
Sameer Bajru
Bombay Natural History Society, Hornbill House, Opp. Lion Gate, Shaheed Bhagat Singh Road, Mumbai, 400 001, Maharashtra, India
Omkar Yadav
Department of Zoology, Shivaji University, Kolhapur, 416 004, Maharashtra, India
Shivanand Yankanchi
Department of Zoology, Shivaji University, Kolhapur, 416 004, Maharashtra, India
International Journal of Zoological Research
Year: 2016 | Volume: 12 | Issue: 1-2 | Page No.: 18-24







ABSTRACT
Although, the Forest Calotes Calotes rouxii is the most common and endemic species of lizard inhabiting the Western Ghats, information on its ecology is remain scarce. In order to better understanding of life history of C. rouxii, we investigated its pre breeding activity patterns and microhabitat utilization in a small sacred grove (~0.65 ha) of the northern Western Ghats. Calotes rouxii showed a unimodal activity pattern, peak activity period was observed at late morning (11.00-12.30 h) and afternoon (13.00-15.45 h). The activity patterns of lizards involved basking and resting at early morning period. Once they attained optimum body temperature at peak period activities such as moving, social displays and foraging were performed. In late afternoon all activities were decreased and they retreat to their refuge sites. Sex specific microhabitat segregation was detected; males occupied higher perches than females. The males encounter during study period were positively correlated with temperature while negatively correlated with humidity, however females encountered were not correlated with humidity and temperature.
PDF Abstract XML References Citation
Received: July 27, 2015;
Accepted: October 19, 2015;
Published: February 15, 2016
How to cite this article
Sameer Bajru, Omkar Yadav and Shivanand Yankanchi, 2016. Daily Activity Patterns and Perch Height Utilization in Pre-Breeding Season by Forest Calotes, Calotes rouxii (Sauria: Agamidae) in the Sacred Grove of the Northern Western Ghats. International Journal of Zoological Research, 12: 18-24.
DOI: 10.3923/ijzr.2016.18.24
URL: https://scialert.net/abstract/?doi=ijzr.2016.18.24
DOI: 10.3923/ijzr.2016.18.24
URL: https://scialert.net/abstract/?doi=ijzr.2016.18.24
INTRODUCTION
Microhabitat use and activity pattern are important component of animal ecology. For lizards in general, activity patterns have been shown to be affected by a wide range of variables such as sex, reproductive behavior and age class; daily and seasonal variations in temperature and other weather conditions and the presence of intra-specific competition (Huey et al., 1977; Rose, 1981; Creusere and Whitford, 1982). Lizard community may achieve the resource portioning by spatial and temporal separation in their activity patterns depending upon the habitat, prey availability and thermal ecology (Zamora-Camacho et al., 2013; Schoener, 1997; Heatwole and Taylor, 1987; Davis and Verbeek, 1972; Zucker, 1986; Martin and Salvador, 1997; Howard and Hailey, 1999; Leal and Fleishman, 2002). Similarly intra specific separation in spatial dimension was also reported in many species of arboreal lizards, male occupy the higher perch than their female counterpart and juveniles (Davis and Verbeek, 1972; Zucker, 1986).
addition to difference in size various size independent morphological attributes such as relative tail length, dewlap size, limb length, wing size (Draco spp.), horn presence or size and head size as well as coloration is common. Establish relationship between the size of dimorphism and habitat use. Thus study of both size and shape dimorphism may be particular informative (Losos et al., 2003).
The Forest Calotes, Calotes rouxii is widely distributed in the forests of the Peninsula especially in the Western Ghats (Daniel, 1992). Mainly it is inhabit in lowlands and mid elevation moist mixed deciduous, semi evergreen and evergreen forests of the Western Ghats. They are diurnal mostly active at base of trees in forests. It is small bodied lizard smaller than Calotes versicolor endemic to India and Bangladesh. It plays a vital role in controlling some insect pests such as cricket, grasshoppers, moths and other insects. They dorsally olive-brown with dark bands along the sides of the head and ventral surface is light brownish (Murthy, 1984). Head is smaller, slender and spines on each side of the tympanum; dark fold in front of shoulder; throat reddish orange in both sexes (Das, 2002). Sexual dimorphism is observed (Fig. 1a and b), males with swollen chicks, small gular sac and swollen tail base covered with thick and enlarged scales. In breeding season males are characterized by break red colour on head, gap and gular sac hence they also called as Forest Bloodsucker (Murthy, 1984). Limbs are moderate, slender and longer adopted for arboreal habit (Smith, 1935).
Studies about habitat use and selection are important to understanding the biological requirements of animals as well as the strategies they use to meet these requirements (Guido and Gainelle, 2001; Manly et al., 2002). Gathering detailed information on patterns of habitat use through studies is also essential for management and conservation purposes (Scott et al., 2002; Guisan and Thuiller, 2005). In this study we investigate microhabitat utilization and activity patterns of C. rouxii during pre breeding season. In addition, we examined whether the microhabitat utilization and activity patterns were correlated with morphology of lizards.
 | |
| Fig. 1(a-b): | Calotes rouxii (a) Male and (b) Female |
MATERIALS AND METHODS
The present study was carried out at the Ambeshwar sacred grove (16°57'25''N, 73°47'44''E). This study site is located 500 m South of Amba village which is 65 km away from Kolhapur District of Maharashtra. The study site comprises of ~0.65 ha of area with average altitude 618.74 m above the sea level. The climate is characterized by average temperature ranges from 15-35°C and average annual rainfall about 3500 mm. This area is a small isolated patch of semi-evergreen forest surrounded by paddy fields and human settlement (Fig. 2). The habitat (Fig. 2) is dominated by Anjani (Memecylon umbellatum), Torch tree (Ixora pavatta) and orange berry (Glycosmis pentaphylla). Forest ground is covered with thick layer of leaf litter and above ground canopy was divided into two distinct strata lower canopy of herbs, shrubs and small trees, Memecylon umbellatum (Melastomacece), Croton zeylanicus (Euphorbiaceae) and Paramignya monophylla (Rutaceae) etc. and higher canopy of large old trees, Caryota urens (Arecaceae), Mangifera indica (Anacardiaceae) and Terminalia chebula (Combretaceae) etc. and woody climbers Gnetum ulva (Gnetaceae), Dalbergia horida (Fabaceae).
The study was conducted during December, 2014 to April 2015; n = 7 days. A single observer was repeatedly surveyed the predetermined path on foot between 0800-1230 h and 1300-1900 h. During each hourly pass, the observer would sample the seven stations located ~10 m apart. At each station, the observer examined the ground and vegetation for lizards. For this 180° scan of vegetation was made, it required 1 min for scanning 5 m radius semicircle. If, individual was sighted observations were made by Olympus 7×35 binocular for 10 min. The path was required ~50 min for completing one sampling cycle and 5 minute pause between sampling cycles, resulting into 9 sample cycles per day (Nicholson et al., 2005).
All animal sampling method was use for collecting the data (Lehner, 1996). Sexual dichromatism in C. rouxii was prominent only during breeding season i.e., May to September (Das, 2002) while we recorded observation in non breeding season. Therefore, sexes were identified by morphological traits such as male having larger head, swollen chick and thick tail base Smith, 1935; Murthy, 1984). Sex of the lizard was determined by direct examination through binocular or in suspected case lizard was caught after taking observation and sex was identified. The various behavioral traits were recorded for each lizard like basking, resting (when sunlight was not falling direct on any part of body of the individual), moving, social displays (head bob, gular pouch extension and pushup) and foraging. The point where lizard sighted at 1 min of 10 min. observation period was considered for perch height. The perch height was measured with measuring tape and visually if individual was beyond reach in various categories from lowest category <2 m to highest category >10 m. Ambient air temperature and relative humidity were recorded with thermometer and hygrometer at every 1 h.
The variations in the number of individuals and activity patterns of lizard during morning, afternoon and late afternoon was analyzed by chi square test.
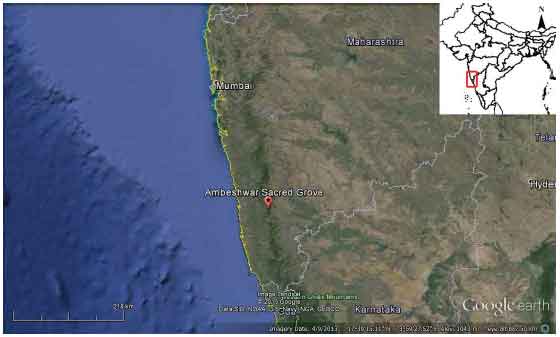 | |
| Fig. 2: | Location map of Ambeshwar sacred grove |
All morphometric variables were log transformed and regressed against SVL before analysis. Perch height and morphometric differences between sexes were analyzed by unpaired Students‘t’ test. Relation between different variables was analyzed by Spearman’s rank correlation. All Statistical analysis was performed by using SPSS 8.0 and MS-Excel. Significance was accepted at p<0.05 level.
RESULTS
The total 63 h (180 min/lizard) of observations were made. There was a significant difference in perch height occupied by males and females (unpaired ‘t’ test; t = 2.10, p<0.05). Female showed non-random distribution ~66% occupied perch height between 2.0-4.0 m, whereas, ~44% at <2.0 m. Males were randomly distributed ~60% at <2.0 m, ~20% at 2.0-4.0 m, ~14% at 4.0-10.0 m and ~7% at >10.0 m (Fig. 3). Minimum perch height occupied by female was 0.40 m and maximum 3.70 m while minimum perch height occupied by male was 1.10 m and maximum 12.0 m.
Frequency of basking, rest, movement, social display and foraging were significantly greater at 11.00-15.45 h (Fig. 4). Even in different time of the day, they perform different activities at different frequencies.
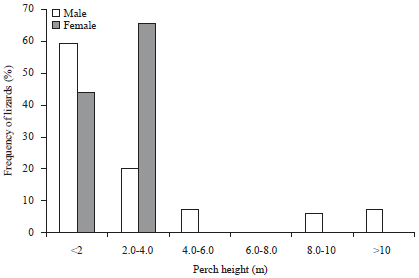 | |
| Fig. 3: | Perch height preference of males and females in Calotes rouxii |
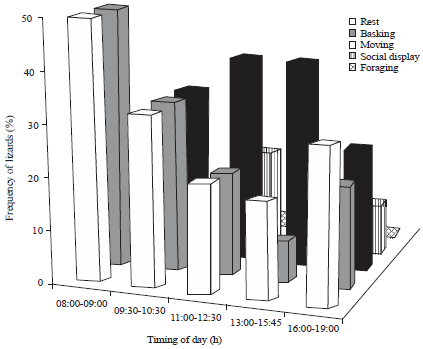 | |
| Fig. 4: | Frequency of different activity patterns in Calotes rouxii |
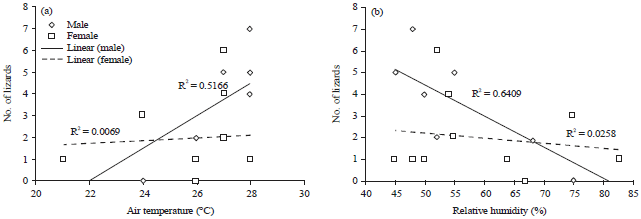 | |
| Fig. 5(a-b): | Number of males and females of Calotes rouxii were recorded in relation to (a) Air temperature and (b) Relative humidity |
For example early morning hours (08.00-09.30 h) only activity performed were basking and resting (~50% lizards), no any movement or social displays. During mid part of morning phase (09.30-10.30 h) basking, rest and movement were equally performed (~33%). In late morning (11.00-12.30 h) basking (~16%) and resting (21%) were declined while movement (~40%) social display and feeding were increased (χ2 =38.18, df =4, p<0.05). Similar trend was observed at afternoon (1300-1545 h) basking and rest were declined while movement (~40%), social display (~13%) and feeding were increased (χ2 = 43.02, df = 4, p<0.05). Late afternoon (16.00-19.00 h) most of the individuals (~35%) were undergo rest and declined other activities (χ2 = 8.96, df = 3, p<0.05).
Total 21±5.2 lizards were recorded, out of which 10±3.0 males and 11±2.2 females. Males and females were responded differently to the air temperature and humidity (Fig. 5a and b). Males were positively correlated with temperature (r = 0.86, p<0.001, n = 63 observations) while negatively correlated with humidity r = -0.80, p<0.001, n = 63 observations).Therefore, the highest encounter rate of lizard was late morning and afternoon hours coincide with high temperature and low humidity period. But females does not show any significant relation with temperature r = -0.013, p>0.05, n= 63 observations) and humidity (r = -0.045, p>0.05, n = 63 observations). Temperature and humidity recorded were highly negatively correlated (r = -0.972, p>0.001, n = 63 observations).
DISCUSSION
The Ambeshwar sacred groove apart from C. rouxii harbors a faunal community which may directly or indirectly influences it. This community comprises of three major components prey, predators and competitors. The potential prey items reported were insects belongs to order Orthoptera (grasshoppers), Hymenoptera (ants), Coleopteran (beetles) and Diptera (flies and mosquitoes). During this study three times lizards were observed resting beside the ant trail and picked up ants marching through the trail, on another occasion one individual was feeding on tortoise beetle while four records of feeding on mosquitoes (personal observations). The possible food competitors recorded were skinks (Mabuya spp.) and insectivorous birds, Orange Headed Thrush (Zoothera citrine), Spot Throated Babbler (Pellorneum albiventre), Common iora (Aegithina tiphia). The potential lizard predators observed were split in two groups, reptiles and aves; reptile predators, Green Vine Snake (Ahaetulla nasuta), Buff Striped Keelback (Amphiesma stolatum), Bamboo Pit Viper (Trimeresurus gramineus) and Malabar Pit Viper (Trimeresurus malbaricus) while avian predators include Shikra (Accipiter badius) and Great Hornbill (Buceros bicornis).
The C. rouxii had significant variations in their daily activity patterns. Morning hours they were basking and rest on lower perches or ground. Once attain the optimum body temperature required for activities they show movement, social displays and feeding behavior. Peak activity period was at late morning (11.00-12.30 h) and afternoon (13.00-15.45 h). Similar trend was observed in Ameiva, very few individuals were active during cooler parts of the day and most observations occurred during the warmer part (Blair, 2009). Activity peaked between 14.30-15.00 h (Nicholson et al., 2005). While the temperature drops down below the optimum level, activities slows down and lizards undergo rest. This unimodal activity pattern was strengthen by Schell et al. (1993) they found the activity of A. chrysolaema restricted during the late morning to early afternoon, coincident with daily maximum temperatures. Our observations support unimodal activity pattern of C. rouxii which was entirely different from bimodal activity pattern of Psammophilus dorsalis and Sitana ponticeriana (Radder et al., 2005). The dense vegetation structure of the study site may be allow the optimum temperature required for lizard activities only attained at mid period (late morning and afternoon) of the day, Hence maximum number of active lizards were recorded during this period. By maintaining an optimal body temperature, a lizard maximizes its ability to perform important functions such as predator avoidance, foraging and reproduction (Huey, 1991). Thermal stratification by vegetation possibly incited unimodal activity pattern in C. rouxii. During unfavorable period of the day lizards were observed in tree holes, natural burrows at base of large trees and resting on perches (personal observation).
Calotes rouxii revealed noticeable sex specific spatial microhabitat segregation. Males occupied the higher perches and randomly distributed. However, females were occupied lower perches and nonrandomly distributed. Similar trend was reported by Radder et al. (2005) in case of Psammophilus dorsalis. Previously both intra and interspecific niche separation within given habitat had been reported for other lizard species. A number of biotic (presence of heterospecific, conspecific, intersexual dietary divergences) and abiotic factors such as temperature (Davis and Verbeek, 1972), light (Martin and Salvador, 1997; Howard and Hailey, 1999), moisture, (Melville and Schulte, 2001; Leal and Fleishman, 2002) are known to influence microhabitat preference in lizards. Li and Lu (1982) suggested that the differential habitat use could be explained in the context of sexual selection in males and natural selection in females. The probable reasons for maintaining higher position on perch were attracting the female and guard the territory. Apart from this basking may be the cause for attaining higher perch position, because in this thick patch of forest only higher canopy receive direct sunlight. Diurnal lizards were primarily communicate using movement based on visual signal often delivered in predictable sequence, together with characteristics posture and posture and in body coloration (Carpenter and Ferguson, 1977) while nocturnal lizards like geckos mainly relay on either vocal or auditory (Frankenberg, 1982) or chemical cues (Greenberg, 1943) for courtship acts. This site have lower stratum of dense vegetation due to network of herbs, shrubs and saplings of trees which may cause obstacle in visual communication for diurnal lizard such as C. rouxii. This may be one of the forces for pushing the male lizard for occupying higher perches. Randomness in perch height occupied by males was possibly due to males are more active than females. They continuously performed activities such as patrolling to guard their territory, displaying for attract the female and repel the rival males. In contrast, females, with cryptic body coloration, may gain by perching near the ground especially during pregnancy when locomotion is retarded by eggs and if the low vegetation offered more food and protection (Kuo et al., 2007).
Thermoregulation influences physiological, behavioral and ecological features in ectotherms (Avery, 1982; Bauwens et al., 1996; Seebacher and Franklin, 2005). Ectotherms such as lizards act to reduce exposure to unfavorable abiotic condition by simply moving out of direct sunlight or by increasing water intake. Thus, understanding the relationship between abiotic condition and activity patterns can important in understanding ecology of animals (Nicholson et al., 2005). Thermoregulation problem was comparatively less acute in C. rouxii than lizards inhabiting on open habitats. Activity patterns were influenced by the two major abiotic factors, temperature and humidity. Only males of C. rouxii showed positive correlation with temperature and negatively correlation with humidity while in female absence of any relationship with these variables. The probable cause behind this relation was different reproductive role played by sexes, males are involved in courtship and territory displays for which they possible need cost effective energy source i.e. temperature. But for females temperature may not be the cost effective source of energy. Diverse response of male and female lizards to temperature and humidity might be misleading, because females are cryptic and less active compared to males therefore chances of their underestimation are more.
ACKNOWLEDGMENTS
We are thankful to the Maharashtra Forest Department for granting permission to conduct the study. Thanks to N.P. Grampurohit, A.A. Kanase for sharing knowledge. We are also grateful to Amolkumar Lokhande, Avinash Adasul, Sachin Patil, Appasaheb Bhusnar for assistance in the field.
REFERENCES
- Bauwens, D., P.E. Hertz and A.M. Castilla, 1996. Thermoregulation in a lacertid lizard: The relative contributions of distinct behavioral mechanisms. Ecology, 77: 1818-1830.
CrossRefDirect Link - Blair, C., 2009. Daily activity patterns and microhabitat use of a Heliothermic lizard, Ameiva exsul (Squamata: Teiidae) in Puerto Rico. South Am. J. Herpetol., 4: 179-185.
CrossRefDirect Link - Kuo, C.Y., Y.S. Lin and Y.K. Lin, 2007. Resource use and morphology of two sympatric Japalura lizards (Iguania: Agamidae). J. Herpetol., 41: 713-723.
CrossRefDirect Link - Davis, J. and N.A.M. Verbeek, 1972. Habitat preferences and the distribution of Uta stansburiana and Sceloporus occidentalis in coastal California. Copeia, 1972: 643-649.
CrossRefDirect Link - Frankenberg, E., 1982. Vocal behavior of the mediterranean house gecko, Hemidactylus turcicus. Copeia, 1982: 770-775.
CrossRefDirect Link - Greenberg, B., 1943. Social behavior of the western banded gecko, Coleonyx variegatus Baird. Physiol. Zool., 16: 110-122.
Direct Link - Guido, M. and D. Gianelle, 2001. Distribution patterns of four Orthoptera species in relation to microhabitat heterogeneity in an ecotonal area. Acta Oecol., 22: 175-185.
CrossRefDirect Link - Guisan, A. and W. Thuiller, 2005. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett., 8: 993-1009.
CrossRefDirect Link - Howard, K.E. and A. Hailey, 1999. Microhabitat separation among diurnal saxicolous lizards in Zimbabwe. J. Trop. Ecol., 15: 367-378.
Direct Link - Huey, R.B., E.R. Pianka and J.A. Hoffman, 1977. Seasonal variation in thermoregulatory behavior and body temperature of diurnal Kalahari lizards. Ecology, 58: 1066-1075.
CrossRefDirect Link - Huey, R.B., 1991. Physiological consequences of habitat selection. Am. Naturalist, 137: S91-S115.
Direct Link - Leal, M. and L.J. Fleishman, 2002. Evidence for habitat partitioning based on adaptation to environmental light in a pair of sympatric lizard species. Proc. Royal Soci. Lond. B: Biol. Sci., 269: 351-359.
CrossRefDirect Link - Li, J.Y. and K.H. Lu, 1982. Population ecology of the lizard Japalura swinhonis formosensis (Sauria: Agamidae) in Taiwan. Copeia, 1982: 425-434.
CrossRefDirect Link - Losos, J.B., M. Butler and T.W. Schoener, 2003. Sexual Dimorphism in Body Size and Shape in Relation to Habitat use Among Species of Caribbean Anolis lizards. In: Lizard Social Behavior, Fox, S.F., J.K. McCoy and T.A. Baird (Eds.). John Hopkins University Press, Biltmore, London, ISBN: 9780801868931, pp: 356-380.
- Martin, J. and A. Salvador, 1997. Microhabitat selection by the Iberian rock lizard Lacerta monticola: Effects on density and spatial distribution of individuals. Biol. Conserv., 79: 303-307.
CrossRefDirect Link - Melville, J. and J.A. Schulte II, 2001. Correlates of active body temperatures and microhabitat occupation in nine species of central Australian agamid lizards. Austral Ecol., 26: 660-669.
Direct Link - Nicholson, K.L., S.M. Torrence, D.M. Ghioca, J. Bhattacharjee and A.E. Andrei et al., 2005. The influence of temperature and humidity on activity patterns of the lizards Anolis stratulus and Ameiva exsul in the British Virgin Islands. Caribbean J. Sci., 41: 870-873.
Direct Link - Radder, R.S., S.K. Saidapur and B.A. Shanbhag, 2005. Population density, microhabitat use and activity pattern of the Indian rock lizard, Psammophilus dorsalis (Agamidae). Curr. Sci., 89: 560-566.
Direct Link - Rose, B., 1981. Factors affecting activity in Sceloporus virgatus. Ecology, 62: 706-716.
CrossRefDirect Link - Schell, P.T., R. Powell, J.S. Parmerlee, A. Lathrop and D.D. Smith, 1993. Notes on the natural history of Ameiva chrysolaema (Sauria: Teiidae) from Barahona, Dominican Republic. Copeia, 1993: 859-862.
CrossRefDirect Link - Seebacher, F. and C.E. Franklin, 2005. Physiological mechanisms of thermoregulation in reptiles: A review. J. Compar. Physiol. B., 175: 533-541.
CrossRefDirect Link - Zamora-Camacho, F.J., S. Reguera, G. Moreno-Rueda and J.M. Pleguezuelos, 2013. Patterns of seasonal activity in a Mediterranean lizard along a 2200m altitudinal gradient. J. Thermal Biol., 38: 64-69.
CrossRefDirect Link - Zucker, N., 1986. Perch height preferences of male and female tree lizards, Urosaurus ornatus: A matter of food competition or social role? J. Herpetol., 20: 547-553.
CrossRefDirect Link