
Mulyadi
Department of Aquaculture, Faculty of Fisheries and Marine Science, Halu Oleo University, 93232 Kendari, JL. HEA Mokodompit, Anduonohu, South East Sulawesi, Indonesia
Indriyani Nur
Department of Aquaculture, Faculty of Fisheries and Marine Science, Halu Oleo University, 93232 Kendari, JL. HEA Mokodompit, Anduonohu, South East Sulawesi, Indonesia
LiveDNA: 62.27287
Wa Iba
Department of Aquaculture, Faculty of Fisheries and Marine Science, Halu Oleo University, 93232 Kendari, JL. HEA Mokodompit, Anduonohu, South East Sulawesi, Indonesia
International Journal of Zoological Research
Year: 2020 | Volume: 16 | Issue: 1 | Page No.: 1-11







ABSTRACT
Background and Objectives: Marine algae, especially brown algae (Sargassum sp.), is a natural compound plant capable of being prophylactic and immunostimulant. This study was aimed to evaluate the efficacy of seaweed (Sargassum sp.) extract to improve resistance and immune response of juvenile white shrimps (Litopenaeus vannamei) infected by Vibrio algynolyticus. Materials and Methods: The ethanolic extract of seaweed was used to evaluate its antibacterial effect by immerse the juvenile shrimp at a dose level of 0 (control), 150, 250 and 350 ppm for 3 h. Shrimp immune response was observed based on total haemocyte count (THC) and differential haemocyte count (DHC). Moreover, the bacterial challenge test was used for the evaluation of shrimp resistance. Results: The immersed shrimp in 150 ppm extract showed a significant increase in the number of THC and improve DHC value. During bacterial challenge test, shrimp juveniles immersed in 150 and 250 ppm had a 100% relative percent survival (RPS) which was higher than those in 350 ppm that had only 50% survival. Moreover, there were no significant histological changes of the hepatopancreas organ following infection in shrimps immersed in 150 and 250 ppm seaweed extract groups, whereas shrimps in control group showed hyperplasia and necrotic in nucleus cells. In addition, changes in the form of excess fat infiltration occured in tissues indicates the vulnerability of shrimp in control group. Conclusion: The study indicates that extract of brown seaweed Sargassum sp. is a potential immunostimulant to be used in juvenile white shrimps culture to control vibriosis.
PDF Abstract XML References Citation
How to cite this article
Mulyadi, Indriyani Nur and Wa Iba, 2020. Efficacy of Seaweed (Sargassum sp.) Extract to Prevent Vibriosis in White Shrimp (Litopenaeus vannamei) Juvenile. International Journal of Zoological Research, 16: 1-11.
DOI: 10.3923/ijzr.2020.1.11
URL: https://scialert.net/abstract/?doi=ijzr.2020.1.11
DOI: 10.3923/ijzr.2020.1.11
URL: https://scialert.net/abstract/?doi=ijzr.2020.1.11
INTRODUCTION
White leg shrimp (Litopenaeus vannamei ) is one of the world largest cultivated shrimp species1,2. Over the next few years, the increasing demand for white shrimps compelled intensive cultivation of this species, causing many issues due to increased disease outbreaks induced by microorganisms resulting in mass mortality3. Disease in white leg shrimp larvae and early juveniles caused by bacteria is an adverse problem in hatchery or nursery phase and will imminently affect its culture sustainability and productivity4,5.
The high mortality of larvae and early juveniles in shrimp hatchery as a result of vibrio bacterial strains infection or known as vibriosis disease is caused by bacteria V. harveyi, V. parahaemolyticus and V. alginolyticus5-8. While most of bacterial species were less pathogenic in nature, it has been recorded that vibrio bacterial diseases in penaeid shrimp cultivation systems involving at least 14 species, include Vibrio harveyi, V. splendidus, V. parahaemolyticus, V. alginolyticus, V. anguillarum, V. vulnificus, V. campbelli, V. fischeri, V. damsella, V. pelagicus, V. orientalis, V. ordalii, V. mediterrani and V. logei 9. Some control measures that may overcome such diseases including supervision of water quality, proper feeding and dosing appropriate immunostimulants.
Pathogen control is essential to prevent vibriosis outbreak in shrimp hatchery and nursery phase. Generally, the common practices in vibrio bacterial control is the application of synthetic chemicals such as antibiotics and drugs compounds during hatchery phase of the shrimp. However, owing to technical, operational, ecological and financial variables and constraints, this was not a very effective approach. In the last few years, the use of many forms of synthetic chemicals in the pathogen control program has been restricted. It is partly due to high cost of synthetic antibiotics, concern about negative effects over the environment, harmful effects on human health and other non-target populations, their non-biodegradable nature, high rate of ecosystem biological magnification and increasing antibiotic resistance10. This indiscriminate and unabated use of chemicals and antibiotics, however, has resulted an intense discussion among environmentalist and government agencies to prohibit these products completely. These factors put forward for the growth of a broad range of shrimp aquaculture sustainability approaches. Thus, the concept of pathogen disease control in aquaculture, particularly using herbal and phytopharmaca for disease prevention has received wide spread attention during the last decade11,12.
Similar to their terrestrial counterpart, marine plants (seaweeds or macroalgae) are also regarded as a wealthy source of bioactive molecules13,14. Some seeweed species were recognized and extracted for a broad range of bioactive compounds with various pharmacological functions, including antioxidants, soluble nutritional fibres, proteins, minerals, vitamins, phytochemicals and polyunsaturated fatty acids15. Marine algae, particularly brown algae (Sargassum sp.), is a natural compound factory that has the capacity as a prophylactic and immunostimulant material. The active ingredients in Sargassum sp. were described in our previous study including tannin, saponin, β-carotene, flavonoids, alkaloids, phenolic, steroid and glycose16. Alginic acid17 and fucoidan18 from brown algae and K-carrageenan from red algae19 showed a role as an immune trigger which helps aquatic animals in preventing various diseases20. Provision of natural immunity triggers proven to be able to optimize the health profile of aquatic animals including various immunity parameters such as total hemocyte count (THC) and phagocytic activity21,22.
The research conducted by Yeh et al.22 showed that administration of S. duplicatum extract on white leg shrimp juvenile could strengthen its immunity during challenge test with pathogenic bacteria. The increase in immunity was due to the polysaccharides extract containing immunostimulants, specifically fucoidan and alginic acid. The intake of bioactive materials also reduces the vulnerability of aquatic animals to various types of diseases such as vibriosis and viral diseases such as white syndrome viruses18.
Application of immunostimulants to mitigate vibriosis infections in white shrimps juveniles needs to take into account the appropriate dosages and administration time. Yeh et al.22 and Huynh et al.23 found that the control of Vibrio sp. in a hatchery with the use of Sargassum sp. extract from Penghu Island, Taiwan, by soaking shrimp larvae for 3 h at a dose of 300 ppm, was able to enhance the immunity of white leg shrimp larvae whereas higher doses did not affect the immune parameters. The study implied that the use of high doses may affect the shrimp immunity and less economical. Therefore, in this study lower dosage of Sargassum sp. extract from Buton Straits, Indonesia was used to determine its efficacy as immunostimulant in white shrimp juveniles.
MATERIALS AND METHODS
Preparation of seaweed extract: Samples of seaweed Sargassum sp. were collected from Bonerita waters, Buton Strait of South East Sulawesi Province between April and May, 2018. Seaweed were air dried at room temperature without sunlight exposure. Samples were subsequently chopped and ground using food grade blender into powder form and stored in a cool and dry place before further processing. The dried and ground seaweed was extracted using ethanol solution (1:4, w /v). After the maceration process with ethanol for 24 h, the extract was filtered and dried. Then, the extract was filtered using filter paper and the solvent evaporated to dryness under reduced pressure at 35°C using a rotary vacuum24. The dried and powdered crude extracts were kept at 4°C and protected from light and moisture until further use.
Experimental shrimp and rearing activity: Juvenile white shrimps were acquired from the hatchery of Benur Kita, Barru, South Sulawesi, Indonesia and was raised at shrimp hatchery in Mata, Kendari, SE-Sulawesi, Indonesia until they reached juvenile size of 4-5 cm. Shrimps from the hatchery were reared for 2 weeks for acclimatization purpose to experiment condition. They fed 5 times a day (every 4 h) using Artemia salina and artificial commercial feed combination at 5% of the shrimp body weight. Furthermore, experimental shrimps were randomly distributed into 12 aquariums (40×25×30 cm) with density of 15 shrimps/aquarium, referring to the optimal density in intensive ponds between25 100-300 shrimps m–2. Continuous aeration, siphoned and saline water renewal were applied to maintain clean and dissolved oxygen. During the experiment, water quality parameters were maintained on a normal level (temperature range was 28-30°C, DO>3 ppm and salinity was 33-35 ppt).
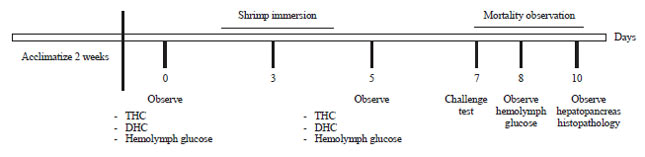
Experimental design: The experimental design of the present study was a complete randomized consisting of 3 treatments in triplicates. Sargassum extract was added to the water at levels of 0.0 (control), 150, 250 and 350 ppm. Juvenile shrimps were immersed for 3 h in aquarium with seaweed extract addition, then the shrimps were returned to normal water medium (Fig. 1).
Challenge test: After the 3 h immersion trial, the shrimps were challenged with V. algynolyticus. Infection was performed via an intramuscular injection of V. algynolyticus at a concentration of 107 CFU mL–1 using a 1 mL syringe and negative control 0.1 mL/individual shrimps were injected with Phosphate Buffered Saline. The clinical symptoms and mortality rates of the shrimps were observed for 72 h after injection.
Observed parameters: The experimental parameters observed were immune response, relative percent survival (RPS), hemolymph glucose, histopathology and clinical signs. The immune response parameters measured at 72 h post challenge test included total hemocyte count (THC) and differensial hemocyte count (DHC). The THC was calculated to discover the number of the shrimp’s hemocyte in reference to the method of Blaxhall and Daisley26, whereas, DHC calculation was based on Martin and Graves27.
Relative percent survival (RPS) versus control was calculated at the end of 72 h of infection using the formula by Amend28:
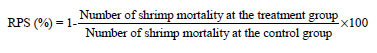
Clinical signs were observed following 6 h of challenge test. The level of glucose in the hemolymph was measured by applying a drop of hemolymph to a chemically treated, disposable ‘test-strip’, which was then inserted into an electronic blood glucose meter. The reaction between the test strip and the hemolymph is detected by the glucose meter and presented in mg dL–1 units29.
Following the challenge test, histological examination was done through microscopic identification of the tissues. Hepatopancreas were removed from each shrimp with clinical indications. Tissues were cut into tiny parts and fixed in 10% formaldehyde. Fixed tissues then were processed for histological preparation (following procedures of Takashima and Hibiya30 and Munford et al.31.
 | |
| Fig. 1: | Experimental design of the study |
Dehydration was used to remove water from the tissues with a series of alcohols: 70-95-100% and then followed by clearing with xylene. Finally, the embedding agent was infiltrated the tissue. Embedding process was performed by placing tissue in fresh paraffin wax and allowing paraffin to cool, then they were cut into sections that can be put on a slide. Slides were stained with haematoxylin and eosin (H and E). Prepared sections and stained were examined under a light microscope (Olympus BX53), photograph was taken (Olympus DP21 camera with Stream program) and histopathological changes were assessed.
Statistical analysis: The immune parameters obtained and RPS data were analyzed using the analysis of variance (ANOVA). Duncan’s test was used to determine significant differences (p<0.05) using SPSS Statistics 20.0 software. Histopathologal cause of bacterial infection was descriptively analyzed.
RESULTS
Hemolymph parameters: The levels of immune response were measured after 5 days of immersion treatment through recording total hemocyte count (THC) and differential hemocyte count (DHC). The highest number of THC was found in blood of shrimp juvenile immersed in 150 ppm of seaweed extract whereas, the lowest THC was in 350 ppm. It is suggested that the higher the concentration, the smaller the THC value (Fig. 2).
Observation on DHC directed at defining patterns of change of hyalin cells, semi-granular cells and granular cells proportion in hemolymph after extract Sargassum treatment. The H cells were the smallest cells with few granules, their nucleus occupied most of the space in cells. G cells were the biggest cells and contained abundant granules, while SG cells were smaller and contained less granules than G cells and were more round in common. Hyalin and semi-granular cells were the two dominant types of cells that demonstrate no distinctions in each treatments, whereas, semi-granular cells reduced significantly in elevated extract concentration (250 and 350 ppm) (Fig. 3, 4).
The data of haemolymph glucose in shrimps before and after treatments showed no significant difference. The average level of haemolymph glucose was 25 mg dL–1 in the initial of experiment and increased slightly at the end of extract exposure treatment at 33 mg dL–1. Juveniles shrimp immersion in the Sargassum extract at lower concentration did not increased the glucose levels, but pathogenic infection of Vibrio sp. caused significant shrimp stress which was characterized by a very dramatic rose in glucose level (107 mg dL–1) (Fig. 5).
Survival: Higher percentage of survived shrimp juveniles was found in those that were immersed in lower seaweed concentration compared to higher one. Survival of shrimps immersed in Sargassum extract showed that at 150 and 250 ppm treatments was 100% whereas, at 350 ppm, it was only 50% at 72 h post-challenge (Fig. 6).
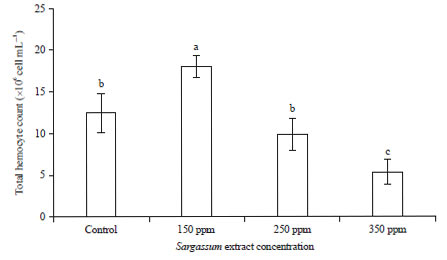 | |
| Fig. 2: | THC of white leg shrimp juveniles after immersed in seaweed extract at different concentration (Mean±SE) |
Different superscript letters indicated a significant difference among treatments (p<0.05) | |
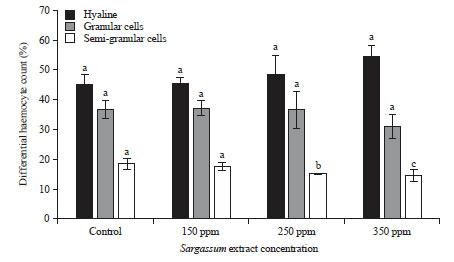 | |
| Fig. 3: | DHC of immersed vannamei shrimp in the different diluted extract dosages |
Different superscript letters indicated significant difference among treatments (p<0.05) | |
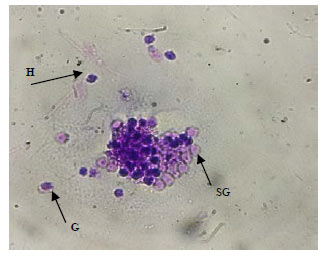 | |
| Fig. 4: | Morphology of shrimp hemocyte cells |
H: Hyaline, G: Granular, SG: Semi-granular | |
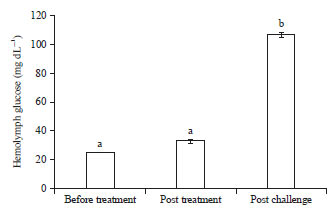 | |
| Fig. 5: | Level of hemolymph glucose of shrimps exposed by immersion in Sargassum extracts 150 ppm (treatment A) and challenged with Vibrio bacteria |
Histopathology: White leg shrimp juveniles that were challenged with Vibrio sp. infection without seaweed extract | |
Histopathology: White leg shrimp juveniles that were challenged with Vibrio sp. infection without seaweed extract immersion treatment showed some clinical signs such as necrosis of the tail and empty intestine (Fig. 7).
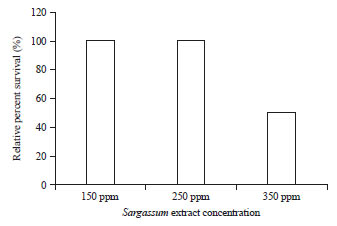 | |
| Fig. 6: | Relative percent survival (RPS) (%) of juvenile white leg shrimps after immersion and challenged with Vibrio sp., at different concentration of seaweed extract |
Histopathology analysis confirmed the clinical signs observation of shrimp juveniles without seaweed extract treatment. Hepatopancreas histopathology showed that those shrimp exhibited severe damage in their tissues include hyperplasia of cells (H), necrotic cells in nucleus (N) and excessive lipid cell infiltration/lipid degeneration (DL) (Fig. 8). Conversely, hepatopancreas of juvenile shrimps immersed in 150 ppm of seaweed extract showed no change or normal-appearing tissues (Fig. 9) whereas, those 250 ppm showed tissues with minimal damage as indicated by minimal amount of lipid degeneration (DL) and hyperplasia (H) without accompanied by necrotic cells found in tissues (Fig. 10). However, shrimp juveniles immersed in 350 ppm of seaweed extract showed a more moderate damage of tissues as shown by hyperplasia (H) and necrotic (N) with moderate intensity in cells (Fig. 11).
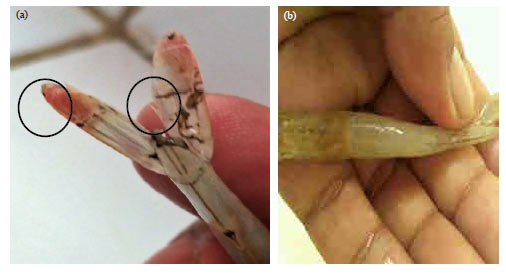 | |
| Fig. 7(a-b): | Clinical signs of shrimp in control group after 72 h shrimps were challenged with Vibrio bacteria. Infected shrimp showing necrosis of the (a) Tail and (b) Pale and empty intestine |
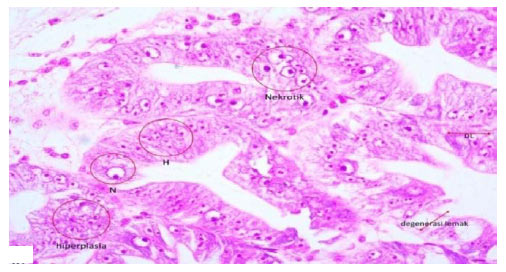 | |
| Fig. 8: | Histopathology of hepatopancreas in control group (shrimp juveniles without Sargassum sp. extract) |
H: Hyperplasia, N: Necrotic, DL: Lipid degeneration | |
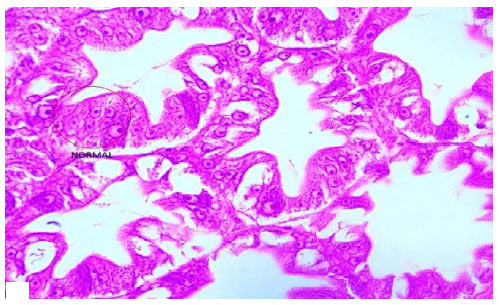 | |
| Fig. 9: | Hepatopancreas tissues in shrimp immersed with Sargassum sp. extract at 150 ppm |
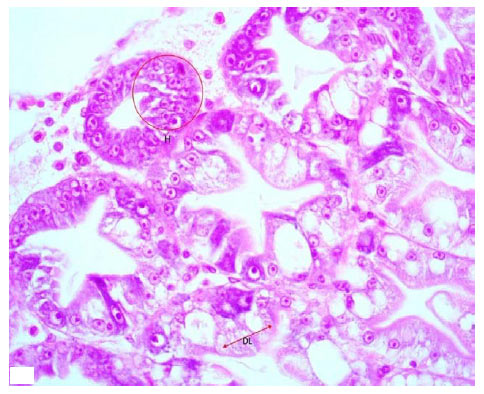 | |
| Fig. 10: | Histopathology of hepatopancreas in treatment B (shrimp with Sargassum sp. extract dose of 250 ppm) |
H: Hyperplasia, DL: Lipid degeneration | |
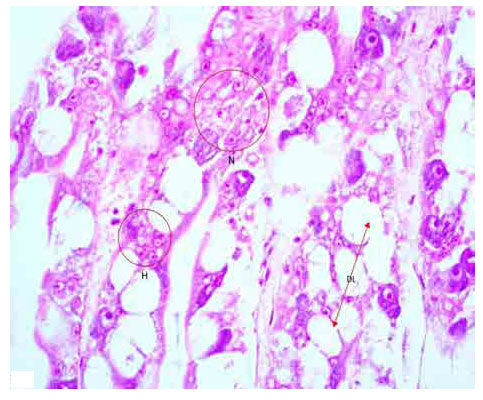 | |
| Fig. 11: | Histopathology of hepatopancreas in shrimp with Sargassum sp. extract at 350 ppm |
H: Hyperplasia, N: Necrotic, DL: Lipid degeneration | |
DISCUSSION
This study showed that lower concentration of seaweed extract may have a better effect on juvenile shrimps blood parameters after immersion in considerably short time compared to higher concentration or without seaweed extract. The higher number of THC of shrimp immersed in 150 ppm seaweed extract indicated that an appropriate concentration, Sargassum sp. bioactive compounds are capable of enhancing immune response due to the higher chance of creation of phagocytic cells, which is very important in controlling microorganism attacks32. Van de Braak33, Felix et al.21, Wang and Chen34 and Maftuch et al.35 stated that the increasing number of THC indicated a rise in both cellular and humoral immune response. In the current research, shrimp receiving the Sargassum extract through immersion showed an enhanced number of haemocytes that indicates haemocyte proliferation. This implied that immune system of white leg shrimps juvenile was ready to deal with pathogenic infections. However, THC significantly decreased at high concentration of Sargassum extract (350 ppm), suggesting that high concentration could paradoxically suppress immune responses (Fig. 2), thus resulting in being easily defeated by pathogens that ultimately leads to low survival (Fig. 6).
Stress is an animal disorder due to sources of inconvenient conditions. There is an increase in glucose hemolymph at the moment of stress to overcome the elevated demands for energy. In the view of Cuzon et al.36, if level of glucose in shrimp exceeds 150 mg dL–1, this indicates that the shrimp is stressed and requires higher energy. Herbal compounds have the capacity to inhibit oxygen anion generation and scavenge free radicals. It has been shown that the herbal antioxidant impact is comparable to that of superoxide dismutase, metal-ion chelators and xanthine oxidase inhibitors. Picrorhiza kurroa herb used as an antistress compound for shrimps is the finest example37. Stress increases susceptibility to diseases, possibly due to chronic elevated cortisol levels, resulting in immunosuppressive. Stress in crayfish can be indicated from the level of glucose in hemolymph29. This study obviously demonstrated a rise in glucose level up to 107 mg dL–1 due to infection with vibriosis, but stress has not escalated mortality as it was supported by the active compound of Sargassum for anti-stress (Fig. 5).
The hepatopancreas histological analysis showed the structural abnormalities in hepatopancreas infected with bacteria. Abnormalities could be seen in the control that was shrimp without immersion extraction treatment Sargasssum sp. Hepatopancreas is the most important organ for a shrimp organism. Hepatopancreas is a target organ for pathogen infection and environmental changes. Histopathological changes in hepatopancreas may be used as indicators for the assessment of the health of crustaceans especially stress levels and shrimp susceptibility38. Necrosis, haemorrhage and vacuolization of hepatopancreas tubule epithelial reduced cells were among the major disease-related histopathological changes observed in infected shrimps. Necrotic cells and tissues would experience decreased activity and eventually die. Munford et al.31 stated that necrotic tissue in an organism can be caused by several factors including bacteria that invade, injury to body parts, trauma, stress and the toxins in the waters. Necrotic is a morphological change which can lead to tissue cell death and shrinking of the nucleus size.
Symptoms of shrimp affected by vibriosis can be identified visually by observing directly both the physical and behavior of shrimps because a sick shrimp will show symptoms that are different from healthy shrimp. The results of the observation of V. alginolyticus bacteria on vannamei shrimp showed clinical symptoms and behavior of shrimp. The first day there was a decrease in shrimp appetite, marked shrimp intestinal contents were not full or intermittent and a lot of leftover feed on the basis of maintenance, but not accompanied by clinical symptoms that were significant from pathogenic infections. On day 2, the infection symptoms were seen in Fig. 7, the larvae looked weak and inactive, the head always faced up and the movements were uncontrolled, the shrimp tail was necrotizing, melanosis on the skin, the body of the shrimp looked pale, swimming legs, telson and uropod were reddish. Clinical symptoms detected in test shrimp indicated the presence of the Vibrio genus bacterial infection. Similar clinical symptoms have also been reported in shrimps with vibriosis39.
Biologically active substances in seaweed Sargassum sp. extracts in this study are tannin, saponin, β-carotene, flavonoids, alkaloids, phenolic, steroid and glycose16. Naturally, the main functions of plant secondary metabolites are to protect plants from attack by insects, herbivores and pathogens, or to survive from other biotic and abiotic stresses. Flavonoids are one of the compounds that play an important part in boosting the immune system. Flavonoids are plant secondary metabolites which have shown protective effects on cancer, heart diseases and retinal inflamation in human40,41. Other important metabolite in seaweed extract is β-carotene that is regarded as damaged cells protector by converting the β-carotene into vitamin A which in turn repairs the cells42. Alkaloids compounds in seaweed may damage the bacteria nucleic acids (DNA and RNA) as the basic structure of these alkaloids are alkylating agents and other substances that react covalently with purine and pyrimidine bases43. Saponins work in shrimp as antimicrobials, tannins function to inhibit bacteria by denaturing proteins and damaging membranes of bacterial cells by dissolving lipid contained in cell walls. Phenolic acts as a reduction agent, a supplier of hydrogen and a potential agent of chelation. These natural bioactive compounds were assumed have various pharmacologic activities of shrimps32, 44. Through a simple method, immersion with the right dose of immunostimulants could be improve the phagocytic capacity response.
The ability of Sargassum sp. used in this study to increase the immune system and prevent the emergence of juvenile shrimp disease might also be attributed to several important polysaccharides in the extract. The major polysaccharides that commonly found in seaweed are agar, carrageenan and alginates45. When lipopolysaccharides, peptidoglycans and β-1,3-glucan molecules are present-commonly found in bacteria and fungi46, cellular reaction, responsible for phagocytosis, melanisation, encapsulation and coagulation, is triggered. This has been proven in Arizo et al.47 study who found that properties of β-glucan present in seaweed Gracillaria edulis reacted with binding β-glucan proteins (βGBP) leading to haemocyte degranulation. Furthermore, the phytochemical pathway of brown seaweed Sargassum sp. in this study might provide additional protection for shrimps juvenile as shown by DHC number and cytoplasmic granules cells when administered at lower concentration (Fig. 3, 4).
The findings of this study showed that shrimp survival, hemolymph parameters and the histology of hepatopancreas were better when treated with 150 and 250 ppm Sargassum sp. This suggests that the sufficient amount of extract of Sargassum sp. to enhance immunity thus survival is between 150 and 250 ppm. More studies are needed regarding the inexpensive extraction methods for commercial applications of Sargassum extract, extract administration of either oral or immersion methods, its concentrations and the exposure time of shrimp to these extracts (in vivo).
CONCLUSION
In conclusion, white shrimp that immersed in the seawater containing the Sargassum extract 150 ppm showed significantly increased immune response by increasing the haemocyte count, proportional differential haemocyte, stabilizing hemolymph glucose level and high resistance against V. alginolyticus infection.
SIGNIFICANCE STATEMENT
This study discovered the beneficial effects of ethanolic extract of brown marine algae Sargassum sp. for the preventi on of vibriosis in the culture of juvenile white shrimp. This study will help the researchers to uncover the potential of marine algae to be used as natural antibiotic and immune stimulant in shrimp culture that many researchers were not able to explore. Thus a new concentration on delivering the marine algae extract may be arrived at.
ACKNOWLEDGMENTS
This research was supported by Postgraduate Research Grant 2019 from the Ministry of Research, Technology and Higher Education of the Republic of Indonesia, Contract No. 519e/UN29.20/PPM/2019.
REFERENCES
- Iba, W., M.A. Rice and G.H. Wikfors, 2014. Microalgae in Eastern pacific white shrimp, Litopenaeus vannamei (Boone 1931) hatcheries: A review on roles and culture environments. Asian Fish. Sci., 27: 212-233.
Direct Link - Saulnier, D., P. Haffner, C. Goarant, P. Levy and D. Ansquer, 2000. Experimental infection models for shrimp vibriosis studies: A review. Aquaculture, 191: 133-144.
CrossRefDirect Link - Liu, L., M. Ge, X. Zheng, Z. Tao, S. Zhou and G. Wang, 2016. Investigation of Vibrio alginolyticus, V. Harveyi and V. Parahaemolyticus in large yellow croaker, Pseudosciaena crocea (Richardson) reared in Xiangshan Bay, China. Aquacult. Rep., 3: 220-224.
CrossRefDirect Link - Widowati, I., M. Zainuri, H.P. Kusumaningrum, Y. Maesaroh and Y. Hardivillier et al., 2018. Identification of agents causing vibriosis in Litopenaeus vannamei shrimp culture in Kendal, Central Java, Indonesia and application of microalgae Dunaliella salina and Tetraselmis chui as bio-control agents against vibriosis. Aquacult. Aquar. Conserv. Legisl., 11: 101-107.
Direct Link - Austin, B. and D. Austin, 2007. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish. 4th Rev. Edn., Springer-Praxis Publishing, Chichester, UK., Pages: 552.
Direct Link - Grenni, P., V. Ancona and A.B. Caracciolo, 2018. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J., 136: 25-39.
CrossRefDirect Link - Bulfon, C., D. Volpatti and M. Galeotti, 2015. Current research on the use of plant‐derived products in farmed fish. Aquacult. Res., 46: 513-551.
CrossRefDirect Link - Reverter, M., N. Bontemps, D. Lecchini, B. Banaigs and P. Sasal, 2014. Use of plant extracts in fish aquaculture as an alternative to chemotherapy: Current status and future perspectives. Aquaculture, 433: 50-61.
CrossRefDirect Link - Thanigaivel, S., S. Vijayakumar, A. Mukherjee, N. Chandrasekaran and J. Thomas, 2014. Antioxidant and antibacterial activity of Chaetomorpha antennina against shrimp pathogen Vibrio parahaemolyticus. Aquaculture, 433: 467-475.
CrossRefDirect Link - Dhargalkar, V.K. and N. Pereira, 2005. Seaweed: Promising plant of the millennium. Sci. Cult., 71: 60-66.
Direct Link - Mulyadi, I. Nur and W. Iba, 2019. Phytochemical test of seaweed extract Sargassum sp. J. Fish. Sci. Innov., 3: 22-25.
Direct Link - Gioacchini, G., P. Smith and O. Carnevali, 2008. Effects of ergosan on the expression of cytokine genes in the liver of juvenile rainbow trout (Oncorhynchus mykiss) exposed to enteric red mouth vaccine. Vet. Immunol. Immunopathol., 123: 215-222.
CrossRefDirect Link - Immanuel, G., M. Sivagnanavelmurugan, T. Marudhupandi, S. Radhakrishnan and A. Palavesam, 2012. The effect of fucoidan from brown seaweed Sargassum wightii on WSSV resistance and immune activity in shrimp Penaeus monodon (Fab). Fish Shellfish Immunol., 32: 551-564.
CrossRefDirect Link - Bagni, M., N. Romano, M.G. Finoia, L. Abelli and G. Scapigliati et al., 2005. Short- and long-term effects of a dietary yeast β-glucan (Macrogard) and alginic acid (Ergosan) preparation on immune response in sea bass (Dicentrarchus labrax). Fish Shellfish Immunol., 18: 311-325.
CrossRefPubMedDirect Link - Felix, S., P. Robins and A. Rajeev, 2004. Immune enhancement assessment of dietry incorporated marine alga Sargassum wightii (Phaeophyceae/Punctariales) in tiger shrimp Penaeus monodon (Crustacia/Penaeidae) through prophenoloxidase (proPO) systems. Indian J. Mar. Sci., 33: 361-364.
Direct Link - Yeh, S.T., C.S. Lee and J.C. Chen, 2006. Administration of hot-water extract of brown seaweed Sargassum duplicatum via immersion and injection enhances the immune resistance of white shrimp Litopenaeus vannamei. Fish Shellfish Immunol., 20: 332-345.
CrossRefDirect Link - Huynh, T.G., S.T. Yeh, Y.C. Lin, J.F. Shyu, L.L. Chen and J.C. Chen, 2011. White shrimp Litopenaeus vannamei immersed in seawater containing Sargassum hemiphyllum var. chinense powder and its extract showed increased immunity and resistance against Vibrio alginolyticus and white spot syndrome virus. Fish Shellfish Immunol., 31: 286-293.
CrossRefDirect Link - Ye, H., C. Zhou, Y. Sun, X. Zhang, J. Liu, Q. Hu and X. Zeng, 2009. Antioxidant activities in vitro of ethanol extract from brown seaweed Sargassum pallidum. Eur. Food Res. Technol., Vol. 230.
CrossRefDirect Link - Samadan, G.M., Rustadi, Djumanto and Murwantoko, 2018. Production performance of whiteleg shrimp Litopenaeus vannamei at different stocking densities reared in sand ponds using plastic mulch. Aquacult. Aquar. Conservat. Legisl., 11: 1213-1221.
Direct Link - Blaxhall, P.C. and K.W. Daisley, 1973. Routine haematological methods for use with fish blood. J. Fish Biol., 5: 771-781.
CrossRefDirect Link - Martin, G.G. and L.B. Graves, 1985. Fine structure and classification of shrimp hemocytes. J. Morphol., 185: 339-348.
CrossRefDirect Link - Caldari-Torres, C., W. Banta-Long, A. Bruss, E. Choi and H. Fiegel et al., 2018. Hemolymph glucose levels as a measure of crayfish stress: A methodology using a human glucometer. FASEB J., Vol. 32.
CrossRefDirect Link - Takashima, F. and T. Hibiya, 1995. An Atlas of Fish Histology: Normal and Pathological Features. 2nd Edn., Kondansha Ltd., Tokyo, ISBN-13: 9781560814122, Pages: 195.
Direct Link - Rahman, M.A. and M.S. Islam, 2013. Antioxidant, antibacterial and cytotoxic effects of the phytochemicals of whole Leucas aspera extract. Asian Pac. J. Trop. Biomed., 3: 273-279.
CrossRefDirect Link - Wang, S.H. and J.C. Chen, 2005. The protective effect of chitin and chitosan against Vibrio alginolyticus in white shrimp Litopenaeus vannamei. Fish Shellfish Immun., 19: 191-204.
CrossRef - Maftuch, E. Prasetio, A. Sudianto, M. Rozik and R. Nurdiyani et al., 2013. Improvement of innate immune responses and defense activity in tiger shrimp (Penaeus monodon Fab.) by intramuscular administration of the outer membrane protein Vibrio alginolyticus. SpringerPlus, Vol. 2.
CrossRefDirect Link - Cuzon, G., A.L. Lawrence, G. Gaxiola, C. Rosas and J. Guillaume, 2004. Nutrition of Litopenaeus vannamei reared in tanks or in ponds. Aquaculture, 235: 513-551.
Direct Link - Citarasu, T., G. Immanuel and M.P. Marian, 1998. Effect of feeding Artemia enriched with stresstol and cod liver oil on growth and stress resistance in the Indian white shrimp Penaeus indicus post larvae. Asian Fish. Sci., 12: 65-75.
Direct Link - Nur, I. and Yusnaini, 2018. Parasites and histopathology of infected spiny lobster Panulirus spp. cultured in outer of Kendari Bay, Indonesia. AACL Bioflux, 11: 108-117.
Direct Link - Huang, H.H., X.L. Liu, J.H. Xiang and P. Wang, 2013. Immune response of Litopenaeus vannamei after infection with Vibrio harveyi. Aquaculture, 406: 115-120.
CrossRefDirect Link - Perez-Cano, F.J. and M. Castell, 2016. Flavonoids, inflammation and immune system. Nutrients, Vol. 8.
CrossRefDirect Link - Widodo, A. and R. Pratiwi, 2018. Phytochemical screening, total flavonoid, antioxidant activity, and toxicity of ethanol extract Cleome gynandra L. herb. J. Islamic Pharm., 3: 41-50.
Direct Link - Dulciana, S., J.A.B. Tenório, I.V. Bastos, F.D.S. Mendonca, J.E. Neto, T.G. da Silva and C.S. Ramos, 2016. Chemical and biological studies of β-carotene after exposure to Cannabis sativa smoke. Toxicol. Rep., 3: 516-522.
CrossRefDirect Link - Andayani, S., M. Fadjar and M.F. Rahman, 2018. Isolation and identification of jellyfish alkaloid (Bougainvillia sp) as immunostimulant to profile of protein and fagosite activity of tiger grouper (Epinephelus fuscoguttatus). Aquacult. Indonesiana, 18: 67-71.
Direct Link - Pereira, L., 2018. Biological and therapeutic properties of the seaweed polysaccharides. Int. Biol. Rev., Vol. 2.
Direct Link - Yepiz-Plascencia, G., T.G. Galvan, F. Vargas-Albores and M. García-Bañuelos, 2000. Synthesis of hemolymph high-density lipoprotein β-glucan binding protein by Penaeus vannamei shrimp hepatopancreas. Mar. Biotechnol., 2: 485-492.
CrossRefDirect Link - Arizo, M.A., R.A. Beroncal, W.A. Chua, J.J. Lim, K.C. Rogando and M.B. Maningas, 2016. Immune response of Macrobrachium rosenbergii immersed in aqueous extract of Gracilaria edulis challenged with white spot syndrome virus. Aquacult. Aquarium Conserv. Legisl., 9: 215-226.
Direct Link