
Yue Chen Zhu
Department of Biology, Center for Muscle Biology, University of Kentucky, Lexington, KY 40506, USA
Robin Lewis Cooper
Department of Biology, Center for Muscle Biology, University of Kentucky, Lexington, KY 40506, USA
LiveDNA: 1.523
International Journal of Zoological Research
Year: 2018 | Volume: 14 | Issue: 2 | Page No.: 49-60







ABSTRACT
This review emphasizes how ectothermic animals respond physiologically to environmental temperature changes and cold stress. Generally ectothermic animals cannot generate heat to maintain their body temperature but they can still survive in various temperature zones. Various organisms have been analyzed anatomically and physiologically in the attempt to understand the underlying mechanisms of this temperature adaptation. A few of the changes in the metabolic and molecular function of proteins are discussed. The potential of how hormonal modulation of cardiac and nervous systems which influences responses during exposure cold exposure is also addressed. The focus of this review is on the effect of cold exposure on synaptic transmission and cardiac function in crustaceans and insects.
PDF Abstract XML References Citation
How to cite this article
Yue Chen Zhu and Robin Lewis Cooper, 2018. Cold Exposure Effects on Cardiac Function and Synaptic Transmission at the Neuromuscular Junction in Invertebrates. International Journal of Zoological Research, 14: 49-60.
DOI: 10.3923/ijzr.2018.49.60
URL: https://scialert.net/abstract/?doi=ijzr.2018.49.60
DOI: 10.3923/ijzr.2018.49.60
URL: https://scialert.net/abstract/?doi=ijzr.2018.49.60
INTRODUCTION
Over the past 3.8 billion years, animals have evolved two mechanisms to contend with temperature fluctuations, ectothermy and endothermy. Endothermic animals maintain their core body temperatures independently of ambient temperature changes. In contrast, ectothermic animals undergo body temperature fluctuations in response to ambient temperature changes. However, they can also modify their behavior to regulate their body temperature. Both ectotherms and endotherms have evolved unique physiological processes that ensure their survival under various temperature conditions. Their physiological mechanisms for surviving both high and low environmental temperatures are both unique and similar in certain respects. The arctic fish Myoxocephalus scorpioides can survive low temperatures (-2-+8°C)1, whereas the tropical fish Poecilia reticulate prefers much warmer waters (25.5-27.8°C)2. Neither fish could endure the native environment of the other for any length of time. Therefore, when one evaluates the effects of warm or cold environments on an organism, the normal survival temperature for that species must be considered. The significance of this review is to compile and summarize the findings related to the effects of cold exposure and present the published findings primarily on cardiac and synaptic transmission at the neuromuscular junction in model invertebrate species (Drosophila and crayfish).
To adjust to low temperatures, endotherms either leave or stay in the environment. Therefore, these animals are either hibernators or migrators. Ectotherms subjected to overwintering may hibernate, migrate or endure freezing. For ectothermic animals, temperature affects their longevity as well as their survival. In certain cases, life span is correlated with metabolic rate. Therefore, it may vary with temperature. In general, ectotherms have relatively low metabolic rates in cold environments. Lower ambient temperatures prolong Drosophila life span3-5. For ectotherms, a major advantage of enduring decreasing temperature is that it enables them to wait for environmental conditions conducive to development. They lower their metabolic rates in response to the cold in order to conserve energy that would otherwise be expended on trying to maintain a constant core body temperature. Nevertheless, there are several ecological disadvantages to this approach, such as slowing of movement and a reduction in the ability to escape predators.
CELLULAR TEMPERATURE SENSING
Certain types of cells have their own sensors and response mechanisms to temperature reductions. These responses may involve transcription, translation, protein refolding and membrane reconstruction. These may occur at various times and cooperate additively or even synergistically6. Under cold stress, DNA acts topologically as a thermosensor. First, cold shock increases negative DNA supercoiling. Second, low temperatures enhance DNA bending and improves its affinity for RNA polymerase. Third, low-temperature conditions may increase the binding affinity of histone-like nucleoid structuring proteins (H-NS), which silence nucleoid proteins6-7.
RNA may also be effective as a post-transcriptional intracellular thermosensor, which undergoes cis- and trans- conformational changes. The cis-acting RNA thermometer consists of the Shine-Dalgarno (SD) sequence and the AUG start codon in the 5 UTR of mRNA, which is base-paired at low temperature. When the temperature rises, this structure begins to melt, binds to a ribosome and initiates translation8. Expression of the cold shock protein may be controlled by secondary structures in the 5 UTR. The trans-acting RNA thermometer mechanism consists of sRNA forming a stable secondary structure that melts at higher temperatures. When the temperature rises, the unwound sRNA pairs with the anti-SD sequence in the 5 UTR region of mRNA and exposed a translation-initiating region9.
Proteins can also work as thermosensors because their structures may be altered by changes in temperature. Alterations in temperature alters protein conformations and possibly disassembles dimers10. These protein sensors include transcriptional regulators, chemosensory proteins, chaperones and proteases11. RheA is a specific repressor of the small heat shock gene hsp18. It binds to the hsp18 promoter only at low temperatures. At high temperatures, then, the RheA repressor protein conformation changes and it no longer binds to the hsp18 promoter region. Therefore, hsp18 transcription can be initiated12.
The lipid components of the membrane change in response to temperature fluctuations. These changes may be gradual because lipid component biosynthesis and incorporation into the lipid bilayer are complex processes. Alterations in lipid composition in response to temperature changes have been extensively studied in bacteria. Cells use various mechanisms to maintain membrane fluidity. Decreasing temperature increases the proportions of unsaturated fatty acids with low melting points and high flexibility like cis-vaccenic acid. Its synthesis from palmitoleic acid is mediated by the cold shock-activated enzyme β-ketoacyl-acyl carrier protein (ACP) synthase II. In certain bacteria, desaturases play important roles in altering the degree of fatty acid saturation in membrane phospholipids13. Decreases in temperature reduce the average chain length of fatty acids14. To the best of our knowledge, changes in membrane lipid content in response to cold exposure have not yet been fully examined in crustaceans.
At the cellular level, temperature reductions can decrease membrane fluidity which, in turn, affects membrane associated cellular functions like active transport and protein secretion. Low temperature also alters nucleic acid structures and reduces ribosome function, which lowers translation and protein folding efficiency. Cold shock proteins are indicators of cold stress in cells. Cold shock proteins may help cells survive suboptimal temperatures. They contain RNA-binding domain subgroups known as cold shock domains. These proteins are evolutionarily conserved in bacteria, plants and animals. They function as RNA chaperones and facilitate transcription and translation15. For example CspA is the main cold shock protein in E. coli. In other organisms, its homolog serves as RNA chaperones. CspA melts secondary structures in nucleic acids and facilitates transcription and translation at low temperature. CspA expression is initially and strongly induced by cold shock. It binds to ribosomes and converts them to cold-adapted ribosomes, which translate non-cold–shock mRNAs15.
MECHANISMS TO SURVIVE OVERWINTERING
Certain insects simply migrate in the attempt to avoid a cold environment. For example, each year the monarch butterfly (Danaus plexippus) migrates from Canada and the northern United States to the southern United States and northern Mexico16-17. However, non-migratory insects avoid cold temperatures either by freeze avoidance or by freeze tolerance. In freeze avoidance, the insect limits internal ice formation to prevent lethal freezing. In freeze tolerance, the insect endures subzero temperatures with antifreeze proteins and/or by raising intracellular osmolality to limit ice crystallization in the intercellular spaces.
Many insect species also overwinter in a diapause stage. Diapause is a period of suspended development during unfavorable environmental conditions. Before entering diapause, the insect activates several mechanisms to enable it to cope with the cold external environment. In this process, it may synthesize large quantities of cryo protectants like glycerol or glucose. Its membrane lipid composition may also change. Several protein chaperones resembling heat shock proteins may also be produced. This adaptation is known to occur in the gall fly (Eurosta solidaginis)18.
Other organisms adjust to low temperatures by undergoing rapid cold hardening (RCH). This quick response to cold enables animals to survive longer periods of low temperature19. This mechanism has been extensively studied in fruit flies and flesh flies20. This process involves the rapid accumulation of cryoprotectants like sorbitol and/or glycerol. These chemicals stabilize both cellular and organellar membrane proteins. Sugars like trehalose and amino acids such as proline function as cryoprotectants21-23.
Cold hardening also occurs in the tiny Antarctic nematode (roundworm) Panagrolaimus sp. DAW1. This organism is indigenous to McMurdo Sound, Antarctica. It endures freezing via adaptive cryoprotective dehydration, which eliminates all body water and prevents intracellular freezing24. Researchers investigated nematode gene expression patterns and identified the biochemical pathways that remained active while the nematodes were frozen. Genetic expression analysis of this species in the frozen state showed that it strongly induced the genes and proteases encoding cuticle formation. This study highlighted the genes enabling the organism to endure extreme low-temperature stress. This unique study was the first of its kind to elucidate a rare adaptation in animals, which helped improve the understanding of long-term cold exposure survival strategies. The various mechanisms of seasonal- and rapid cold hardening have been reviewed by Teets and Denlinger20.
EFFECTS OF COLD ON CRAYFISH AND DROSOPHILA
Various crayfish and Drosophila species, as well as other animals naturally occur in different temperature ranges. This review concentrated on the physiological responses to acute and chronic cold in two model invertebrates, namely, the arthropods Procambarus clarkii and Drosophila melanogaster. These species are highlighted for study because they are common, easily analyzed and have practical economic applications in the food and pharmaceutical industries. They are naturally exposed to temperatures in the range25-26 of 15-30°C. These temperature conditions are also readily simulated in a laboratory.
One must consider the behavioral reactions of these two species that enable them to avoid cold and hot environments. Procambarus clarkia prefers27 21.8-25.9°C, whereas Drosophila melanogaster28 prefers~18°C. P. clarki and D. melanogaster significantly differ in terms of their relative responses to temperature change. Moreover, these reactions vary with developmental stage. Both juvenile and adult crayfish swim (tail flip) or walk towards a new location in the water. Both forms dig burrows but adult crayfish are comparatively stronger and can dig deeper burrows through harder substrates. Furthermore, a larger crayfish can cover a greater distance in a shorter time than a smaller one. Since it has a relatively larger surface-to-volume ratio, a juvenile is more rapidly affected internally by water temperature change than an adult. The D. melanogaster adults can fly to various open locations and hide in barns, houses, leaf litter, tree crevices and underground. The larval forms move far more slowly than the adults and tend to seek solid media such as dirt, detritus, fruit and dead animal surfaces or body cavities. However, the temperature of these media may substantially differ from that of the external environment. Therefore, the ambient air or water temperature or wind chill factor may not, in fact, reflect the true physiological condition of D. melanogaster or crayfish. Insects that normally do not survive cold exposures may suddenly emerge after a cold snap, since they could be protected in certain sheltered locations where temperatures are actually somewhat higher than the external environmental temperature. Insects may be caught in a jet stream or wind and be transported thousands of kilometers to new locations29. Water in ship ballast and fruit or other cargo can transport insects and crustaceans to foreign environments where seasonal temperatures differ from that in their native habitats. Once established in their new locations, these species may become invasive30.
It is not yet fully understood how the internal organs of ectotherms continue to function during and after short or long-term cold exposures and are not significantly damaged thereafter. These phenomena merit further investigation. A more thorough understanding of how animals like invertebrates endure short and long-term cold exposure may guide researchers in the application of human cryotherapies such as cold-induced coma, injured tissue icing, nerve block and organ preservation. Previous studies have investigated larval Drosophila heart function in response to low temperatures. Moreover, the effects of heart-stimulating compounds have been analyzed as treatments for cold stress31-32. Serotonin (5-HT), acetylcholine (Ach), octopamine (OA), dopamine (DA) and combinations of these affect larval hearts by modulating ion fluxes. Light activation of transgenically expressed channel rhodopsin proteins under cold conditions has also been investigated32.
Intact larvae were exposed to cold for 10 days or acutely and then dissected to provide access to the bathing saline containing modulators to address the effect on heart rate. The modulators increased heart rate at higher and lower temperatures. Heart rate declined in room temperature (20-21°C)-acclimated flies subjected to low temperatures (10°C). Heart rate also rose at room temperature following modulator (octopamine, acetylcholine, dopamine and 5-HT) treatment. However, heart rate did not increase in cold acclimated larvae exposed to octopamine, acetylcholine or dopamine but did increase when exposed to 5-HT. This study was the first to showed the modulators have different effects on D. melanogaster heart rate at various temperatures. Pharmacological and genetic analyses have been used to investigate the receptor subtypes of 5-HT, dopamine and acetylcholine in larvae at room temperature33-37. However, the effects of cold on these subtypes have not yet been studied. Receptor subtype expression may be differentially regulated at various temperatures to enable the animal to acclimate. It was also found that 5-HT and octopamine levels declined in the hemolymph of larvae acclimated to the cold. This discovery contradicted a hypothesis that the levels of these modulators would, in fact, rise in response to low temperature exposure. Using optogenetics to excite larval cardiac myocytes demonstrated that heart muscle contraction could be accelerated at low temperatures if the myocytes were depolarized. Therefore, it was suggested that endogenous pacemaker activity was a limiting factor in reducing heart rate in response to cold exposure. To date, there have been no reports on electrical event measurements in cold-exposed larval hearts. For this reason, it was unknown whether the heart was pacing but the myocytes limited its ability to contract in the cold. It was also shown that increasing Ca2+ in the bathing medium increased the heart rate after excitation by inducing channel rhodopsin protein expression. The increases in ion flux into the myocytes limited heart rate. Therefore, under low temperatures, the ion flux regulating pacemaker activity is reduced31-32.
Effects of cold exposure on insect nervous systems: Low temperatures can alter osmolality and ion composition in insect neurons. The brain and large isolated nerves are protected by the blood-brain barrier from direct solute exchange in the hemolymph. Therefore, the neuronal microenvironment differs from that of the hemolymph. However, ionic changes in the hemolymph or culture media can rapidly affect neuronal function. Alterations in the relative Na+, K+ and Ca2+ concentrations in the extracellular fluid around the neurons can potentiate cold exposure effects38.
Understanding the mechanism by which temperature fluctuations influence nervous system function may facilitate effective treatment for chill coma and injury in mammals as well as invertebrates. Chilling was associated with a rise in extracellular K+ in locust nervous systems39. Increases in K+ levels depolarize cells and alter their function. Circadian rhythms and temperature cycles also influence neuronal activity. Temperature fluctuations reset circadian clocks by affecting dorsal and lateral posterior neurons in D. melanogaster40. Synapses of the photoreceptor terminals in the house fly (Musca domestica) are affected by temperature variations41. Photoreceptor terminal synapses can disappear within 2-3 h after cold shock and regenerate 20 h later. Cold reversibly induces mitochondrial hypertrophy in epithelial glial cells.
Temperature effects in drosophila: The thermal environment influences ectotherm development rates. In a recent study, Drosophila melanogaster matured from larvae to adults42 in 7.5 d at 29.5°C and in 49th d at 12°C. Gradual cold exposure in insects allows various metabolic processes to continue functioning well enough to help the organism survive and develop. In Chymomyza costata, sugars, polyols and free amino acids are used for long-term cold acclimation. Trehalose and glucose are energy stores and osmoprotectants against freezing. Proline is used for cold conditioning in the beetle Alphitobius diaperinus and Drosophila melanogaster43. Increases in glycerol levels are also correlated with cold acclimation.
Temperature effects in crustaceans: Overwintering may trigger behavioral and physiological changes in crustaceans. In slow, progressive cold exposures, crustaceans, such as crayfish, burrow in the attempt to escape low temperatures. Nevertheless, this strategy may actually subject the animals to even deeper cold. Various ecological factors associated with cold can impact crayfish behavior, including a reduction in food availability, thermoclines caused by melting snow and ice and alterations in diet and predation. The lower wintertime temperatures may reduce physical activity in crayfish, which, in turn, can cause losses of skeletal muscle mass and neuronal function. Cold-induced inactivity is a confounding factor and experimentally difficult to control. It is, therefore, challenging to infer the physiological mechanisms accounting for differential responses to cold exposure by comparing only blood or hemolymph-borne factors or gene or protein expression without directly measuring physiological function. Low temperature (10°C) tolerances were compared between the prawn Macrobrachium rosenbergii, which is indigenous to warm (26-32°C) waters (CABI, 2017) and the crayfish Procambarus clarkii. They were maintained at 21°C, then rapidly chilled to 5°C. The crayfish survived, whereas the prawns died within 2 h. They were also conditioned under gradual chronic cold exposure. All prawns died at 11°C by the second week but the crayfish survived into the 3rd week at44 5°C. All crayfish remained responsive to environmental stimuli at 5°C. Therefore, their nervous systems (sensory, CNS and motor neurons), neuromuscular junctions and muscles were functional under both acute and chronic exposures to 5°C.
The red swamp crayfish Procambarus clarkia is indigenous to the southern United States and northern Mexico. The range of this species has expanded within recent decades. It can now be found from the Great Lakes to the Scandinavian Peninsula. This species has been used as a neuroscience model and for food production. Consequently, it has been transported and naturalized in many different parts of the world beyond its native territory. It also adapts very well to being shipped and housed in laboratories and various outdoor environments.
The stark contrast in the abilities of these two crustacean species to survive cold exposure may be explained by several cellular, biophysical and biochemical factors. Cells must be able to perceive temperature to respond to it. This sensory mechanism may occur at the DNA, RNA and/or protein levels. Therefore, cellular temperature change response may involve transcription, translation, protein conformational changes and membrane lipid bilayer reconstruction.
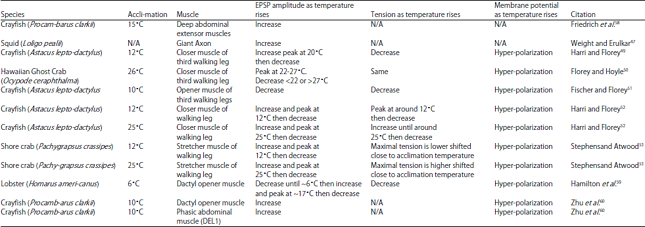
Effect of cold on synaptic transmission in crayfish: Synaptic transmission is the critical step in functional neuronal behavior. In endotherms, body temperature remains at a constant physiological temperature. Therefore, synaptic transmission is not significantly affected by environmental temperature changes. However, it is as yet unknown how synaptic transmission in ectotherms remains functional when challenged by low ambient temperatures. Earlier studies reported that temperature altered synaptic transmission in the giant axon of the squid, an ectotherm45. Low temperatures reduced the Ca2+ current in the presynaptic terminal. Llinas et al.46 reported two main effects of temperature on postsynaptic response. A reduction from 15-5°C increased synaptic delay and decreased the peak amplitude of the excitatory postsynaptic potential (EPSP) in the giant axon of squid. Although, the Ca2+ current in the presynaptic terminal reacts to temperature change, it did not induce changes in EPSP amplitude in the postsynaptic terminal46. A study by Weight and Erulkar47 also demonstrated the EPSP amplitudes increased with temperature in the squid synapses. However, this report did not indicate the initial acclimated temperature of the squid.
Electrophysiological studies of the neuromuscular junction (NMJ) of crustaceans have elucidated the effects of temperature change on synaptic transmission in ectotherms. In crayfish muscle fiber as temperature increases synaptic delay decreases and EPSP amplitude increases48. The EPSP in the closer muscle of the crayfish Astacus leptodactylus acclimated to 12°C increased in amplitude as the temperature rose to 20°C. It then declined as the temperature ascended further. However, closer muscle tension increased with falling temperature49. This response is paradoxical since EPSP amplitudes tend to shrink as the temperature drops. The closer muscle of the Hawaiian ghost crab Ocypode ceratophthalma acclimated50 to 26-27.5°C had maximal EPSP amplitude and muscle tension between 22-28°C. However, both parameters declined as the temperature fell below 22°C or rose above 27°C. Fischer and Florey51 demonstrated that the tension of the opener muscle of the crayfish Astacus leptodactylus acclimated to 10°C increased as temperature decreased. The membrane potential become more negative as the temperature increased from 5-25°C. Unlike other crayfish species, in Astacus leptodactylus, the EPSP amplitude increased as the temperature decreased48. Harri and Florey52 examined the effects of acclimatization to 12 and 25°C in the crayfish Astacus leptodactylus. The EPSP amplitudes were maximal at the acclimated temperatures and the muscle tension shifted towards them. These findings imply that in this particular crayfish species, both synaptic function and muscle contraction are plastic and adjust to the acclimated temperatures of the whole animal. However, this physiological acclimatization to cold does not occur in other crustaceans like prawn that are adapted to warmer waters.
Stephens and Atwood53 acclimated the shore crab Pachygrapsus crassipes to 12 and 21°C and found that their membrane potentials hyperpolarized at higher temperatures. In addition, the EPSP amplitudes initially increased to the maximum value near the acclimation temperatures and decreased thereafter at higher temperatures. In cold-conditioned crabs, the maximal muscle tension was measured at 8-16°C and was found to drop as the temperature increased. In warm-conditioned crabs, the maximal muscle tension was measured at 13-20°C, which also decreased with rising temperature.
Crayfish have long been used as physiological models because they are easy to maintain in relatively simple salt solutions. For NMJ neurotransmission studies, crayfish are valuable because they are easy to dissect and their synaptic transmission rates can be readily quantified54-57. Although, D. melanogaster is better suited as a genetic model, crayfish are large and have individually identifiable motor neurons that facilitate experimental conditions and procedures. The NMJ innervation in crayfish mimics the central synaptic physiology of mammals in that its postsynaptic EPSPs are graded and its neurotransmitter is glutamate. The opener muscle in crayfish is commonly used in NMJ studies because it is easy to dissect and its excitatory nerve is large enough to facilitate intracellular recordings. The nerve termini on the opener muscle support the use of focal macropatch electrodes to evaluate single vesicular events in quantal analysis. Crayfish NMJ remains viable in saline for hours while maintaining consistent synaptic responses. These can be compared with the abundant literature data for low output synapses in the opener NMJ innervation. The high output NMJs of the abdominal extensor have been well characterized in the common Red Swamp Crayfish but the effects of temperature change on alterations in synaptic transmission have not yet been determined.
A synopsis of the various studies on the effects of temperature on crustaceans and squid synaptic transmission is presented in Table 1.
POTENTIAL MECHANISMS IN ALTERING SYNAPTIC TRANSMISSION
Synaptic efficacy is correlated with vesicle fusion. The number of quantal events in crayfish NMJs varies with temperature61. This is also the case for rodents. In the rodent CNS, the correlation between vesicle pool depletion and the speed of endocytosis relative to that of exocytosis is comparatively weak at lower temperatures and relatively strong at higher temperatures62. Both the vesicle recycling rate and the type of exocytosis changed with temperature variation. At 12°C, kiss-and-run endocytosis accounted for <35% of the vesicle fusion. At 28°C, however, this percentage dropped by half 63. Rapid cold shock from 20°C to <4°C, over a few seconds, stuns the nervous system. It results in a lack of a cardiac response to tactile stimulation and silencing of EMG activity in the closer muscles of the chela64. Gradual cold exposure (from 20-10°C) over a 2 week period dampened the excitatory synaptic responses in the walking leg opener muscle of the crayfish Procambarus clarkii. The stimulation frequency that normally evokes a response (10 stimuli at 40 Hz) barely produce any measurable EPSP after the tenth stimulus. Nevertheless, by increasing the stimulation frequency to 60-80 Hz or by adding serotonin (100 nM) at 40 Hz, EPSP responses were observed60. The motor nerve in crustaceans corresponds to the tonic or phasic motor neurons in vertebrates because these nerves are associated with phenotypic muscles. Phasic motor neurons innervate fast-twitch muscles and produce large EPSPs. In crayfish, the abdominal fast-twitch muscle is mainly responsible for tail flips for escape. The opener muscles in the walking leg and chelipeds of crayfish are innervated with tonic-like motor nerves. However, these innervations differ locally and are associated with variations in muscle phenotype65.
| Table 1: | Summary of various studies of the effects of temperature on synaptic transmission |
 | |
Tonic NMJs have low mean quantal content but facilitate with increases in stimulation frequency and are generally fatigue-resistant. They have few primed and docked synaptic vesicles but many reserve vesicles in the presynaptic terminal. In contrast, phasic NMJs produce large EPSPs and fatigue faster and sooner than tonic NMJs. Tonic NMJs have comparatively more primed and docked synaptic vesicles in their phasic neuron terminals and few reserve vesicles. Therefore, relatively more vesicles fuse at phasic NMJs and cannot recycle fast enough to contend with high-frequency stimulation. Consequently, the EPSP amplitudes decrease and the phasic motor units undergo fatigue66-70.
Cold exposure may alter synaptic transmission by affecting motor nerve recruitment, electrical conduction, presynaptic terminal depolarization, Ca2+ disposition, vesicle dynamics and the postsynaptic response of glutamate receptors in depolarizing muscle fibers. Muscle contraction and force development also determine the net output of the motor unit. The motor unit and the muscle may offset the effects of cold by modulating synaptic responses and muscle function. This modulation may occur as a result of changes in hemolymph ion composition and levels of peptides and hormones like the biogenic amines serotonin and octopamine58,71. Modulators such as serotonin (5-HT), octopamine (OA) and dopamine (DA) have important effects on neuronal, cardiac, GI, ventilatory and skeletal muscle function in crustaceans and insects72-75, review76. DA, 5-HT and OA increase in crab hemolymph during exercise77. The OA also increases in insects during movement78-79. These modulators may serve in stress responses in invertebrates80-81. However, the mechanism by which modulation alters neuromuscular junction transmission in response to temperature variation has not yet been fully elucidated. Little is known about the way temperature alters neuromuscular system sensitivity to endogenous neuromodulatory hormones. In the crayfish Procambarus clarkii, modulation by the neuropeptide DF2 is temperature-dependent58. DF2 may help crayfish compensate for the loss of synapse efficacy caused by chilling. The DF2 increased the EPSP amplitude by 300% at 5°C. At 20°C, it increased the EPSP amplitude by 50%. Hamilton et al.59 demonstrated that 5-HT modulates lobster opener muscle subjected to cold stress. The 5-HT increased EPSP amplitude at 2°C by 30% and the tension by 800%. In contrast, the control muscle developed no tension at 2°C. This study reported that 5-HT increased both the EPSP amplitude and the size of neuronally-evoked relaxations. In addition, 5-HT increased resting muscle tension at 2°C. Therefore, the modulation of evoked contractions by 5-HT is temperature-dependent and resting muscle tension is 5-HT-dependent. However, Hamilton et al.59 did not indicate whether 5-HT increases in the hemolymph in response to cold acclimation.
ROLE OF MODULATORS ON CARDIAC AND SYNAPTIC FUNCTION
Recent studies addressed the ways in which cold modulation influences cardiac and neural function in D. melanogaster and P. clarkii. It was assumed that 5-HT, dopamine and octopamine increase in the hemolymph of cold-acclimated larvae. Earlier reports showed that these compounds alone can increase heart rate33,35,82,83. Heart rates lows dramatically in response to acute cold exposure. Therefore, myocytes may be unable to depolarize rapidly in the cold because of altered ion flux. However, optogenetic studies have shown that heart rate can be placed in the cold32. Hormonal or optogenetic control is more easily evaluated in D. melanogaster larvae than in the adults, since the heart is myogenic in the former. On the contrary, in very late larval stages, the third instar and adults, the heart is neurally innervated84. It was recently shown that the heart rate in larval D. melanogasteris regulated by optogenetically driven hormone release from the CNS36. This discovery provides us with a new approach towards understanding the purpose of modulation in the cardiac function of cold-exposed Drosophila.
Much remains to be learned about the effects of modulators on the physiological functions of NMJs in animals acclimated to low temperatures. Only three significant studies examined the effects of modulators on synaptic function at the NMJs of crustaceans58-60. In lobsters, 5-HT enhances synapse output and muscle tension generation in a temperature-dependent manner59. The lobsters used in this study were already naturally cold-conditioned to ~10°C. Therefore, a temperature drop to 2°C may be the reason there is no significant effect. However, the most robust responses in synaptic transmission and tension were closely correlated with the temperatures to which the lobsters were acclimated. Moreover, 5-HT enhanced synaptic transmission at both cold and warm acclimated temperatures (2-20°C). Nevertheless, neither 5-HT nor any other modulator was measured in the hemolymph of these lobsters at various temperatures. Furthermore, no other reports were found measuring modulator levels in crustaceans subjected to temperature variations. One other study addressed the effects of dopamine and different temperatures on the neural circuits of the stomatogastric nervous system of acclimated lobsters85. This study demonstrated that dopamine increased muscle force between 9 and 15°C and negated temperature dependence in synaptic transmission. However, dopamine levels were not measured in the hemolymph of the lobsters exposed to various temperatures. The effects of modulation on synaptic transmission in ectotherms subjected to temperature changes have not been fully investigated.
The effects of an acute drop in temperature to <4°C were investigated in the commercially important blue crab (Callinectes sapidus), red swamp crayfish (Procambarus clarkii) and whiteleg shrimp (Litopenaeus vannamei). In the crayfish, the serotonin levels were lower in response to cold shock and electric stunning than they were at 20°C. Nevertheless, these differences were not observed in the crabs or shrimp64. Response to cold shock have been assessed to investigate/elucidate the potential of cold shock as a humane method for slaughter of crustaceans64,86.
CONCLUSION AND FUTURE DIRECTIONS
Invertebrates rapidly cooled to temperatures lower than those to which they are acclimated present with synaptic, cardiac, neuromodulatory and hormonal responses, which differ from those observed in animals subjected to rapid cooling. Changes in the biophysical properties of skeletal muscle cells and synaptic transmission at the neuromuscular junction were analyzed because the cells in these tissues are large and their responses are easily recorded. Recently, there has been a renewed interest in the mechanism by which modulators enable invertebrate tissues to acclimate to cold exposure.
To explain the findings obtained regarding the effects of cold exposure on Drosophila hearts, it should be determined whether the modulator receptor subtypes in the myocytes are differentially expressed at various temperatures. Only a few of the 30 known modulators in invertebrate hemolymph have been investigated so far. The effects of other biogenic amines and peptides on larval heart function should be investigated. It should also be established whether their concentrations in the hemolymph vary in response to environmental temperature change. Certain studies demonstrated that insect tissues differ in their Ca2+ disposition under cold stress. However, these analyses have not yet been applied to cardiac tissues. Therefore, it remains to be seen whether myocytes acclimate to the cold by increasing resting or active Ca2+ fluxes. It would also be useful to determine whether acute cold exposure in crayfish reduces Ca2+ influx after its release is evoked and whether it prolongs Ca2+ residence in the nerve terminal. This information may explain the mechanism behind the observed alteration in synaptic transmission in response to low temperatures. Nevertheless, impaired vesicle fusion or intracellular Ca2+ perception may also account for cold-induced reduction of synaptic transmission. These mechanisms might be elucidated by quantal analysis with electrophysiology and by imaging using vesicle-associated vital dyes such as FM1-43. It is as yet unknown whether skeletal muscles change in terms of their ability to generate force in response to acute cold or gradual cooling. It is also uncertain how rapid or slow chilling combined with modulators like 5-HT and octopamine differ in terms of their effects on low- and high-output NMJs.
ACKNOWLEDGMENT
We thank Ms. Elena Piana (Sea Farms Limited, Redditch, Worcestershire, United Kingdom) for editorial suggestions. Funding provided by personal funds (RLC).
REFERENCES
- Ragland, S.S. and R.S. Sohal, 1975. Ambient temperature, physical activity and aging in the housefly, Musca domestica. Exp. Gerontol., 10: 279-289.
CrossRefPubMedDirect Link - Speakman, J.R., 2005. Body size, energy metabolism and lifespan. J. Exp. Biol., 208: 1717-1730.
PubMedDirect Link - Klinkert, B. and F. Narberhaus, 2009. Microbial thermosensors. Cell. Mol. Life Sci., 66: 2661-2676.
CrossRefPubMedDirect Link - Phadtare, S. and K. Severinov, 2010. RNA remodeling and gene regulation by cold shock proteins. RNA Biol., 7: 788-795.
CrossRefPubMedDirect Link - Narberhaus, F., T. Waldminghaus and S. Chowdhury, 2006. RNA thermometers. FEMS Microbiol. Rev., 30: 3-16.
CrossRefPubMedDirect Link - Lybecker, M.C. and D.S. Samuels, 2007. Temperature-induced regulation of RpoS by a small RNA in Borrelia burgdorferi. Mol. Microbiol., 64: 1075-1089.
CrossRefPubMedDirect Link - Hurme, R., K.D. Berndt, S.J. Normark and M. Rhen, 1997. A proteinaceous gene regulatory thermometer in Salmonella. Cell, 90: 55-64.
CrossRefPubMedDirect Link - Winter, J. and U. Jakob, 2004. Beyond transcription-new mechanisms for the regulation of molecular chaperones. Crit. Rev. Biochem. Mol. Biol., 39: 297-317.
CrossRefPubMedDirect Link - Servant, P. and P. Mazodier, 2001. Negative regulation of the heat shock response in Streptomyces. Arch. Microbiol., 176: 237-242.
CrossRefPubMedDirect Link - Aguilar, P.S., J.E. Cronan Jr. and D. de Mendoza, 1998. A Bacillus subtilis gene induced by cold shock encodes a membrane phospholipid desaturase. J. Bacteriol., 180: 2194-2200.
Direct Link - Suutari, M., A. Rintamaki and S. Laakso, 1997. Membrane phospholipids in temperature adaptation of Candida utilis: Alterations in fatty acid chain length and unsaturation. J. Lipid. Res., 38: 790-794.
PubMedDirect Link - Jiang, W., Y. Hou and M. Inouye, 1997. CspA, the major cold-shock protein of Escherichia coli, is an RNA chaperone. J. Biol. Chem., 272: 196-202.
CrossRefDirect Link - Altizer, S., K.A. Hobson, A.K. Davis, J.C. de Roode and L.I. Wassenaar, 2015. Do healthy monarchs migrate farther? Tracking natal origins of parasitized vs. uninfected monarch butterflies overwintering in Mexico. PLoS One, Vol. 10, No. 11.
CrossRefDirect Link - Lee, Jr.R.E., R.A. Dommel, K.H. Joplin and D.L. Denlinger, 1995. Cryobiology of the freeze-tolerant gall fly Eurosta solidaginis: Overwintering energetics and heat shock proteins. Clin. Res., 5: 61-67.
Direct Link - Lee, Jr.R.E., C.P. Chen and D.L. Denlinger, 1987. A rapid cold-hardening process in insects. Science, 238: 1415-1417.
CrossRefPubMedDirect Link - Teets, N.M. and D.L. Denlinger, 2013. Physiological mechanisms of seasonal and rapid cold-hardening in insects. Physiol. Entomol., 38: 105-116.
CrossRefDirect Link - Withers, L.A. and P.J. King, 1979. Proline: A novel cryoprotectant for the freeze preservation of cultured cells of Zea mays L. Plant Physiol., 64: 675-678.
PubMedDirect Link - Bhandal, I.S., R.M. Hauptmann and J.M. Widholm, 1985. Trehalose as cryoprotectant for the freeze preservation of carrot and tobacco cells. Plant Physiol., 78: 430-432.
PubMedDirect Link - Tsai, S., G. Chong, P.J. Meng and C. Lin, 2017. Sugars as supplemental cryoprotectants for marine organisms. Rev. Aquacult., 10: 703-715.
CrossRefDirect Link - Thorne, M.A.S., A. Seybold, C. Marshall and D. Wharton, 2017. Molecular snapshot of an intracellular freezing event in an Antarctic nematode. Cryobiology, 75: 117-124.
CrossRefPubMedDirect Link - Maguire, S.E., P.S. Schmidt and A. Sehgal, 2014. Natural populations of Drosophila melanogaster reveal features of an uncharacterized circadian property: The lower temperature limit of rhythmicity. J. Biol. Rhythms, 29: 167-180.
CrossRefPubMedDirect Link - Tattersall, G.J., J.P. Luebbert, O.K. LePine, K.G. Ormerod and A.J. Mercier, 2012. Thermal games in crayfish depend on establishment of social hierarchies. J. Exp. Biol., 215: 1892-1904.
CrossRefPubMedDirect Link - Dillon, M.E., G. Wang, P.A. Garrity and R.B. Huey, 2009. Review: Thermal preference in Drosophila. J. Therm. Biol., 34: 109-119.
CrossRefPubMedDirect Link - Hulme, P.E., 2009. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Applied Ecol., 46: 10-18.
CrossRefDirect Link - Zhu, Y.C., E. Yocom, J. Sifers, H. Uradu and R.L. Cooper, 2016. Modulatory effects on Drosophila larva hearts: Room temperature, acute and chronic cold stress. J. Comp. Physiol. A., 186: 829-841.
CrossRefPubMedDirect Link - Zhu, Y.C., H. Uradu, Z.R. Majeed and R.L. Cooper, 2016. Optogenetic stimulation of Drosophila heart rate at different temperatures and Ca2+ concentrations. Physiol. Rep., Vol. 4, No. 3.
CrossRefDirect Link - Majeed, Z.R., C.D. Nichols and R.L. Cooper, 2013. 5-HT stimulation of heart rate in Drosophila does not act through cAMP as revealed by pharmacogenetics. J. Applied Physiol., 115: 1656-1665.
CrossRefPubMedDirect Link - Majeed, Z.R., A. Stacy and R.L. Cooper, 2014. Pharmacological and genetic identification of serotonin receptor subtypes on Drosophila larval heart and aorta. J. Comp. Physiol. B., 184: 205-219.
CrossRefPubMedDirect Link - Titlow, J.S., J.M. Rufer, K.E. King and R.L. Cooper, 2013. Pharmacological analysis of dopamine modulation in the Drosophila melanogaster larval heart. Physiol. Rep., Vol. 1, No. 2.
CrossRefDirect Link - Malloy, C., J. Sifers, A. Mikos, A. Samadi, A. Omar, C. Hermanns and R.L. Cooper, 2017. Using optogenetics to assess neuroendocrine modulation of heart rate in Drosophila melanogaster larvae. J. Comp. Physiol. A, 203: 791-806.
CrossRefPubMedDirect Link - Malloy, C.A., K. Ritter, J. Robinson, C. English and R.L. Cooper, 2015. Pharmacological identification of cholinergic receptor subtypes on Drosophila melanogaster larval heart. J. Comp. Physiol. B, 186: 45-57.
CrossRefPubMedDirect Link - MacMillan, H.A. and B.J. Sinclair, 2011. The role of the gut in insect chilling injury: Cold-induced disruption of osmoregulation in the fall field cricket, Gryllus pennsylvanicus. J. Exp. Biol., 214: 726-734.
CrossRefPubMedDirect Link - Rodgers, C.I., G.A.B. Armstrong and R.M. Robertson, 2010. Coma in response to environmental stress in the locust: A model for cortical spreading depression. J. Insect. Physiol., 56: 980-990.
CrossRefPubMedDirect Link - Miyasako, Y., Y. Umerzaki and K. Tomioka, 2007. Separate sets of cerebral clock neurons are responsible for light and temperature entrainment of Drosophila circadian locomotor rhythms. J. Biol. Rhythms, 22: 115-126.
CrossRefPubMedDirect Link - Brandstater, J.H. and I.A. Meinetzhagen, 1995. The rapid assembly of synaptic sites in photoreceptor terminals of the fly’s optic lobe recovering from cold shock. Proc. Natl. Acad. Sci. USA., 92: 2677-2681.
PubMedDirect Link - Schou, M.F., T.N. Kristensen, A. Pedersen, B.G. Karlsson, V. Loeschcke and A. Malmendal, 2017. Metabolic and functional characterization of effects of developmental temperature in Drosophila melanogaster. Am. J. Physiol. Regul. Integr. Comp. Physiol., 312: R211-R222.
CrossRefPubMedDirect Link - Kostal, V., P. Simek, H. Zahradnickova, J. Cimlova and T. Stetina, 2012. Conversion of the chill susceptible fruit fly larva (Drosophila melanogaster) to a freeze tolerant organism. Proc. Natl. Acad. Sci. USA., 109: 3270-3274.
CrossRefPubMedDirect Link - Chung, Y.S., R.M. Cooper, J. Graff and R.L. Cooper, 2012. The acute and chronic effect of low temperature on survival, heart rate and neural function in crayfish (Procambarus clarkii) and prawn (Macrobrachium rosenbergii) species. Open J. Mol. Integr. Physiol., 2: 75-86.
CrossRefDirect Link - Katz, B. and R. Miledi, 1965. The effect of temperature on the synaptic delay at the neuromuscular junction. J. Physiol., 181: 656-670.
CrossRefPubMedDirect Link - Weight, F.F. and S.D. Erulkar, 1976. Synaptic transmission and effects of temperature at the squid giant synapse. Nature, 261: 720-722.
CrossRefDirect Link - Charlton, M.P. and H.L. Atwood, 1979. Synaptic transmission: Temperature-sensitivity of calcium entry in presynaptic terminals. Brain Res., 170: 543-546.
CrossRefDirect Link - Harri, M. and E. Florey, 1977. The effects of temperature on a neuromuscular system of the crayfish, Astacus leptodactylus. J. Compa. Physiol., 117: 47-61.
CrossRefDirect Link - Florey, E. and G. Hoyle, 1976. The effects of temperature on a nerve-muscle system of the Hawaiian ghost crab, Ocypode ceratophthalma (Pallas). J. Compa. Physiol. A: Neuroethol. Sens. Neural Behav. Physiol., 110: 51-64.
Direct Link - Fischer, L. and E. Florey, 1987. Octopamine action on the contractile system of crustacean skeletal muscle. Compa. Biochem. Physiol. Part C: Compa. Pharmacol., 88: 335-342.
CrossRefDirect Link - Harri, M. and E. Florey, 1979. The effects of acclimation temperature on a neuromuscular system of the crayfish, Astacus leptodactylus. J. Exp. Biol., 78: 281-293.
Direct Link - Stephens, P.J. and H.L. Atwood, 1982. Thermal acclimation in a crustacean neuromuscular system. J. Exp. Biol., 98: 39-47.
PubMedDirect Link - Dudel, J. and S.W. Kuffier, 1961. The quantal nature of transmission and spontaneous miniature potentials at the crayfish neuromuscular junction. J. Physiol., 155: 514-529.
PubMedDirect Link - Dudel, J. and S.W. Kuffler, 1961. Presynaptic inhibition at the crayfish neuromuscular junction. J. Physiol., 155: 543-562.
PubMedDirect Link - Dudel, J., 1963. Presynaptic inhibition of the excitatory nerve terminal in the neuromuscular junction of the crayfish. Pfluger's Arch. Gesamte Physiol. Menschen Tiere, 277: 537-557.
CrossRefDirect Link - Dudel, J., 1965. The mechanism of presynaptic inhibition at the crayfish neuromuscular junction. Pfluger's Arch. Gesamte Physiol. Menschen Tiere, 284: 66-80.
CrossRefDirect Link - Friedrich, R.W., P.A. Quigley, M. Srivastava, M. Skerrett and A.J. Mercier, 1994. Temperature dependence of synaptic modulation by a FMRFamide-related neuropeptide in crayfish. Neurosci. Lett., 169: 56-58.
CrossRefDirect Link - Hamilton, J.L., C.R. Edwards, S.R. Holt and M.K. Worden, 2007. Temperature dependent modulation of lobster neuromuscular properties by serotonin. J. Exp. Biol., 210: 1025-1035.
CrossRefDirect Link - Zhu, Y.C., L. de Castro and R.L. Cooper, 2018. Effect of temperature change on synaptic transmission at crayfish neuromuscular junctions. Biol. Open, Vol. 7.
CrossRefDirect Link - Johnson, E.W. and A. Wernig, 1971. The binomial nature of transmitter release at the crayfish neuromuscular junction. J. Physiol., 218: 757-767.
CrossRefPubMedDirect Link - Fernandez-Alfonso, T. and T.A. Ryan, 2004. The kinetics of synaptic vesicle pool depletion at CNS synaptic terminals. Neuron, 41: 943-953.
CrossRefDirect Link - Zhang, Z. and M.B. Jackson, 2008. Temperature dependence of fusion kinetics and fusion pores in Ca2+-triggered exocytosis from PC12 cells. J. Gen. Physiol., 131: 117-124.
CrossRefPubMedDirect Link - Weineck, K., A.J. Ray, L. Fleckenstein, M. Medley, N. Dzubuk, E. Piana and R.L. Cooper, 2018. Physiological changes as a measure of crustacean welfare under different standardized stunning techniques: Cooling and electroshock. Animals, Vol. 8, No. 9.
CrossRefDirect Link - Mykles, D.L., S. Medler, A. Koenders and R. Cooper, 2002. Myofibrillar protein isoform expression is correlated with synaptic efficacy in slow fibres of the claw and leg opener muscles of crayfish and lobster. J. Exp. Biol., 205: 513-522.
PubMedDirect Link - Atwood, H.L. and R.L. Cooper, 1995. Functional and structural parallels in crustacean and Drosophila neuromuscular systems. Am. Zool., 35: 556-565.
PubMedDirect Link - Atwood, H.L. and R.L. Cooper, 1996. Synaptic diversity and differentiation: Crustacean neuromuscular junctions. Invert. Neurosci., 1: 291-307.
CrossRefDirect Link - Logsdon, S., A.F.M. Johnstone, K. Viele and R.L. Cooper, 2006. Regulation of synaptic vesicles pools within motor nerve terminals during short-term facilitation and neuromodulation. J. Applied Physiol., 100: 662-671.
CrossRefPubMedDirect Link - Wu, W.H. and R.L. Cooper, 2010. Physiological recordings of high and low output NMJs on the crayfish leg extensor muscle. J. Vis. Exp. Vol. 45.
CrossRef - Wu, W.H. and R.L. Cooper, 2012. The regulation and packaging of synaptic vesicles as related to recruitment within glutamatergic synapses. Neuroscience, 225: 185-198.
CrossRefPubMedDirect Link - Colton, C.K. and A.R. Freeman, 1974. Dual response of lobster muscle fibers to L-glutamate. J. Comp. Biochem. Physiol. C, 51: 275-284.
PubMedDirect Link - Strawn, J.R., W.S. Neckameyer and R.L. Cooper, 2000. The effects of 5-HT on sensory, central and motor neurons driving the abdominal superficial flexor muscles in the crayfish. Comp. Biochem. Physiol. B: Biochem. Mol. Biol., 127: 533-550.
PubMedDirect Link - Listerman, L.R., J. Deskins, H. Bradacs and R.L. Cooper, 2000. Heart rate within male crayfish: Social interactions and effects of 5-HT. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol., 125: 251-263.
CrossRefPubMedDirect Link - Cooper, A.S., B. Leksrisawat, A.J. Mercier, A.B. Gilberts and R.L. Cooper, 2011. Physiological experimentations with the crayfish hindgut. J. Visual. Exp., Vol. 18.
CrossRefDirect Link - Cooper, R.M., H.S. Finucane, M. Adami and R.L. Cooper, 2011. Heart and ventilatory measures in crayfish during copulation. Open J. Mol. Integr. Physiol., 1: 36-42.
CrossRefDirect Link - Shuranova, Z.P., Y.M. Burmistrov, J.R. Strawn and R.L. Cooper, 2006. Evidence for an autonomic nervous system in decapod crustaceans. Int. J. Zool. Res., 2: 242-283.
CrossRefDirect Link - Sneddon, L.U., A.C. Taylor, F.A. Huntingford and D.G. Watson, 2000. Agonistic behaviour and biogenic amines in shore crabs Carcinus maenas. J. Exp. Biol., 203: 537-545.
PubMedDirect Link - Goosey, M.W. and D.J. Candy, 1980. The D-octopamine content of the haemolymph of the locust, Schistocerca Americana gregaria and its elevation during flight. Insect Biochem., 10: 393-397.
CrossRefDirect Link - David, J.C. and J.F. Coulon, 1985. Octopamine in invertebrates and vertebrates. A review. Progress Neurobiol., 24: 141-185.
CrossRefPubMedDirect Link - Page, M.P., W. Hailes and R.L. Cooper, 2007. Modification of the tail flip escape response in crayfish by neuromodulation and behavioral state with and without descending CNS input. Int. J. Zool. Res., 3: 132-144.
CrossRefDirect Link - Even, N., J.M. Devaud and A.B. Barron, 2012. General stress responses in the honey bee. Insects, 3: 1271-1298.
CrossRefPubMedDirect Link - Johnson, E., J. Ringo and H. Dowse, 1997. Modulation of Drosophila heartbeat by neurotransmitters. J. Comp. Physiol. B, 167: 89-97.
CrossRefPubMedDirect Link - Johnson, E., T. Sherry, J. Ringo and H. Dowse, 2002. Modulation of the cardiac pacemaker of Drosophila: Cellular mechanisms. J. Compa. Physiol. B, 172: 227-236.
CrossRefPubMedDirect Link - Johnstone, A.F.M. and R.L. Cooper, 2006. Direct innervation of the Drosophila melanogaster larval aorta. Brain Res., 1083: 159-163.
CrossRefDirect Link - Thuma, J.B., K.H. Hobbs, H.J. Burstein, N.S. Seiter and S.L. Hooper, 2013. Temperature sensitivity of the pyloric neuromuscular system and its modulation by dopamine. PloS One, Vol. 8, No. 6.
CrossRefDirect Link - Diggles, B.K., 2018. Review of some scientific issues related to crustacean welfare. ICES J. Mar. Sci.
CrossRefDirect Link