect Link - Cooper, A.S., K.E. Rymond, M.A. Ward, E.L. Bocook and R.L. Cooper, 2009. Monitoring heart function in larval Drosophila melanogaster for physiological studies. J. Vis. Exp., Vol. 33.
CrossRefDirect Link - Majeed, Z.R., A. Stacy and R.L. Cooper, 2014. Pharmacological and genetic identification of serotonin receptor subtypes on Drosophila larval heart and aorta. J. Comp. Physiol. B., 184: 205-219.
CrossRefPubMedDirect Link - Malloy, C.A., K. Ritter, J. Robinson, C. English and R.L. Cooper, 2015. Pharmacological identification of cholinergic receptor subtypes on Drosophila melanogaster larval heart. J. Comp. Physiol. B, 186: 45-57.
CrossRefPubMedDirect Link - Ramachandran, G., 2014. Gram-positive and gram-negative bacterial toxins in sepsis: A brief review. Virulence, 5: 213-218.
CrossRefDirect Link - Paquette, N., M. Broemer, K. Aggarwal, L. Chen and M. Husson et al., 2010. Caspase-mediated cleavage, IAP binding and ubiquitination: Linking three mechanisms crucial for Drosophila NF-κB signaling. Mol. Cell, 37: 172-182.
CrossRefDirect Link - Linhartova, I., L. Bumba, J. Masin, M. Basler and R. Osicka et al., 2010. RTX proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev., 34: 1076-1112.
CrossRef - Gray, M., G. Szabo, A.S. Otero, L. Gray and E. Hewlett, 1998. Distinct mechanisms for K+ efflux, intoxication and hemolysis by Bordetella pertussis AC toxin. J. Biol. Chem., 273: 18260-18267.
CrossRefDirect Link - Hertle, R., 2000. Serratia type pore forming toxins. Curr. Protein Pept. Sci., 1: 75-89.
CrossRefPubMedDirect Link