
U.E. Bais
Department of Zoology, Indira Gandhi (sr.) College, CIDCO, New Nanded, India
M.V. Lokhande
Department of Zoology, Indira Gandhi (sr.) College, CIDCO, New Nanded, India
International Journal of Zoological Research
Year: 2012 | Volume: 8 | Issue: 1 | Page No.: 23-32







ABSTRACT
Histopathological changes were observed in the gill, liver, stomach and intestine of Ophiocephallus striatus exposed to the cadmium chloride. The fishes were exposed to medinlethal concentration of 0.63 mg L-1 at 96 h. The histopathological change due to the toxicity of cadium chloride in experimental gills of fish observed hypertrophy, destroy of gills lamellae and blood congestion possibly caused the haematological changes in experimental fishes like reduced RBC count, decreased blood hemoglobin and anaemia. In experimental liver of fish observed degenerative changes, necrosis and hypertrophy possibly responsible for biochemical changes like reduced liver glycogen. Hepatocelluar dissociation in experimental fish possibly caused the decreased tissue glycogen and lipid in liver. Degenerative changes (ex., cloudy swelling) possibly caused the endocrine dysfunction leads to increases transaminase activities (hypersecretion of SGOT and SGPT) in present study of haematological study. In experimental stomach of fish hyperchromasia, disintegration of epithelium duct and desquamation of gastric mucosa is observed. Desquamation is a postmortem autolysis which indicates the extremely severe harmful effect of cadmium on the stomach of fish. In experimental intestine of fish degenerative changes in the tips of villi like hydropic degeneration, cloudy swelling and necrosis is observed the histological changes is possibly due to the fulfillment of extra energy requirement under the toxicity of cadmium chloride.
PDF Abstract XML References Citation
Received: August 06, 2011;
Accepted: November 04, 2011;
Published: January 13, 2012
How to cite this article
U.E. Bais and M.V. Lokhande, 2012. Effect of Cadmium Chloride on Histopathological Changes in the Freshwater Fish Ophiocephalus striatus (Channa). International Journal of Zoological Research, 8: 23-32.
DOI: 10.3923/ijzr.2012.23.32
URL: https://scialert.net/abstract/?doi=ijzr.2012.23.32
DOI: 10.3923/ijzr.2012.23.32
URL: https://scialert.net/abstract/?doi=ijzr.2012.23.32
INTRODUCTION
Histopathology is promising field for research in aquatic toxicology as it provides the real picture of the toxic of xenobiotics in vital functions of a living organism (Anees, 1976). The extent of histopathological damages induced in the Tet fish and the amount of cell damages in relation to concentration of toxicants are utilized in assessing the toxicity of pollutants. Couch (1975) stated that gill, liver, intestine and kidney of fishes are best suited organs for histopathological studies. Cadmium is naturally released to the environment from volcanic sources (some of 60% of total natural emission) and along with rain water it reaches up to the water bodies (Boyland, 1998). Small amount of cadmium minerals are also associated with lead minerals. Cadmium occurs in earth crust along with zinc, lead-zinc compounds, lead zinc-copper ores. It is usually found as cadmium sulphide. The average concentration in earth curst is about 0.2 ppm cadmium compounds such as cadmium oxides, carbonates, sulphide and hydroxide are insoluble in water but cadmium flourides, bromide, chlorides, iodide, nitrate and sulphate are particularly water soluble. Cadmium is used for electroplating of metals (Butterworth, 1995). The histopathological studies on fish revealed that various toxicants produce pathological changes such as necrobiotic changes in the liver, tubular damage of kidney, gill lamellar abnormalities and also toxic-teratological abnormalities. The objective of the present study is to highlight various histopathological changes in gills, liver, stomach and intestine of Ophiocephalus striatus exposed to cadmium chloride for 96 h.
MATERIALS AND METHODS
Healthy test fish Ophiocephalus striatus weights 100 g and total length 20 to 25 + cm were selected and exposed to sublethal concentration (LC50 0.63 mg L-1 at 96 h) of cadmium chloride. All the fishes were dissected out on the 4th day (96 h) at median lethal (LC50) concentration of cadmium chloride along with control. Gill, liver, stomach and intestine of experimental (LC50) and control at 96 h fishes were separated and immediately transferred into 10% buffered formaldehyde solution. At the end of experiment fish was taken out, sacrificed and the tissue of gill, liver, stomach and intestine excised out. The tissues were fixed in Bouin’s fluid and then they were processed by the usual procedure and embedded in paraffin wax. The tissues then processed for clearing by using xylene and transferred to molten paraffin (58-60°C). Prepared blocks for tissue holding section was done at 3 micron on rotatory microscope-after staining. Haematoxylin and eosin staining given nuclei in blue colour stain and rest tissues parts were identifying by pink colour staining.
Mounting: Each individual stained tissue slide was mounted with paraffin. Place one drop of paraffin on cover slip and glass slide were reverse in position kept for drying for one day. These mounted slides of individual slide were used for microscopic examination.
Microscopic examination: All the tissues microscopic view taken at high-resolution power with the help of Panasonic 7 megapixel digital camera. All the slides were observed under low and high resolution for their histological findings.
Histological study of gill: The respiratory organs of an animal are so constructed as to obtain necessary oxygen for the purpose of intracellular oxidation and liberation of energy for the maintenance of life and get rid of CO2. In vertebrates, the respiratory surfaces are of various kinds. Some of these are suited for respiration in water while others for a terrestrial or ariel mode of life.
The main respiratory organs in fishes are the gills. Also, the gill is the first organ after the skin which is directly exposed to the toxicant or pollutant. So that, alteration in gill structure is the early indicator of toxicant or pollution under the stress condition. So that, assessment of gill histology is a good indicator of health status of fishes.
Histological study of liver: The liver plays a vital role in the metabolic processes of the body. These processes can be broadly classified as synthesis storage and excretion. The liver synthesizes a variety of organic compounds like albumin fibrinogen urea, uric acid prothrombin lipoprotein transferring glycoprotein hippuric acid cholesterol and other lipids.
Glucose is stored in the liver as glycogen and is return to the blood circulation when the glucose level depletes other than glycogen the liver stores fat soluble vitamins (A, D, E and K) and vitamin B12. The excretory and detoxifying function of the liver are evident from the process of removal of free bilirubin, the product of haemoglobin breakdown. Many toxic substances are similarly removed from blood circulation through the bile duct, many of these substances are insoluble in water and hence cannot follow the kidney route. Diagnostic test results for the assessment of liver function include increases serum transaminase activity.
Histological study of stomach: Stomach basically is a structure meant for the storage of food stomach assumes different shapes according to the availability of space in the body cavities of different fishes. It is usually differentiated in to a broad anterior part lying closer to the heart and called cardiac stomach and into a posterior narrower part called pyloric stomach. Stomach of teleosts presents a variety of different shapes. An acidic gastric juice is secreted by the mucosa of the stomach in carnivores fishes. So that, stomach is important organ in the digestive system of teleosts fishes.
So that, histopathological study of helpful in assessment of health status as well as metabolism of fishes.
Histological study of intestine: Intestine is the important organ of digestive system of fishes. It is divisible in to two main parts an anterior long but narrower part called small intestine and a posterior short but broader part called the large intestine.
The functional importance of intestine is absorption. Maximum absorption of digested food products takes place in intestine.
Along with abortion of food there are several other function of intestine is protective function, activator function, hemopoietic function, hormonal function etc. So that, it is important to access the histological structure of intestine as a diagnostic tool for normal health status of the fish.
RESULTS
The normal structure is observed in the gill of control fish (Fig. 1) where as, in experimental (LC50) hypertrophy of lamellar epithelium, destroy of gill lamellae and blood congestion is observed (Fig. 2).
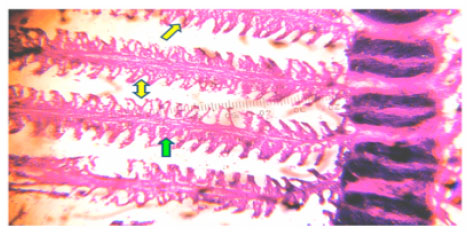 | |
| Fig. 1: | Photomicrograph of Ophiocephalus striatus gill control at 96 h shows normal gill lamellae (yellow arrow) normal epithelial cells (yellow arrow) |
 | |
| Fig. 2: | Photomicrograph of Ophiocephalus striatus gill experimental (median lethal LC50) at 96 h shows hypertomy (green arrow) blood conjugation (yellow arrow) destroy of gill lamellae (red arrow) under light microscope |
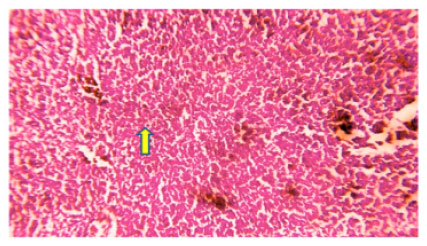 | |
| Fig. 3: | Photomicrograph of Ophiocephalus striatus liver control at 96 h shows normal hepatic tissues showing hepatocytes with granular cytoplasm with round nucleus hepatocytes cells (yellow arrow) |
In control fish normal hepatic tissue showing hepatocytes with granular cytoplasm with round nucleus hepatocytes cells is observed in liver (Fig. 3) whereas, in experimental (LC50) at 96 h degenerative changes hepatocellular dissociation necrosis and hypertrophy is observed in experimental (LC50) at 96 h liver of fish (Fig. 4).
In control fish stomach shows normal epithelial duct, normal submucosa with blood capillary (Fig. 5). Whereas, in experimental (LC50) at 96 h stomach of fish shows hyperchromasia of epithelium cells, disintegration of epithelium duct and desquamation of gastric mucosa is observed (Fig. 6).
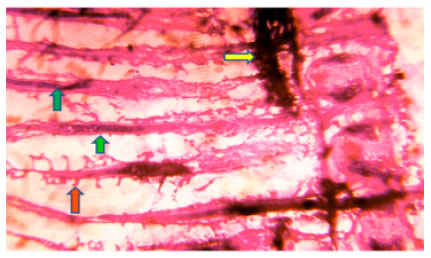
In control fish normal columnar epithelium, circular muscle fibre and normal intestinal submucosa is observed (Fig. 7).
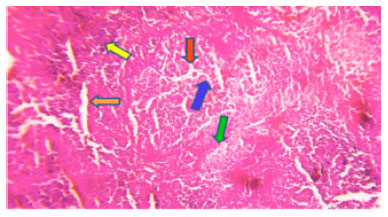 | |
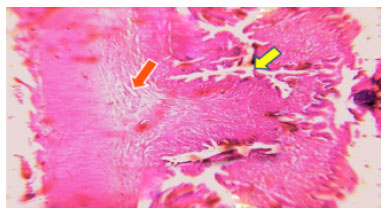
| Fig. 4: | Photomicrograph of Ophiocephalus striatus liver experimental (median lethal LC50) at 96 h shows hepatocellular dissociation (green arrow) degenerative changes includes cloudy swelling (yellow arrow) hydropic degeneration (red arrow) hypertrophy (blue arrow) necrosis (orange arrow) |
 | |
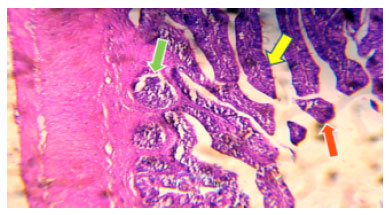
| Fig. 5: | Photomicrograph of Ophiocephalus striatus stomach control shows epithelial duct (yellow arrow) normal submucosa with blood capillaries (red arrow) |
 | |
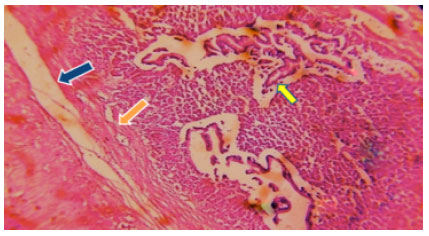
| Fig. 6: | Photomicrograph of Ophicephalus striatus stomach experimental shows hyperchromatic of epithelial duct (red arrow) desquamation gastric mucosa (green arrow) |
 | |
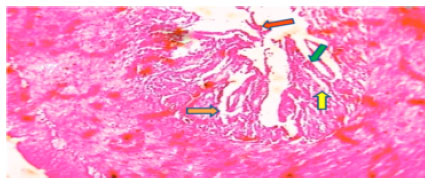
| Fig. 7: | Photomicrograph of Ophiocephalus striatus intestine control at 96 h shows normal columnar epithelium (yellow arrow) circular muscle fibre (blue arrow) and normal intestinal submucosa with blood capillaries (orange arrow) |
 | |
| Fig. 8: | Photomicrograph of Ophiocephalus striatus intestine experimental LC50 at 96 h shows degenerative changes in tips of villi as hydropic degeneration (yellow arrow) loss of structural integrity of mucosal folds (red arrow) degenerative change of mucosal epithelium as cloudy swelling (green arrow) and necrosis (orange arrow) |
Whereas, in experimental (LC50) at 96 h degenerative changes in the tips of villi as hydropic degeneration, loss of structural integrity of mucosal folds, degenerative changes of mucosal epithelium as cloudy swelling and necrosis is observed (Fig. 8) in the intestine of fish.
DISCUSSION
The gills of the brown bullhead, Ictalurus nebulosus exposed to experimental copper solution of 5 ppm showed swollen and hyperemic lamellae, necrosis and disaggregation of the epithelium (Benedetti et al., 1989). Gill damage at 0.3 ppm copper varied depending on individual stress, where gills of stressed specimens were very swollen and showed degeneration of the gill epithelium with focal necrosis.
Two species of telecosts the zebrafish Brachydanio rerio and the Rainbow trout, Salmo gairdneri were exposed to different concentration of cadmium and the gill morphology was assessed following 6 weeks of exposure (Karlsson-Norrgren et al., 1985). At low cadmium concentration of 3 μg lit., the minor changes in gill alterations in the chloride cells of the secondary lamellae were observed compare with that of the controls in zebra fish.
In gills of treated fish, the number of mitosis was significantly higher in both primary and secondary epithelium than in control gills. Newly formed epithelial cells were observed to differentiate into chloride and mucus cells. Gardner and Yevich (1970) shows microscopic examination revealed pathological changes in the gill filaments and respiratory lamellae of the Mummichog fundulus heteroclitus after 20 h exposure to 50 ppm of cadmium. Histopathological evaluation showed necrosis and sloughing of respiratory filaments, hypertrophy of gills filaments and hyperplasia of the epithelial surface of respiratory lamellae and interlamellar filament epithelium. The hyperplasia was accompanied by a marked increase in mitotic figures. Changes did not involve all gill filament and were not localized to specific areas of the bronchial arches.
Athikesavan et al. (2006) stated that nickel showed a tissue specific alteration in the tissues, mucus proliferation, fusion of the gill lamellae and hypertrophy of gill tissue were observed in fresh water fish Hypothalamichthys molitrix sub-lethal exposed to nickel.
In teleostean fishes each gills possess primary and secondary gill lamellae in indicated with red arrow in experimental fishes photograph. Primary gill lamellae possess a epithelial cells layer called primary epithelium and their principle function is to convey the dissolved oxygen with the help of RBC. In present study, gill lamellae is greatly destroyed in experimental fishes and possibly caused the severe respiratory problem and reduced RBC in blood. Also hypertrophy in primary gill lamellae of experimental fishes indicates increased functional demand of RBC under toxicant condition.
Hypertrophy cans occur from an increase in functional demand which may be compensatory or hormonal in organ. In present investigation hypertrophy of lamellar epithelium observed in experimental (LC50) fish possibly caused to stress condition of cadmium and increases the respiratory rate.
In a histopathological study of the effects of experimental copper pollution at high concentration (5 ppm) and low concentration (0.3 ppm) on bullheads inctaturus nebulosus analysis of liver exposed at either concentration did not reveal diffuse changes in the hepatic parenchyma (Benedetti et al., 1989) However, the authors reported areas of patchy hepatic degeneration. Histochemical staining further demonstrated lower liver glycogen contents than in the controls in all treatments with scattered areas of normal and depleted glycogen content in the treated specimen.
In present study, depleted liver glycogen in biochemical studies is possibly due to the degenerative changes in experimental (LC50) at 96 h liver of fish. A histopathology study in snake head fish Channa punctatus to the effects of 6.8 mg L-1 lead nitrate for 125 days showed liver cord disarray necrosis inflammation of portal areas hardening of connective tissue shrinkage of nucleus and septa formation around blood vessels (Sastry and Gupta, 1978b) No fatty infiltration or glycogen depletion was observed Treatment of snake head fish Channa punctuates with a sub-lethal concentration of lead nitrate produced considerable hepatic structural damage and decreased alkaline phosphate and aminopeptidase’s activities.
A series of experiments by Sastry and Gupta (1978a) have shown that significant pathological changes were produced by 0.3 mg L-1 mercuric chloride in the liver of snake head fish Channa punctatus over a period of 30 days. Histopathological changes included hepatocellular granulation and vacuolation of the cytoplasm hypertrophy of nucleus necrosis, fatty infiltration and proliferation of connective tissue, glycogen depletion and cirrhosis.
Above similar kinds of changes is observed in the experimental (LC50) at 96 h liver of fish in present study of experimental liver of fish observed hepatocellular dissociation which leads to the complete dissolution of hepatocytes. Hepatocytes can contains glycogen and lipids which are related to the normal metabolic function of the liver. In present study of biochemical changes both glycogen and lipid are significantly depleted in experimental (LC50) at 96 h liver of fish so it is evident that decreased liver glycogen and lipid is caused due to the histological changes in experimental liver of fish. Cloudy swelling is characterized by the enlarged cell size possibly due to the endocrine dysfunction leads to the increase in transaminase activities. In present study of haematological studies SGPT and SGOT level is increased significantly in experimental fish over the control. So, it is also evident that liver dysfunction also responsible for increased SGPT and SGOT level in experimental haematological studies.
In present study, experimental (LC50) at 96 h stomach of fish shows hyperchromatic epithelial cells possibly due to the cadmium toxicity. Also experimental (LC50) at 96 h stomach observed disintegration of glandular epithelium duct and desquamation of gastric mucosa. Desquamation is characterized by sloughing of cell from an epithelial surface due to necrosis or some degenerative changes or from postmortem autolysis.
Bhatnager and Shrivastava (1975) reported tunica muscularis with nuclear depolarization and cytoplasmic hyperchromasia, general edema, loss of pepsinogen granules, disintegration of epithelium duct with desquamation of gastric mucosa in stomach of Heteropheustus fossilis exposed to copper. Sastry and Gupta (1978a) mentioned the cytoplasmic hyperchromasia, edema, loss of pepsinogen granules from chief cells, disintegration of glandular epithelium, desquamation of gastric mucosa. In the stomach of Channa punctatus exposed to sub-lethal concentration of mercuric chloride.
Sastry and Gupta (1978b) reported that hypersecretion of pepsin is leads to the degradation of tissue protein in Channa punctutus, also mentioned increased ammonia and urea in the experimental fish exposed to heavy metal lead nitrate. He concluded that these condition is possibly due to the extremely adverse effects on stomach of fish due to lead nitrate toxicity.
In present study, in experimental (LC50) at 96 h tissue protein is decreased in biochemical studies. So, it is evident that alteration in histology of experimental organ (e.g., stomach) their reflection is seen in biochemical as well as haematological studies in present study. So that, from above discussion very adverse effects of cadmium on the experimental stomach of fish is concluded.
Gardner and Yevich (1970), Gutierrez et al. (1978) and Newman and MacLean (1974) reported that toxic lesions most common in the intestine, include hyperemia, degenerative changes in the tips of villi, loss of structural integrity of mucosal folds, degenerative mucosal epithelium (hypertrophy, vacuolation, hyper-chromasia) necrosis, desquamation of mucosal epithelium, cellular debris, excessive mucus in gut of lumen, necrosis of submucosa and inflammatory infiltration of submucosa in intestine of different fishes exposed to Cadmium Chloride (CdCl2).
Gardner and Yevich (1970) mentioned the histopathological changes in intestine like hypertrophy leads to the increased serum glucose in estuarine teleost fish exposed to cadmium. He also concluded that this condition is possibly due to the fulfillment of extra energy requirement under stress condition.
Teleostean fishes intestine is lined by a simple columnar epithelium consisting of food absorptive cells and mucus secreting cells. In present study of experimental intestine disintegration of mucosal fold and mucosal epithelium possibly caused the severe problem in absorption of food materials. Also necrosis condition indicates that complete dissolution of intestinal cells caused the severe problem in digestion of food.
Establier et al. (1978) reported intestinal toxic lesions, includes hyperemia, loss of structural integrity of mucosal folds, necrosis, cellular debris, vacuolation in intestine of Mugil auratus exposed to inorganic and organic mercury. Sastry and Gupta (1978a) reported the toxic lesions includes hyperemia, degenerative changes in the hips of mucosal folds, hypertrophy and necrosis in intestine of Channa punctatus exposed to mercuric chloride. Anees (1976) reported that toxic lesions in the intestine include hyperemia, degenerative changes in the tips of villi loss of structural integrity of mucosal folds, degeneration of mucosal epithelium, include hypertrophy necrosis, increased number of mucus goblet cells, vacuolation etc. in fresh water telecast Channa punctatus exposed to sub lethal and chronic levels of there organophosphotous insecticides. Walsh and Ribelin (1975) reported hyperemia, degenerative changes in the tips of villi, loss of structural integrity of mucosal folds, hypertrophy vacuolation and necrosis in the intestine of Cyprinus carpio exposed to the pesticide atrazine. Smith and Piper (1975) reported the toxic lesions in the intestine include hyperemia, degenerative changes in the tips of villi, hypertrophy, vacuolation hyperchromasia, necrosis, desquamation of mucosal epithelium, submucosal vasculature is observed in different fishes chronically exposed to ammonia.
From above discussion and present study of histology of intestine it is clear that very adverse effect of cadmium chloride on the intestine of fish. Also intestinal changes can be used for indicator of biochemical and haematological changes in experimental (LC50) at 96 h fishes in present study.
CONCLUSION
In experimental gills of fish observed hypertrophy, destroy of gills lamellae and blood congestion possibly caused the haematological changes in experimental fishes like reduced RBC count, decreased blood hemoglobin and anemia. In experimental (LC50) at 96 h liver of fish observed degenerative changes, necrosis and hypertrophy possibly responsible for biochemical changes like reduced liver glycogen. Hepatocellular dissociation in experimental fish possibly caused the decreased tissue glycogen and lipid in liver. In experimental (LC50) at 96 h pancreas of fish congestion, hyperplasia, edema and necrosis is observed. Hyperplasia possibly responsible for the increased serum glucose level. Hyperplasia conditions indicate endocrine dysfunction (hyperglycemic condition).
In experimental (LC50) at 96 h stomach of fish hyperchromasia, disintegration of epithelium duct and desquamation of gastric mucosa is observed. Desquamation is a postmortem autolysis which indicates the extremely severe harmful effect of cadmium on the stomach of fish.
In experimental (LC50) at 96 h intestine of fish degenerative changes in the tips of villi like hydropic degeneration, cloudy swelling and necrosis is observed the histological changes is possibly due to the fulfillment of extra energy requirement under the toxicity of cadmium chloride. After above overall consideration and result it has been concluded that heavy metal cadmium causes deleterious effects on Ophiocephalus (channa) striatus and significantly alters the normal metabolism and reproduction and ultimately growth of the fishes. Affected growth of fish possibly affects the whole community and trophic levels of food chain and ultimately on the aquatic ecosystem.
REFERENCES
- Anees, M.A., 1976. Intestinal pathology in a freshwater teleost, Channa punctatus (bloch) exposed to sub-lethal and chronic levels of three organophosphorus insecticides. Acta Physiol. Lat. Am., 26: 63-67.
PubMed - Athikesavan, S., S. Vincent, T. Ambrose and B. Velmurugan, 2006. Nickel induced histopathological changes in the different tissues of freshwater fish, Hypophthalmichthys molitrix (Valenciennes). J. Environ. Biol., 27: 391-395.
PubMedDirect Link - Benedetti, I., A.G. Albano and L. Mola, 1989. Histomorphological changes in some organs of the brown bullhead, Ictalurus nebulosus LeSueur, following short- and long-term exposure to copper. J. Fish Biol., 34: 273-280.
CrossRef - Gardner, G.R. and P.P. Yevich, 1970. Histological and hematological responses of an estuarine teleost to cadmium. J. Fish Res. Board, Canada, 27: 2185-2196.
CrossRefDirect Link - Karlsson-Norrgren, L., P. Runn, C. Haux and L. Forlin, 1985. Cadmium-induced changes in gill morphology of zebrafish, Brachydanio rerio (Hamilton-Buchanan) and rainbow trout, Salmo gairdneri Richardson. J. Fish Biol., 27: 81-95.
CrossRef - Sastry, K.V. and P.K. Gupta, 1978. Effects of merucuric chloride on the digestive system of Channa punctutus: A hstopathological study. Environ. Res, 16: 270-278.
CrossRefDirect Link - Sastry, K.V. and P.K. Gupta, 1978. Histopathological and enzymological studies on the effects of chronic lead nitrate intoxication in the digestive system of a freshwater teleost, Channa punctatus. Environ. Res., 17: 472-479.
CrossRefPubMedDirect Link