
Bahaa EL-Deen Wadea El-Aswad
Department of Medical Parasitology, Faculty of Medicine, Menoufia University, Egypt
Amira Fathy Abd El-Atty
Department of Medical Parasitology, Faculty of Medicine, Menoufia University, Egypt
Manal Ahmed El-Melegy
Department of Medical Parasitology, Faculty of Medicine, Menoufia University, Egypt
Raafat Mohamed Shaapan
Department of Zoonotic Diseases, Veterinary Research Division, National Research Centre, Giza, Egypt
LiveDNA: 20.659
Amany Fawzy Atia
Department of Medical Parasitology, Faculty of Medicine, Menoufia University, Egypt
Noha Ahmed El Kaddy
Department of Pathology, Faculty of Medicine, Menoufia University, Egypt
Mohamed Mohamed Abd El-Ghaffar
Department of Medical Parasitology, Faculty of Medicine, Menoufia University, Egypt
International Journal of Zoological Research
Year: 2018 | Volume: 14 | Issue: 1 | Page No.: 21-29







ABSTRACT
Background and Objective: A zoonotic Trypanosoma evansi (T. evansi ) has the widest range and geographical distribution attributed to the mechanical transmission by many vectors. Inducible nitric oxide synthase (iNOS) activation by interleukin-17 (IL-17) were expressed in mice experimentally infected with T. evansi in addition to detection of serum level of IL-17 for better understanding the roles of iNOS and IL-17 in controlling this protozoan infection. Materials and Methods: Infected group (I) mice were injected with 4.5×105 trypomastigotes intraperitoneally, subdivided into I-3, I-5, I-10 and I-20 subgroups (10 mice, each), then sacrificed at 3, 5, 10 and 20 days post infection (p.i.), respectively, whereas control group mice (C) were injected with saline, subdivided into C-3, C-5, C-10 and C-20 subgroups (6 mice, each) and sacrificed along the parallel infected subgroup one. Results: iNOS was immunohistochemically detected with more expression in brain and the IL-17 revealed more in spleen than liver or brain, also its serum level increased significantly with progression of the infection. There was a negative correlation between parasitemia and either iNOS or IL-17. Conclusion: Trypanosoma evansi infection caused increasing both IL-17 and iNOS expression which both were coinciding with a low number of trypomastigotes referring strongly to their roles in limiting the infection.
PDF Abstract XML References Citation
Received: July 12, 2018;
Accepted: July 26, 2018;
Published: October 24, 2018
How to cite this article
Bahaa EL-Deen Wadea El-Aswad, Amira Fathy Abd El-Atty, Manal Ahmed El-Melegy, Raafat Mohamed Shaapan, Amany Fawzy Atia, Noha Ahmed El Kaddy and Mohamed Mohamed Abd El-Ghaffar, 2018. Roles of Inducible Nitric Oxide Synthase and Interleukin-17 in Mice Experimentally Infected with Trypanosoma evansi. International Journal of Zoological Research, 14: 21-29.
DOI: 10.3923/ijzr.2018.21.29
URL: https://scialert.net/abstract/?doi=ijzr.2018.21.29
DOI: 10.3923/ijzr.2018.21.29
URL: https://scialert.net/abstract/?doi=ijzr.2018.21.29
INTRODUCTION
Trypanosoma evansi (T. evansi ) is the most widely distributed pathogenic Trypanosoma species which can affect many animals' species causing a disease known popularly as “Surra” of varying pathological lesions and inflammation in the infected animals1. The flagellate protozoan T. evansi is a, not capable of infecting the invertebrate vector, so transmitted mechanical via very broad vectors, Tabanas sp., Haematobia sp. and Stomoxys sp2. The infection with T. evansi was demonstrated in many animal species (camels, sheep cattle and buffaloes) causing abortion and trans-placental transmitted to the fetus in cattle and buffaloes while, a single case human infection was reported3,4.
The pathogenesis of T. evansi infection depends on the virulence of the strain, host susceptibility, stress and epidemiological conditions. Anemia is the main outcome of infection consequently, fever, edema, enlarged lymph nodes, icterus and emaciation5. There were many suspected cases of human T. evansi infection6, including an Egyptian farmer7, in addition to two other cases that were confirmed by molecular technique, one Indian farmer who had deficiency of apolipoprotein L1 (APOL1)8, while the other one was discovered in Vietnam and she was a previously healthy woman without APOL1 deficiency9.
However, many epidemiological studies (parasitological, serological and molecular techniques) on Trypanosoma evansi infection in camels in Egypt10-12, few reports about T. evansi infection in other animals in Egypt, T. evansi infection in Egyptian water buffaloes calves was proved by hematological and biochemical studies by Hilali et al.13. Also, one suspected human case of T. evansi, positive with ELISA test was reported without isolation and other confirmatory tests 7.
Microscopic examination at by help of 100x oil immersion lens used for the detection of trypomastigote forms of T. evansi in wet films, Giemsa stained blood smears, this method is being specific but less sensitive, principally in detecting parasites during low levels of parasitemia14. Several serological tests such as IFAT, ELISA and the specific card agglutination test (CAT) used for T. evansi mass screening, however, antibodies may be missing due to serological latency15. Polymerase Chain Reaction (PCR) test for T. evansi displayed at low parasitemia was performed with primer sets (TBR1 and TBR2) specific for Trypanosoma satellite DNA regions, positive results showed a single band (164 bp) only for T. evansi16.
Few reports are available regarding the immunological responses against T. evansi infection contrary to other trypanosomes species. Nitric oxide (NO) is synthesized by three iso forms of NO synthase enzymes, endothelial (eNOS), neuronal (nNOS) and inducible (iNOS). Unlike eNOS and nNOS, iNOS is expressed by many cell types when exposed to stimulatory conditions like infection, hypoxia and injury17. It exerts its functions by catalyzing the conversion of L-arginine into nitric oxide (NO)18. The expression of iNOS in tissues can be used as an indirect indicator of NO level. NO is an uncharged free radical which has many physiological effects, such as vasodilatation, lymphocytes proliferation, production of cytokines, in addition to being a neurotransmission mediator19,20. Interleukin (IL) 17 is a pro-inflammatory cytokine which has a remarkable role in mediating the inflammation process21 and it is involved in many autoimmune diseases22. While its role in protecting against T. evansi has not been investigated yet.
This study was carried out to explore the expressions of iNOS and IL-17 by immunohistochemistry method in some organs of the mice experimentally infected with T. evansi, in addition to detection of serum level of IL-17 for better understanding pathogenesis of the acute T. evansi infection and to investigate the roles of iNOS and IL-17 in controlling this protozoan infection.
MATERIALS AND METHODS
This prospective study was carried out to evaluate the role of Nitric Oxide Synthase and Interleukin-17 to control of Trypanosoma evansi in experimentally infected mice, during the period June-December, 2017at the Parasitology Department, Menoufia University, Menoufia, Egypt.
Animal model: Female BALB/c mice (7 weeks old and 18±5 g in weight) were brought to the department of medical parasitology, Faculty of Medicine, Menoufia University. The animals were kept under controlled conditions of temperature (25±2°C) and humidity (70%). They were free to access commercial diet and water.
Experimental design and T. evansi infection: The mice were divided into two groups, group I (infected mice, n = 40) and group C (control uninfected mice, n = 24). Trypomastigotes of T. evansi were obtained from blood of naturally T. evansi infected camel that kept in the Faculty of Veterinary Medicine, Cairo University. The strain was maintained in mice, where their infected blood was used for doing the experimental infection according to procedures described by Desquesnes et al.3. On day 0, each mouse of the group I was inoculated intraperitoneally with 0.25 mL containing 4.5×105 trypomastigotes. The mice of group C were injected with 0.25 mL saline intraperitoneally at the same time of group I and each subgroup was sacrificed along with the corresponding infected mice subgroup.
Animal sacrificing and blood sampling: The mice were anesthetized with ether and blood was collected by heart puncture method as the procedures described by Lewis23, during the planned periods. Sera were separated, divided and kept at -20°C till used.
Estimation of parasitemia: The degree of parasitemia was estimated daily for each infected mouse as the method stated by Joshi et al.8, briefly, a drop of blood was obtained from the tail vein from each mouse to detect the presence of T. evansi using direct and Giemsa stained blood smears. Trypanosoma evansi try promastigotes were counted in each mouse blood sample and the mean of these counts was estimated to express the degree of parasitemia.
Immunohistochemical detection of iNOSin mice tissues: The iNOS enzyme was detected in the mice tissues by using iNOS Polyclonal Antibody (Thermo Fisher Scientific) following the instructions reported by Wilson et al.24 as follows, paraffin-embedded tissue sections (5 μm thickness) were induced as antigen, the specimens were blocked by 3% bovine serum albumin-PBS for 30 min, rinsed in PBS, then, the tissues were probed with anti-iNOS polyclonal primary antibody or without primary antibody (negative control) overnight in a humid chamber. The specimens were washed three times with PBS and peroxidase activity was quenched. After that, the specimens were incubated with the biotinylated rabbit anti-polyvalent secondary antibody (1:500) for 30 min followed by adding streptavidin conjugated to horseradish peroxidase for 30 min, then they were washed three times for 2 min/wash. The prepared 3,3-diaminobenzidine (DAB) chromogen solution was added for 10 min, the specimens were counter stained with Mayer’s hematoxylin. Within each case, 1000 cells were counted, the percentage of positively stained cells was calculated and the expression was categorized into the following grades:
| 0 | = | <5% |
| I | = | 5-25% |
| II | = | 26-51% |
| III | = | 51-75% |
Immunohistochemical detection of IL-17 in mice tissues: IL-17 was detected in the tissues using Interleukin 17A Monoclonal Antibody Kits (Biorbyt) following the manufacturer’s instructions. Briefly, paraffin tissue sections were mounted on slides, endogenous peroxidase activity was blocked with 3% hydrogen peroxide solution for 10 min at room temperature. Heat-induced epitope retrieval procedure was used for antigen retrieval. Rabbit anti-mouse IL-17 primary monoclonal antibody was applied, then the slides were incubated overnight at room temperature. Negative control slides were prepared by adding all the reagents but not the primary antibody. After three washes in PBS, biotinylated goat anti-mouse polyclonal secondary antibody was applied, incubated for 30 min. Streptavidin peroxidase was applied and incubated for 30 min, then the prepared DAB chromogen was added for 10-15 min. After counterstaining with hematoxylin, the slides were washed, dehydrated cleared with xylene and the coverslip was mounted. For each slide, 1000 cells were counted at x 400 magnification and according to the percentage of the IL-17 positive cells, the sections were classified into four grades:
| 0 | = | <5% |
| I | = | 5-25% |
| II | = | 26-51% |
| III | = | 51%25 |
Detection of mouse IL-17 in mice serum: The collected mice sera were serologically assayed to determine IL-17 by using a sandwich enzyme-linked immunosorbent assay (ELISA) according to the method described by Shaapan et al.26, briefly, 96-well flat-bottom plate was coated with 100 μL/well of the capture antibody, washed with PBS/Tween (T) 20 and the nonspecific binding sites were blocked with PBS/T/1% dry milk powder. After washing three times, the serum samples or the recombinant cytokine were added in a volume of 100 μL/well. After 3 times, 100 μL/well of the biotinylated anti-cytokine detection antibody was loaded. After incubation for 1 h with gentle shaking, the plate was washed three times, then, the streptavidin-alkaline phosphatase conjugate was added and the plate was left for 30 min. At last, the plate was washed for five times, the reaction was developed with para-nitrophenyl phosphate (Sigma) and the absorbance at 405 nm was measured using a microplate ELISA reader (Bio-Rad). IL-17 concentration was calculated from the generated standard curve.
Statistical analysis: SPSS computer program (version 12 windows) was used for data analysis. Comparison relative to the infected untreated control group was performed using unpaired Student’s t-test. Data were expressed as Mean±Standard Deviation (SD). Pearson’s correlation coefficient (r) was performed on the paired data obtained by the individual infected cases. The data were considered significant if p<0.05.
Ethical approval: Ethical clearance and approval were obtained from the National Research Centre Ethics Committee.
RESULTS
Parasitemia: Trypanosoma evansi was detected in peripheral blood of all the infected mice starting from 24-48 h p.i. Trypomastigote was spindle-shaped, an extra-erythrocytes organism having a thin posterior end, large undulating membrane, free flagellum, small sub-terminal kinetoplast and central nucleus (Fig. 1a). Parasitemia increased progressively to reach its peak (142.9±37.74 trypomastigotes/HPF), in mice subgroup I-5 the (day 5 p.i.), then it decreased as the infection progressed to record32.9±6.79 trypomastigotes/ HPF in the subgroup I-10 (day 10 p.i.) and in the subgroup I-20 (day 20 p.i.) there were only 0.7±0.82 trypomastigotes/HPF (Fig. 1b). During this study, further peaks of parasitemia were not recorded. The control mice subgroups remained clinically healthy during the whole period of the experiment.
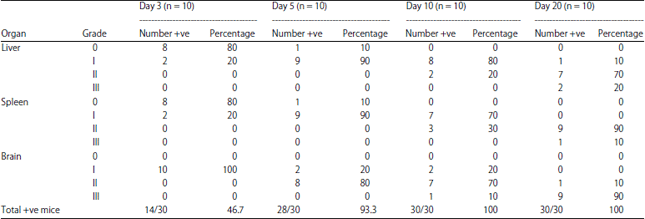
Immunohistochemical detection of iNOS: The tissues of the control mice did not show iNOS enzyme. Brain tissues have expressed the enzyme in a greater extent than liver and spleen along the course of the infection. In subgroup I-3, all brains of infected mice (100%) expressed iNOS in grade I, whereas only 20% of each of livers and spleens expressed the enzyme in grade I. At peak of parasitemia (subgroup I-5), 90% of the livers and spleens showed the enzyme in grade I, while 100% of the brains revealed the enzyme (20% in grade I and 80% in grade II). Regarding subgroup I-10, all brains (20, 70 and 10% in grades I, II and III, respectively), livers (80 and 20% in grades I and II, respectively) and spleens (70 and 30% in grades I and II, respectively) showed the enzyme. At the end of the experiment (subgroup I-20), expression of the enzyme involved all the examined tissues and it extended more extensively, where 90 and 10% of brains expressed the enzyme in grade III and II, respectively. In livers, 10, 70% and 20% of the tissue expressed the enzyme in grade I, II and III, respectively, while, 90 and 10% of spleens expressed the enzyme in grade II and III, respectively.
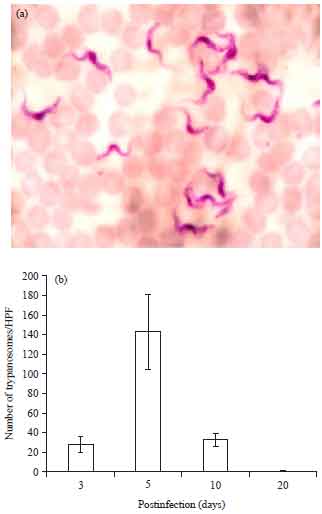 | |
| Fig. 1(a-b): | Parasitemia of T. evansi in infected mice, (a) Trypomastigotes of T. evansi with central nucleus, flagella and undulating membrane in a peripheral blood smear from an infected mice, Giemsa stained (X1000) and (b) parasite number among the infected mice subgroups |
Collectively, the enzyme was expressed in 46.7% (14/30) in subgroup I-3, 93.3% (28/30) in subgroup I-5, whereas it was 100% (30/30) in subgroups I-10 and I-20 (Table 1 and Fig. 2). There was a negative correlation (p<0.01 and r= -0.40) between the expression of the enzyme (percentage of positives cells) and parasitemia (Fig. 3).
Immunohistochemically detection of IL-17: Cells secreting IL-17 were macrophage and lymphocyte-like. The cytokine was expressed largely by spleen, followed by liver, then brain. In subgroup I-3, IL-17 was expressed in grad I in liver (70%), spleen (90%) and brain (10%). In subgroup I-5, 100% spleen (in grade II) and 100% brain (in grade I), 100% of the liver (90% in grade II and 10% in grade I) revealed IL-17. In subgroup I-10, 70% of the liver was positive for the cytokine in grade II and 30% showed it in grade III, 40% of spleens revealed the IL-17 in grade II and 60% was of grade III, while 70 and 30% of the brains expressed it in grade I and grade II, respectively. Finally, 90% of spleens, 70% of livers and only 20% of brains showed the cytokine with grade III in the subgroup I-20. Collectively, IL-17 was expressed in 56.7% (17/30) and 100% (30/30) in the other subgroups (Table 2 and Fig. 4). There was a negative correlation (p<0.05 and r = -0.31) between the expression of IL-17 expressed (% of positives cells) and parasitemia (Fig. 5).
| Table 1: | iNOS expression among different tissues of T. evansi infected mice subgroups |
 | |
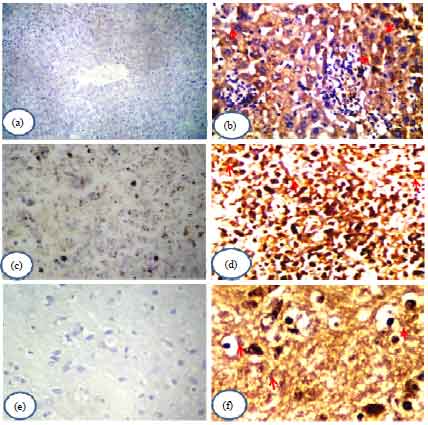 | |
| Fig. 2(a-f): | iNOS expression in the tissues of T. evansi infected mice, (a) Liver of control uninfected mice shows no iNOS expression (x100), (b) Liver of infected mice showing iNOS expressed in grade III (x400) (red head arrows), (c) Spleen of control uninfected mice has no iNOS expression (x200), (d) Spleen of infected mice reveals iNOS expressed in grade III (x400)(red head arrows), (e) Brain of control uninfected mice has no iNOS expression (x400) and (f) Brain of infected mice shows iNOS expressed in grade III (x400)(red head arrows) |
ELISA detection of IL-17 in mice serum: Serum level of IL-17 was significantly (p<0.001) high in all the infected mice subgroups in comparison with the control ones and its level was increasing as the infection progressed reaching its highest level at day 20 p.i. (Fig. 6).
DISCUSSION
In the present study, the peak of T. evansi parasitemia occurred at day 5 p.i., then the parasite level decreased until the end of the experiment and this was similar to other results which studied T. evansi infection in mice27,28. We determined iNOS locally in liver, spleen and brain to examine its distribution in these organs during the course of acute infection, besides it reflected the NO level indirectly because of its very short half-life, detection of NO itself is difficult.
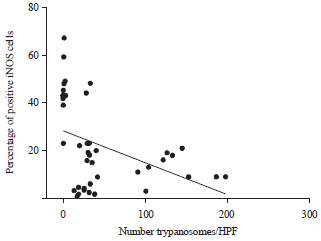 | |
| Fig. 3: | Correlation between percentage of positive iNOS cells and number of T. evansi trypomastigotes in infected mice (p<0.01 and r = -0.40) |
Other researchers detect NO indirectly through measurement of nitrites/nitrates, the circulatory stable end products of NO29 or 3-nitrotyrosine, formed by the reaction of peroxynitrite with the phenol group of tyrosine30.
The brain showed iNOS enzyme in greater level than liver and spleen along the whole course of the infection reaching 90% with grade III in subgroup I-20. The expression of iNOS in liver, spleen and brain of the infected mice subgroups correlated negatively with trypomastigotes counts, where its expression was low at peak of parasitemia and it increased coinciding with low parasites number. So, this enzyme, through the production of NO, seemed to play an important role in controlling T. evansi infection and brain was the organ which participated strongly in its production. This is the first report which studied iNOS expression during T. evansi infection and these results denoted an important role of NO in controlling T. evansi infection. Our results were in parallel with other reports by Paim et al.30, which revealed the importance of NO in controlling trypanosomes infections and found that 3-NT residues, the indirect determinant of NO, was low in the striatum of brain of T. evansi infected rats on 5 days p.i. and it increased in the cerebellum, cerebral cortex and hippocampus on day 10 and 20 p.i. Also, da Silva et al.28, reported a protective role of NO against T. evansi in the experimentally infected rat as indicated by increasing serum nitrites/nitrates level, end products of NO, when the number of circulating parasites was low with T. evansi on 15 and 30 days p.i.
In this study, IL-17 increased, whether in the sera or locally in the tissues of T. evansi infected mice, therefore IL-17 might enhance the production of NO indirectly through inducing production of various pro-inflammatory cytokines, such as TNF-α, IL-6 and IL-1 which in turn mediate the transcription of i-NOS enzyme19, IL-17 has a direct ability to up-regulate expression of mRNA of iNOS31 and IL-17 synergizes with IFN-γ to enhance iNOS expression32.
| Table 2: | IL-17 expression in different tissues of T. evansi infected mice subgroups |
 | |
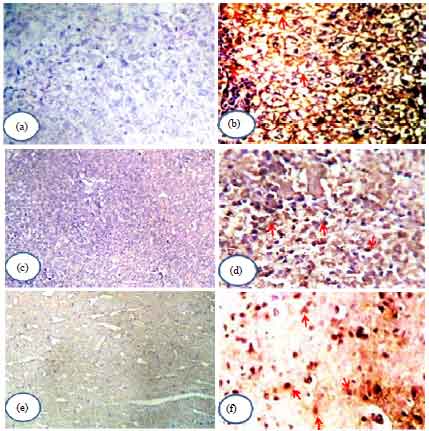 | |
| Fig. 4(a-f): | IL-17 expression in the tissues, (a) Liver of control uninfected mice has no IL-17 expression (x100), (b) Liver of infected mice shows IL-17 expressed in grade III (x200)(red head arrows), (c) Spleen of control uninfected mice has no IL-17 expression (x100), (d) Spleen of infected mice reveals IL-17 expressed in grade III (x200)(red head arrows), (e) Brain of control uninfected mice has no IL-17 expression (x200) and (f) Brain of infected mice shows IL-17 expressed in grade III (x400)(red head arrows) |
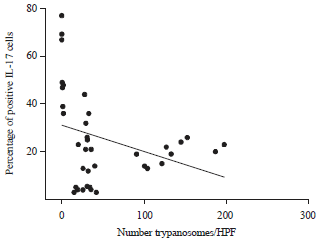 | |
| Fig. 5: | Correlation between percent of positive IL-17 and number of T. evansi trypomastigotes in infected mice (p<0.05 and r = -0.31) |
The generated peroxynitrite anion that has a direct lethal effect on many microbes and it mediates production of hydroxyl radical which can initiate lipid peroxidation and the strong oxidative capacity of NO and its by-products could act in synergism with other lethal molecules like H2O2 to form the toxic antimicrobial network of the host33.
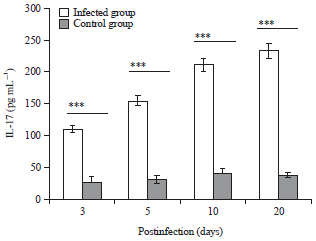 | |
| Fig. 6: | Serum level of IL-17 in T. evansi infected mice subgroups compared with the uninfected controls. *Represents a statistical difference (***p<0.001) |
Herein, by the immunohistochemical method, IL-17 was expressed more by the spleen. The expression of the cytokine negatively sscorrelated with the level of parasitemia where it becoming more with the lowering of the infection, in the subgroup I-20, where 90% of spleens, 70% of livers and only 20 % of brains showed the cytokine with grade III. The IL-17 serum level was increasing significantly as the infection progressed reaching its highest level at end of the experiment and this indicated the protective role of IL-17 against T. evansi infection in experimentally infected mice. It is worthy to mention that this is the first report that measured IL-17 in T. evansi infected host. IL-17 shows a strong role in combating other trypanosoma species infectionm34.
IL-17 is a strong pro-inflammatory cytokine and it has been reported to have an important role in host protection against bacteria, fungi and parasites35. In addition to Th-17 cells, CD8+T cells, neutrophils, monocytes and natural killer cells can produce this cytokine. IL-17 can induce macrophages and other types of cells to produce other several pro-inflammatory cytokines, such as IFN-γ, IL-1, IL-6 and TNF-α mediating iNOS activation added to some chemokines production19. The protective role of IL-17 against T. evansi might be through regulation of the Th1 cells differentiation and pro-inflammatory cytokines production36, induction of iNOS34, moreover IL-17 was suggested to trap T. cruzi in endosomal/lysosomal compartments of the macrophages for longer time37 and this could be in the case of T. evansi. Also, Il-17 has proven to control other parasites. IL-17 was detected in 80% of resistant persons to L. donovani in Sudan38. Mice genetically deficient in IL-17 A receptor have high L. infantum parasitemia, in addition to low IFN-γ and iNOS mRNA39.
CONCLUSION
Trypanosoma evansi infected mice showed parasitemia which reached its peak at 5 days p.i... Both iNOS and IL-17 expression, in addition to IL-17 serum level, increased as the number of the parasites decreased referring strongly to their role in controlling T. evansi in the infected mice.
SIGNIFICANCE STATEMENT
This study can be beneficial to give a novel aspect dealing about the roles of NO Synthase and Interleukin-17 for control of economically important, zoonotic parasite species causing trypanosomiasis infection (Trypanosoma evansi ) in experimentally infected mice. The T. evansi infected mice showed parasitemia which reached its peak at 5 days p.i. The study will help the researcher to explore and know an important information about both iNOS and IL-17 expression, in addition to IL-17 serum level, increased as the number of the parasites decreased referring strongly to their role in controlling T. evansi infection.
REFERENCES
- Narnaware, S.D., S.K. Ghorui, S. Kumar and N.V. Patil, 2016. Vertical transmission of Trypanosoma evansi in dromedary camels and studies on fetal pathology, diagnosis and treatment. Acta Parasitol., 61: 329-336.
CrossRefDirect Link - Lai, D.H., H. Hashimi, Z.R. Lun, F.J. Ayala and J. Lukes, 2008. Adaptations of Trypanosoma brucei to gradual loss of kinetoplast DNA: Trypanosoma equiperdum and Trypanosoma evansi are petite mutants of T. brucei. Proc. Natl. Acad. Sci. USA., 105: 1999-2004.
CrossRefDirect Link - Desquesnes, M., A. Dargantes, D.H. Lai, Z.R. Lun, P. Holzmuller and S. Jittapalapong, 2013. Trypanosoma evansi and surra: A review and perspectives on transmission, epidemiology and control, impact and zoonotic aspects. BioMed Res. Int., Vol. 2013.
CrossRef - Shaapan, R.M., 2016. The common zoonotic protozoal diseases causing abortion. J. Parasitic Dis., 40: 1116-1129.
CrossRefDirect Link - Holland, W.G., F. Claes, L.N. My, N.G. Thanh and P.T. Tam et al., 2001. A comparative evaluation of parasitological tests and a PCR for Trypanosoma evansi diagnosis in experimentally infected water buffaloes. Vet. Parasitol., 97: 23-33.
CrossRefDirect Link - Truc, P., P. Buscher, G. Cuny, M.I. Gonzatti and J. Jannin et al., 2013. Atypical human infections by animal trypanosomes. PLoS Neglected Trop. Dis., Vol. 7.
CrossRef - Haridy, F.M., M.T. El-Metwally, H.H. Khalil and T.A. Morsy, 2011. Trypanosoma evansi in dromedary camel: With a case report of zoonosis in greater Cairo, Egypt. J. Egypt. Soc. Parasitol., 41: 65-76.
PubMedDirect Link - Joshi, P.P., A. Chaudhari, V.R. Shegokar, R.M. Powar and V.S. Dani et al., 2006. Treatment and follow-up of the first case of human trypanosomiasis caused by Trypanosoma evansi in India. Trans. Royal Soc. Trop. Med. Hygiene, 100: 989-991.
CrossRefDirect Link - Van Vinh Chau, N., L.B. Chau, M. Desquesnes, S. Herder and N.P.H. Lan et al., 2016. A clinical and epidemiological investigation of the first reported human infection with the zoonotic parasite Trypanosoma evansi in Southeast Asia. Clin. Infect. Dis., 62: 1002-1008.
CrossRefDirect Link - Abdel-Rady, A., 2008. Epidemiological studies (parasitological, serological and molecular techniques) of Trypanosoma evansi infection in camels (Camelus dromedarius) in Egypt. Vet. World, 1: 325-328.
Direct Link - Saleh, M.A., M.B. Al-Salahy and S.A. Sanousi, 2009. Oxidative stress in blood of camels (Camelus dromedaries) naturally infected with Trypanosoma evansi. Vet. Parasitol., 162: 192-199.
CrossRefPubMedDirect Link - Amer, S., O. Ryu, C. Tada, Y. Fukuda, N. Inoue and Y. Nakai, 2011. Molecular identification and phylogenetic analysis of Trypanosoma evansi from dromedary camels (Camelus dromedarius) in Egypt, a pilot study. Acta Trop., 117: 39-46.
CrossRefDirect Link - Hilali, M., A. Abdel-Gawad, A. Nassar and A. Abdel-Wahab, 2006. Hematological and biochemical changes in water buffalo calves (Bubalus bubalis) infected with Trypanosoma evansi. Vet. Parasitol., 139: 237-243.
CrossRefPubMedDirect Link - Monzon, C.M., C.B. Hoyos and G.A. Jara, 1995. Outbreaks of equine trypanosomiasis caused by Trypanosoma evansi in Formosa Province, Argentina. Rev. Sci. Techn. (Int. Office Epizootics), 14: 747-752.
PubMedDirect Link - Wernery, U., R. Zachariah, J.A. Mumford and T. Luckins, 2001. Preliminary evaluation of diagnostic tests using horses experimentally infected with Trypanosoma evansi. Vet. J., 161: 287-300.
CrossRefDirect Link - Masiga, D.K., A.J. Smyth, P. Hayes, T.J. Bromidge and W.C. Gibson, 1992. Sensitive detection of trypanosomes in tsetse flies by DNA amplification. Int. J. Parasitol., 22: 909-918.
CrossRefDirect Link - Poulos, T.L. and H. Li, 2017. Nitric oxide synthase and structure-based inhibitor design. Nitric Oxide, 63: 68-77.
CrossRefDirect Link - Kandhare, A.D., M.V. Patil and S.L. Bodhankar, 2015. L-arginine attenuates the ethylene glycol induced urolithiasis in ininephrectomized hypertensive rats: Role of KIM-1, NGAL and NOs. Renal Fail., 37: 709-721.
CrossRefDirect Link - Burke, S.J., B.L. Updegraff, R.M. Bellich, M.R. Goff and D. Lu et al., 2013. Regulation of iNOS gene transcription by IL-1β and IFN-γ requires a coactivator exchange mechanism. Mol. Endocrinol., 27: 1724-1742.
CrossRefDirect Link - Kumar, S., R.K. Singh and T.R. Bhardwaj, 2017. Therapeutic role of nitric oxide as emerging molecule. Biomed. Pharm., 85: 182-201.
CrossRefDirect Link - Gu, C., L. Wu and X. Li, 2013. IL-17 family: Cytokines, receptors and signaling. Cytokine, 64: 477-485.
CrossRefDirect Link - Karri, S.K. and A. Sheela, 2017. Potential route of Th17/Treg cell dynamics in targeting type 1 diabetes and rheumatoid arthritis: An autoimmune disorder perspective. Br. J. Biomed. Sci., 74: 8-15.
CrossRefDirect Link - Wilson, K.T., S. Fu, K.S. Ramanujam and S.J. Meltzer, 1998. Increased expression of inducible nitric oxide synthase and cyclooxygenase-2 in Barrett's esophagus and associated adenocarcinomas. Cancer Res., 58: 2929-2934.
Direct Link - Du, W.J., J.H. Zhen, Z.Q. Zeng, Z.M. Zheng, Y. Xu, L.Y. Qin and S.J. Chen, 2013. Expression of interleukin-17 associated with disease progression and liver fibrosis with hepatitis B virus infection: IL-17 in HBV infection. Diagnostic Pathol., Vol. 8.
CrossRef - Shaapan, R.M., F.A. El-Nawawi and M.A.A. Tawfik, 2008. Sensitivity and specificity of various serological tests for the detection of Toxoplasma gondii infection in naturally infected sheep. Vet. Parasitol., 153: 359-362.
CrossRefDirect Link - Bal, M.S., L.D. Singla, H. Kumar, A. Vasudev, K. Gupta and P.D. Juyal, 2012. Pathological studies on experimental Trypanosoma evansi infection in swiss albino mice. J. Parasitic Dis., 36: 260-264.
CrossRefDirect Link - Da Silva, C.B., P. Wolkmer, F.C. Paim, A.S. da Silva and L.C. Siqueira et al., 2013. Iron metabolism and its relationship to anemia and immune system in Trypanosoma evansi infected rats. Exp. Parasitol., 133: 357-364.
CrossRefDirect Link - Tatsch, E., G.V. Bochi, R.S. Pereira, H. Kober and V.A. Agertt et al., 2011. A simple and inexpensive automated technique for measurement of serum nitrite/nitrate. Clin. Biochem., 44: 348-350.
CrossRefDirect Link - Paim, F.C., A.S. da Silva, P. Wolkmer, M.M. Costa and C.B. da Silva et al., 2011. Trypanosoma evansi: Concentration of 3-nitrotyrosine in the brain of infected rats. Exp. Parasitol., 129: 27-30.
CrossRefDirect Link - Krstic, A., A. Krstic, V. Ilic, A. Krstic and V. Ilic et al., 2009. p38 MAPK signaling mediates IL-17-induced nitric oxide synthase expression in bone marrow cells. Growth Factors, 27: 79-90.
CrossRefDirect Link - Gao, Q., Y. Liu, Y. Wu, Q. Zhao and L. Wang et al., 2016. IL-17 intensifies IFN-γ-induced NOS2 upregulation in RAW 264.7 cells by further activating STAT1 and NF-κB. Int. J. Mol. Med., 37: 347-358.
CrossRefDirect Link - Sun, J., C. Steenbergen and E. Murphy, 2006. S-nitrosylation: NO-related redox signaling to protect against oxidative stress. Antioxid. Redox Signaling, 8: 1693-1705.
CrossRefDirect Link - Mou, Z., P. Jia, S. Kuriakose, F. Khadem and J.E. Uzonna, 2010. Interleukin-17-mediated control of parasitemia in experimental Trypanosoma congolense infection in mice. Infect. Immun., 78: 5271-5279.
CrossRefDirect Link - Banerjee, A., P. Bhattacharya, A.B. Joshi, N. Ismail, R. Dey and H.L. Nakhasi, 2016. Role of pro-inflammatory cytokine IL-17 in Leishmania pathogenesis and in protective immunity by Leishmania vaccines. Cell. Immunol., 309: 37-41.
CrossRefDirect Link - Miyazaki, Y., S. Hamano, S. Wang, Y. Shimanoe, Y. Iwakura and H. Yoshida, 2010. IL-17 is necessary for host protection against acute-phase Trypanosoma cruzi infection. J. Immunol., 185: 1150-1157.
CrossRefDirect Link - Erdmann, H., C. Roßnagel, J. Bohme, Y. Iwakura, T. Jacobs, U.E. Schaible and C. Holscher, 2013. IL-17A promotes macrophage effector mechanisms against Trypanosoma cruzi by trapping parasites in the endolysosomal compartment. Immunobiology, 218: 910-923.
CrossRefDirect Link - Pitta, M.G.R., A. Romano, S. Cabantous, S. Henri and A. Hammad et al., 2009. IL-17 and IL-22 are associated with protection against human kala azar caused by Leishmania donovani. J. Clin. Invest., 119: 2379-2387.
CrossRefPubMedDirect Link - Nascimento, M.S.L., V. Carregaro, D.S. Lima-Junior, D.L. Costa and B. Ryffel et al., 2014. Interleukin 17A acts synergistically with interferon γ to promote protection against Leishmania infantum infection. J. Infect. Dis., 211: 1015-1026.
CrossRefDirect Link