
Hassan A. Elfadaly
Department of Zoonotic Diseases, National Research Centre, Giza, Egypt
Mohey A. Hassanain
Department of Zoonotic Diseases, National Research Centre, Giza, Egypt
LiveDNA: 20.10587
Raafat M. Shaapan
Department of Zoonotic Diseases, National Research Centre, Giza, Egypt
Nawal A. Hassanain
Department of Zoonotic Diseases, National Research Centre, Giza, Egypt
Ashraaf M. Barakat
Department of Zoonotic Diseases, National Research Centre, Giza, Egypt
International Journal of Zoological Research
Year: 2017 | Volume: 13 | Issue: 1 | Page No.: 6-11







ABSTRACT
Background: The wastages nourished small ruminants and poultry are still free fed on street wastages and possibly exposed to T. gondii oocysts through feces of outdoor shedder cats and they are regarded as high prevalent sources for human toxoplasmosis via their meat containing viable T. gondii tissue cysts. Materials and Methods: A total No. of 859 samples of both blood and their matching tissue were collected from wastages nourished 455 sheep, 237 goats, 124 chickens and 43 ducks respectively from Giza governorate, Egypt. All animals were assayed serologically using Latex Agglutination Test (LAT) as a screen test and the results were confirmed by ELISA. Tissue samples which were identical to seropositive sera were digested and microscopically examined and exposed to DNA confirmation. The microscopic definite bradyzoites containing sera were bio-assayed through intra-peritoneal passage in mice as viability test to determine both LD50 and LD100 for each species isolate. Histopathological examination was done on symptomatic morbid and dead mice. Results: Corresponding to small ruminants and poultry, results of seropositive percentages were 47.5 and 29.3%, total microscopic 30.1 and 32.7%, DNA detection 74.8 and 71.4% and the total percentages of mice viability test 39.4 and 31.3%. In Addition, the total percentages of LD50 were 30.3 and 31.3%, while the LD100 were 9.1 and 0% in small ruminants and poultry respectively. The histopathological examination of inoculated mice signified cyst forming T. gondii in acute and chronic lesions within vital organs. Conclusion: The wastages nourished small ruminants and poultry is of zoonotic impact and significance and must be directed for incriminate this animal feeding pattern and for avoiding consumption under cooked meat of animals or birds.
PDF Abstract XML References Citation
Received: October 30, 2016;
Accepted: November 30, 2016;
Published: December 15, 2016
How to cite this article
Hassan A. Elfadaly, Mohey A. Hassanain, Raafat M. Shaapan, Nawal A. Hassanain and Ashraaf M. Barakat, 2017. Detection of Toxoplasma gondii from Wastage Nourished Small Ruminant and Poultry: Zoonotic Significance. International Journal of Zoological Research, 13: 6-11.
DOI: 10.3923/ijzr.2017.6.11
URL: https://scialert.net/abstract/?doi=ijzr.2017.6.11
DOI: 10.3923/ijzr.2017.6.11
URL: https://scialert.net/abstract/?doi=ijzr.2017.6.11
INTRODUCTION
Postnatal human’s toxoplasmosis is mainly through consuming undercooked meat containing tissue cysts plus to food or water contaminated with environmental oocysts shed in cat feces1. Also, opportunistic toxoplasmosis during pregnancy is possibly series to estradiol hormonal shift2 or due to temporary gravidity hyperglycemia3 in addition to anti-inflammatory corticosteroids therapy4.
Meat tissue cysts signify the main source for human toxoplasmosis, attributable to its persistence viability in under cooked tissues5. Consequence to the high cost of animal ration in growing countries, some animals and poultry flocks are still free nourished on street wastages of human residuals and share habitats with the T. gondii oocysts shedder cats in their subset. So, the wastages nourished animals or poultry possible represent higher prevalent source of human toxoplasmosis through eating inadequately cooked meat containing tissue cysts6.
Little is known of the biological characters of isolates from the Middle East and Africa7. Moreover, there have been relatively few recent reports on Egyptian small ruminants and poultry concerning toxoplasmosis8-10. Serological assay is not enough for confirming T. gondii tissue cysts harboring animals. However, the present study aimed to evaluate the zoonotic bio-hazard of seropositive wastages nourished animals and poultry through mice viability test and DNA recognition along with histopathological examination of inoculated mice reflecting to how extent the possible human infection via meat harboring virulent tissue cysts.
MATERIALS AND METHODS
Blood and tissue samples: A total number of 859 blood samples plus its corresponding tissue samples were taken from wastage nourished Egyptian sheep (455), goats (237), chickens (124) and duck flocks (43) in different rural areas in Giza governorate, Egypt.
Serological assays: Sera were separated, labeled and kept at -20°C until examined serologically against toxoplasma infection by Latex Agglutination Test (LAT) according to the manufacturer’s instructions (Toxocheck-MT, Eiken Chemical, Tokyo, Japan), results were considered positive when agglutination observed at dilutions of 1:64 and greater. Also, enzyme linked immunosorbent assay (ELISA) was carried out according to the method described by Aubert et al.11 using soluble crude antigen prepared from Toxoplasma RH strain tachyzoites.
Preparation of meat samples and digestion: Tissue samples were prepared as described elsewhere12,13. Twenty grams of the equivalent tissue samples from diaphragm and thigh muscles were collected from small ruminants during individual slaughtering outside abattoirs and from various batches of free range chickens and ducks from diverse villages in rural areas in Giza governorate, Egypt. The tissues were cut into small cubes about 5×5×5 cm and classified into two groups, the 1st was frozen at -80oC for further DNA extraction while the 2nd group was exposed to pepsin digestion and then bio-assayed for both microscopic examination and mice viability test.
Viability test with LD50 and LD100 determination in mice: The test procedures were done according to Elfadaly et al.2, a total number of 115 seronegative Swiss Webster Albino mice obtained from Laboratory Animals House, National Research Centre, Egypt were classified corresponding to animals and poultry isolates as 74 for sheep, 25 for goats, 11 for chickens and 5 for ducks. The inoculated mice were followed up daily for any apparent clinical signs of febrile response or acute toxoplasmosis with the exclusion of dead mice before 48 h of inoculation. The predicted signs may be varied between ascites, roughcast hairs, tottering gait, hunched appearance, with evidence of early emaciation and dehydration or death usually within 72-84 h (with highly virulent types) or nervous manifestations and partial off food (with moderately virulent types). Depending on the virulence of the isolate, the procedure will be continued. If ascites occurred, peritoneal exudates were collected from ascetic mice within 72-84 h DPI for microscopic examination for tachyzoites. If mice didn’t show ascites they were sacrificed within 15th day post inoculation by cervical dislocation, samples were collected from heart, lung, liver, spleen and kidney for histopathological examination according to Ajzenberg et al.14. The LD50 and LD100 characters are differentiate the dissimilar virulent among varied isolates which could kill 50 or 100% of inoculated mice, respectively were recorded for each species isolates.
DNA detection: Preparation of specimens and isolation of DNA for PCR was carried out according to Burg et al.15.
Extraction of genomic DNA of T. gondii from the collected tissue: Samples were extracted by a kit (ViVantis Co., Malaysia).
PCR amplification of B1 gene: The B1 gene was amplified16, using primers 1 (5’-TCG GAG AGA GAA GTT CGTCGC AT-3’) and 2 (5’-AGC CTC TCT CTT CAA GCA GCG TA-3’). The following reaction mixture was added in a 0.2 mL PCR tubes: DNA template (100 ng μL‾1) 10 μL, Taq polymerase (5 μ μL‾1) 1 μL, 10x enzyme buffer 2 μL, dNTPs 0.8 μL, each primer 1 μL and Bidest water to 20 μL. The mixture was briefly spine and placed in the thermal cycler (T-gradient, Biometra, Germany), which was programmed as follow: Initial denaturing 95°C/2 min and 40 cycles consisting of denaturing 95°C/1 min, annealing 55°C/30 sec, extension 72°C/45 sec and final extension 72°C/10 min. The PCR products were electrophoresed at 80 v/15 min and finally examined using UV trans-illuminator. About 100 bp DNA ladder (Finnzymes) was used as a marker.
Histopathological methods: Tissue specimens about 0.5 cm3 were individually obtained from lung, liver, spleen, brain, kidney and cardiac muscles of inoculated mice, fixed in 10% neutral buffered formal saline solution, sectioned and further processed according to conventional routine histological technique as described by Biancifiori et al.17, including cutting at 5 μm and staining with hematoxylin and eosin (H and E), Periodic Acid-Schiff (PAS) and toluidine blue stains. The stained tissue sections were examined by optical microscope at power magnification of x400 and the protozoal associated lesions, pathological alterations and the T. gondii bradyzoites were scored.
Ethical approval: The study was approved ethically by the Medical Research Ethics Committee, National Research Centre, Egypt under registration No. 1-2 /0-2-1.2012.
RESULTS
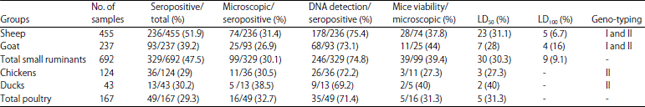
The serologic, microscopic, DNA, mice viability and LD50 and LD100 results were displayed in Table 1.
Seropositive percentages of T. gondii infection were recorded as 51.9, 39.2, 29 and 30.2% corresponding to sheep, goats, chickens and ducks with total seropositive percentages within small ruminants and poultry 47.5 and 29.3%, respectively.
Histopathological lesions of various vital organs of acute infected mice within 12 DPI through T. gondii isolates from small ruminants or poultry were recorded. The mice organs showed tachyzoites infiltration: Kidney, peritoneal exudates tachyzoites and leukocytes at acute phase 72 HPI, spleen, liver, cardiac muscles and lung (Fig. 1).
| Table 1: | Seropositive, microscopic examination, DNA detection, mice viability and LD50 and LD100 of small ruminants and poultry examined samples |
 | |
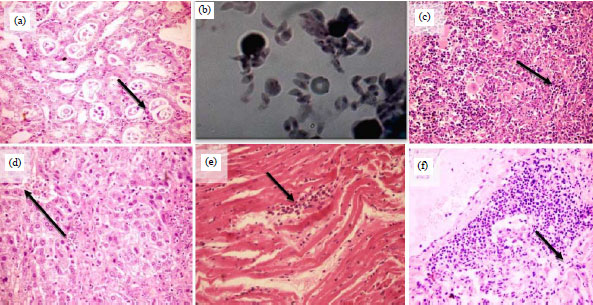 | |
| Fig. 1(a-f): | Histopathological lesions of various vital organs of acute infected mice within12 DPI with T. gondii isolates from small ruminants or poultry. The mice organs show tachyzoites infiltration, (a) Kidney, (b) Mice peritoneal exudates tachyzoites and leukocytes at acute phase 72 HPI, (c) Spleen, (d) Liver, (e) Cardiac muscles and (f) Lung (X400) |
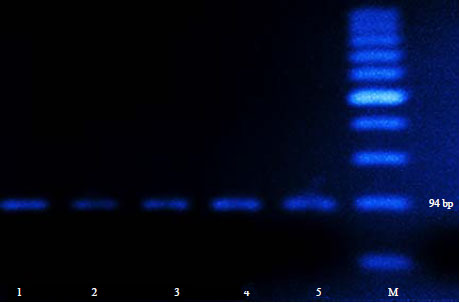 | |
| Fig. 2: | An agarose gel electrophoresis showing PCR amplification product of T. gondii using B1 gene. Lane M: Marker, Lane 1: Positive control, Lanes 2, 3: Positive T. gondii DNA Egyptian isolates at 94 bp corresponding to sheep and goat, Lanes 4, 5: Positive samples of isolated chicken and ducks isolates at 94 bp |
The microscopic examination and DNA detection of T. gondii were only performed on the equivalent digested tissue samples of positive sera, the percentage values were 31.4 and 75.4, 26.9 and 73.1, 30.5 and 72.2 and 38.5 and 69.2 corresponding to sheep, goats, chickens and ducks, respectively and the total microscopic and DNA detection percentages within small ruminants and poultry were 30.1, 74.8 and 32.7, 71.4%, respectively (Fig. 2).
The mice viability test of T. gondii was only concerned with the microscopic positive tissue samples which were showing bradyzoites similar protozoa, the recorded values and percentages were 28/74 (37.8) sheep, 11/25 (44) goat, 3/11 (27.3) chickens and 2/5 (40) ducks (Table 1, Fig. 1, 2), the total percentages of mice viability test were 39.4 and 31.3 corresponding to small ruminants and poultry, respectively.
The T. gondii LD50 and LD100 were recorded varied percentage values 31.1 and 6.7, 28 and 16, 27.3 and 0 and 40 and 0 with the corresponding to sheep, goats, chickens and ducks, respectively with total percentages within small ruminants and poultry 30.3, 9.1 and 31.3, 0, respectively.
DISCUSSION
Up to 60% of Egyptians are T. gondii seropositive3. This higher ratio may be linked to consuming undercooked mutton or poultry meat rather than oocysts contaminated food or water. The current study supports this concept through definite wastages nourished small ruminants and poultry as a great source of toxoplasmosis through overall higher seropositive ratios 47.5 and 29.3, respectively. While in the previous Egyptian study; sheep was recorded lower seropositive value 34%18, the higher difference between the two studies possible contributed to the wastage nourished feeding behavior, which usually maximize exposure with sporulated T. gondii oocysts13. Also, the results agree with Dubey et al.8 who were run with our results and overcome high prevalence in Egyptian free range chickens 40.4 and ducks 15% from rural area surrounding Giza. This probably due to free range poultry mainly infected via environmental oocysts. Moreover, they like omnivore’s species, possible feed on tissue cyst through engulfing raw meat remaining19.
The obtained results denote that not all seropositive animals or birds were harboring virulent T. gondii bradyzoites in their tissues. Accurately; serological assay does not reveal tissue cyst harboring hosts. So, the current study evaluated the zoonotic bio-hazard of seropositive small ruminants and poultry through mouse viability test and histopathology of inoculated mice along with DNA recognition, act as specific T. gondii bio-indicators for the possible human infection. However, the main significant factors shared in non-synchronized results between seropositive hosts and their tissues containing cysts probably due to the transferred maternal antibodies or sequence to incompatibility between the collected examined samples with the carcasses/tissue cyst distribution2, besides the possible cross-reactivity with other parasites as Trichinella spiralis20. However, mouse viability test is accurate and sensitive than DNA or microscopic tests, it firmly excludes the possible presence of other T. gondii cyst forming protozoa or T. gondii non virulent strains which could be exciting PCR cycle through DNA similar sequence but they could not induce morbid mice.
According to mice viability and histopathology tests, goats were recorded the highest species harboring T. gondii tissue cysts (44%), followed by sheep (37.8%), while poultry was recorded the lowest value (31.3%). The results agree with most studies which signified sheep and goats as the most T. gondii susceptible species21,22. The equivalent digested tissue samples of small ruminants and poultry were confirmed DNA positive and reflect higher PCR sensitivity than the inferior microscopic ones.
The LD50 and LD100 are the lethal dosages that differentiate the dissimilar virulent among varied isolates which could kill 50 or 100% of inoculated mice respectively. It has been suggested that type I and II are more prevalent in clinical toxoplasmosis16. The results in current study signified a total 39.4% of small ruminants and 31.3% of poultry isolates were successfully passed in to mice with dissimilar morbidity and mortality ratios. Also, small ruminants were set higher in total LD50 and LD100 30.3 and 9.1% than poultry 31.3 and 0%. Our results are corporate other studies which validated the vast majority of virulent Egyptian sheep and poultry isolates were corresponding to type II strains which the predominant human types6. Also, from São Paulo state, Brazil, T. gondii was isolated from tissue homogenates of 16 sheep, where six of the 16 isolates killed 100% of infected mice23.
Mixed infection in the same meat sample may be involved but higher virulent types hides the biological properties of less virulent ones24. Therefore, the parasite isolation practice could results in a single strain even several strains were found in the inoculum.
CONCLUSION
The bio-viability and histopathology along with DNA confirm of seropositive hosts are on target T. gondii tissue cysts bio-indicators, signify zoonotic impact of wastages nourished small ruminants and poultry, must be directed for incriminate this animal feeding pattern and for avoiding consumption under cooked meat of animals or birds.
ACKNOWLEDGMENT
Researchers are thankful to Department Zoonotic Diseases Department, National Research Centre, Egypt for providing technical and financial assistance to carry out this research work.
REFERENCES
- Tenter, A.M., A.R. Heckeroth and L.M. Weiss, 2000. Toxoplasma gondii: From animals to humans. Int. J. Parasitol., 30: 1217-1258.
CrossRefPubMedDirect Link - Elfadaly, H.A., M.A. Hassanain, R.M. Shaapan, A.M. Barakat and N.I. Toaleb, 2012. Serological and hormonal assays of murine materno-fetal Toxoplasma gondii infection with emphasis on virulent strains. World J. Med. Sci., 7: 248-254.
Direct Link - Hassanain, M.A., H.A. El-Fadaly and N.A. Hassanain, 2014. Toxoplasma gondii parasite load elevation in diabetic rats as latent opportunistic character. Ann. Trop. Med. Pubblic Health, 7: 110-115.
Direct Link - Elfadaly, H.A., M.A. Hassanain, R.M. Shaapan, N.A. Hassanain and A.M. Barakat, 2015. Corticosteroids opportunist higher Toxoplasma gondii brain cysts in latent infected mice. Int. J. Zool. Res., 11: 169-176.
CrossRefDirect Link - Fromont, E.G., B. Riche and M. Rabilloud, 2009. Toxoplasma seroprevalence in a rural population in France: Detection of a household effect. BMC Infect Dis., Vol. 9.
CrossRefDirect Link - Hassanain, M.A., H.A. Elfadaly, R.M. Shaapan, N.A. Hassanain and A.M. Barakat, 2011. Biological assay of Toxoplasma gondii Egyptian mutton isolates. Int. J. Zool. Res., 7: 330-337.
CrossRefDirect Link - Dubey, J.P., 2009. Toxoplasmosis in sheep-The last 20 years. Vet. Parasitol., 163: 1-14.
CrossRefPubMedDirect Link - Dubey, J.P., D.H. Graham, E. Dahl, M. Hilali and A. El-Ghaysh et al., 2003. Isolation and molecular characterization of Toxoplasma gondii from chickens and ducks from Egypt. Vet. Parasitol., 114: 89-95.
CrossRefPubMedDirect Link - Barakat, A.M.A., M.M. Abd Elaziz and H.A. Fadaly, 2009. Comparative diagnosis of toxoplasmosis in Egyptian small ruminants by indirect hemagglutination assay and ELISA. Global Veterinaria, 3: 9-14.
Direct Link - Ghoneim, N.H., S.I. Shalaby, N.A. Hassanain, G.S.G. Zeedan, Y.A. Soliman and A.M. Abdalhamed, 2010. Comparative study between serological and molecular methods for diagnosis of toxoplasmosis in women and small ruminants in Egypt. Foodborne Pathog. Dis., 7: 17-22.
CrossRefDirect Link - Aubert, D., G.T. Maine, I. Villena, J.C. Hunt and L. Howard et al., 2000. Recombinant antigens to detect Toxoplasma gondii-specific immunoglobulin G and immunoglobulin M in human sera by enzyme immunoassay. J. Clin. Microbiol., 38: 1144-1150.
Direct Link - Sharma, S.P. and J.P. Dubey, 1981. Quantitative survival of Toxoplasma gondii tachyzoites and bradyzoites in pepsin and in trypsin solutions. Am. J. Vet. Res., 42: 128-130.
PubMedDirect Link - Ajzenberg, D., A.L. Banuls, M. Tibayrenc and M.L. Darde, 2002. Microsatellite analysis of Toxoplasma gondii shows considerable polymorphism structured into two main clonal groups. Int. J. Parasitol., 32: 27-38.
CrossRefPubMedDirect Link - Burg, J.L., C.M. Grover, P. Pouletty and J.C. Boothroyd, 1989. Direct and sensitive detection of a pathogenic protozoan, Toxoplasma gondii, by polymerase chain reaction. J. Clin. Microbiol., 27: 1787-1792.
PubMedDirect Link - Howe, D.K., S. Honore, F. Derouin and L.D. Sibley, 1997. Determination of genotypes of Toxoplasma gondii strains isolated from patients with toxoplasmosis. J. Clin. Microbiol., 35: 1411-1414.
Direct Link - Biancifiori, F., C. Rondini, V. Grelloni and T. Frescura, 1986. Avian toxoplasmosis: Experimental infection of chicken and pigeon. Comp. Immunol. Microbiol. Infect. Dis., 9: 337-346.
CrossRefDirect Link - Shaapan, R.M., F.A. El-Nawawi and M.A.A. Tawfik, 2008. Sensitivity and specificity of various serological tests for the detection of Toxoplasma gondii infection in naturally infected sheep. Vet. Parasitol., 153: 359-362.
CrossRefDirect Link - Cook, A.J., R.E. Gilbert, W. Buffolano, J. Zufferey and E. Petersen et al., 2000. Sources of toxoplasma infection in pregnant women: European multicentre case-control study. Br. Med. J., 321: 142-147.
CrossRefDirect Link - Hassanain, M.A., N.A Hassanain, R.M. Shaapan, H.A. El Fadaly and F.M. El Mogazy, 2009. Problem of toxoplasmosis and detection of possible cross-reactivity with Trichinella spiralis. Proceedings of the 14th Congress of the International Society for Animal Hygiene, July 19-23, 2009, Vechta, Germany, pp: 97-100.
- Aspinall, T.V., D. Marlee, J.E. Hyde and P.F.G. Sims, 2002. Prevalence of Toxoplasma gondii in commercial meat products as monitored by polymerase chain reaction-food for thought? Int. J. Parasitol., 32: 1193-1199.
CrossRefDirect Link - Dubey, J.P., N. Sundar, D. Hill, G.V. Velmurugan and L.A. Bandini et al., 2008. High prevalence and abundant atypical genotypes of Toxoplasma gondii isolated from lambs destined for human consumption in the USA. Int. J. Parasitol., 38: 999-1006.
CrossRefPubMedDirect Link - Boyer, K.M., E. Holfels, N. Roizen, C. Swisher and D. Mack et al., 2005. Risk factors for Toxoplasma gondii infection in mothers of infants with congenital toxoplasmosis: Implications for prenatal management and screening. Am. J. Obstet. Gynecol., 192: 564-571.
CrossRefPubMedDirect Link - Aspinall, T.V., E.C. Guy, K.E. Roberts, D.H.M. Joynson, J.E. Hyde and P.F.G. Sims, 2003. Molecular evidence for multiple Toxoplasma gondii infections in individual patients in England and Wales: Public health implications. Int. J. Parasitol., 33: 97-103.
CrossRefPubMedDirect Link